Trends Sci. 2026; 23(10): 13197
miR-15b-5p in Cancers: Expression Patterns and Regulatory Pathways
Kharisma Arethusa Maisaroh1, Putri Rohmatul Laili1, Reza Adhi Pratama1,
Auraga Dewantoro1, Dyah Laksmi Dewi2, Teguh Aryandono2 and Irianiwati Widodo3,*
1Master Program in Biotechnology, Graduate School of Universitas Gadjah Mada, Yogyakarta, Indonesia
2Division of Surgical Oncology, Department of Surgery, Faculty of Medicine, Public Health and Nursing,
Universitas Gadjah Mada, Yogyakarta, Indonesia
3Department of Anatomical Pathology, Department of Surgery, Faculty of Medicine, Public Health and Nursing, Universitas Gadjah Mada, Yogyakarta, Indonesia
(*Corresponding author’s e-mail: [email protected])
Received: 29 December 2025, Revised: 20 March 2026, Accepted: 27 March 2026, Published: 20 May 2026
Abstract
miR-15b-5p is a multifunctional microRNA (miRNA) that is highly context-dependent in cancer, acting as either an oncomiR or a tumor suppressor miRNA (TSM) depending on tumor type, molecular characteristics, and microenvironment. This review synthesizes existing evidence to outline pathway and network integration underlying the context-dependent biological roles of miR-15b-5p across different cancers. Integrated multi-omics cancer profiling demonstrates significant upregulation across several solid tumor types, including non-small cell lung cancer (NSCLC), colorectal cancer (CRC), prostate cancer (PCa), melanoma, and early-stage breast cancer. Conversely, downregulation has been observed in some malignancies, such as Hodgkin’s lymphoma, renal cell carcinoma, and KRAS-mutant CRC. This indicates the dual roles of miR-15b-5p. Upstream, miR-15b-5p is regulated by multiple lncRNA-miRNA-mRNA axes, which affect key targets such as CHRM3, PPM1D, CCND1, CDC42, CA2, and CCNE2, which are involved in oncogenic pathways such as PI3K/Akt, NF-κB, p53, MAPK/ERK, Wnt/β-catenin, TGF-β, and VEGF signaling. In our analysis, the PI3K/Akt network and its interaction with p53 and mTOR appear to be prominently affected signaling pathways, although other regulators such as PAQR3, AXIN2, and ACVR2A may contribute to broader tumor-specific phenotypes. By systematically integrating evidence across diverse malignancies and distinguishing experimentally validated targets from predicted interactions, this study provides a consolidated and mechanistically grounded framework positioning miR-15b-5p as a context-dependent therapeutic, prognostic, and diagnostic candidate. While current evidence remains largely preclinical, our synthesis clarifies inconsistencies in the literature and establishes a structured foundation for future clinical validation.
Keywords: miRNA, miR-15b-5p, Profiling, OncomiR, Tumor suppressor miRNA, Pathways, Cancers
Introduction
MicroRNAs (miRNAs) are 19-25-nucleotide non-coding RNAs that function as regulators of gene expression, primarily at the post-transcriptional level [1,2]. miRNA biogenesis begins with the transcription of pri-miRNAs by RNA polymerases II/III, then processed by the Drosha-DGCR8 complex into hairpin-structured pre-miRNAs. These pre-miRNAs are then exported to the cytoplasm via Exportin-5 and subsequently cleaved by Dicer to generate mature miRNAs, then loaded into Argonaute (AGO) proteins to mediate gene regulation [2–4]. The first miRNA, lin-4 and lin-7, initially discovered in Caenorhabditis elegans, were thought to be small RNAs without biological function, but their conservation across species later revealed essential regulatory roles. It is known that more than 1,000 human miRNAs regulate over 30% of mammalian genes, underscoring their importance as key components of gene-expression control [1–3].
miRNAs regulate gene expression by binding to the 3’UTR of target mRNAs, where the degree of complementarity determines whether regulation proceeds through mRNA degradation or translational repression. Because a single miRNA can regulate multiple transcripts, and each mRNA can be targeted by several miRNAs, target identification requires integrating bioinformatic predictions with experimental validation [2,5,6]. In cancer, miRNAs modulate genes involved in essential cellular processes such as proliferation, apoptosis, differentiation, metabolism, metastasis, and immune response [7–10].
Dysregulation of miRNA expression disrupts molecular homeostasis and drives tumor initiation, progression, and heterogeneity, ultimately shaping their potential as diagnostic, prognostic, and therapeutic-stratification biomarkers [1,4,11]. Notably, the miR-15 family targets several oncogenes, including BCL2, Cyclin D1, and MCL1, as well as key cell cycle-related genes such as cyclin B, cyclin D1, cyclin D3, and cyclin E, thereby suppressing cancer cell proliferation. The miR-15 family can partially compensate for p53 loss and is regulated by epigenetic mechanisms. Therefore, epigenetic modulators that restore miR-15 expression represent a promising strategy for miRNA-based anticancer therapy. Thus, the miR-15 family emerges as a tumor suppressor in cancer [12].
miR-15b-5p belongs to the miR-15 family encoded by the MIR15B gene, which is located on chromosome 3q25.33 [13,14]. miR-15b-5p is upregulated in NSCLC [15], gastric cancer [16], and PCa [17] compared to healthy controls. Meanwhile, it has also been found to be downregulated in KRAS-mutated CRC compared to non-mutated samples [18] and classical Hodgkin lymphoma (cHL) compared to reactive lymphadenopathy samples [19]. Previous studies suggest that miR-15b-5p has a context-dependent dual function, acting either as an oncomiR or TSM, reflecting its complexity and tissue-specific regulatory roles in cancer [14].
Unlike previous reviews that primarily summarized individual targets, this review proposes a pathway-integrated conceptual framework by explaining how miR-15b-5p exerts context-dependent dual roles across cancers. Rather than acting through a single linear lncRNA-miRNA-mRNA axis, miR-15b-5p functions as a regulatory node within interconnected signaling networks, coordinately modulating multiple downstream pathways and contributing to its context-dependent effects across various cancers. This network-oriented perspective may help reconcile conflicting reports, which describe it as either oncogenic or tumor-suppressive and provide a conceptual basis for future mechanistic and translational studies.
Materials and methods
This study utilized narrative approaches to conduct a comprehensive review analysis. Relevant literature was searched using the PubMed database with the keywords “miR-15b-5p; AND cancer” within the search section. In addition to PubMed, we searched on Google Scholar using the same keywords to support relevant literature.
The initial screening was performed based on titles and abstracts. The inclusion criteria for the literature comprised original research articles and review papers published in English between 2015 and 2025. Articles identified during the initial screening were further assessed through full-text review to determine their relevance to the scope of this review. Non-English publications, lacking cancer-related studies, and non-human biological samples, such as animal cancer cell line models, were excluded from the analysis. To improve interpretability, this review differentiates between predicted and experimentally validated miR-15b-5p target genes. Predicted targets were derived from bioinformatic and expression profiling studies, whereas validated targets were supported by in vitro and in vivo studies.
To ensure systematic evaluation, selected studies were organized using a structured Excel-based checklist, which included information on miRNA type, associated lncRNAs, target genes, major findings, experimental methods, sample type (e.g., plasma, tissue, FFPE), study population, and DOI. This structured evaluation facilitated the identification of studies that were sufficiently relevant and informative to serve as the primary sources for this review.
The extracted data then categorized into 4 main areas:
The dual roles of miR-15b-5p in cancers, including its oncogenic and tumor-suppressive functions through profiling studies.
Upstream regulatory mechanisms influencing miR-15b-5p.
Experimentally validated and predicted target genes of miR-15b-5p and their involvement in key signaling pathways.
The potential clinical implications of miR-15b-5p, including its therapeutic relevance in reducing cancer aggressiveness and improving treatment outcomes.
To guide the analysis, several research questions were addressed, including how miR-15b-5p exhibits dual roles in cancer through profiling studies and how its upstream regulators influence its expression in different tumor contexts. We also examined how miR-15b-5p regulates multiple signaling pathways through its predicted and experimentally validated target genes. Finally, we discussed the broader implications of miR-15b-5p in cancer biology, including its potential therapeutic relevance in attenuating tumor aggressiveness and improving treatment outcomes.
Results and discussions
Profiling miR-15b-5p and its dual roles in cancer
Profiling studies consistently show that miR-15b-5p is dysregulated in various cancer types. This dysregulation has been detected using quantitative Real-Time Reverse Transcription PCR (qRT-PCR) [15,20–22], droplet digital Reverse Transcription PCR (dd-RT-PCR) [23], next-generation sequencing (NGS) [24,25], microarray [16,26], NanoString [27,28], and miRNA-seq [15,29].
miR-15b-5p is commonly upregulated, particularly in solid tumors. In NSCLC, miR-15b-5p is significantly upregulated in serum, plasma, and tumor-derived exosomes, whereas in CRC, it is upregulated in both systemic and tumor tissues. PCa similarly shows robust upregulation in serum and at the transcriptomic level, both in single-cell and bulk RNA-seq datasets [17,30]. This consistent upregulation suggests that miR-15b-5p may function as a conserved oncogenic factor in various malignancies.
Moreover, miR-15b-5p has also been reported to be downregulated in several types of cancer, such as cHL [19], renal cell carcinoma [31], Erdheim-Chester disease (ECD) [27], and specific gastrointestinal cancer [29]. In CRC, however, its expression appears to be compartment-specific (Table 1). miR-15b-5p is upregulated in CRC serum compared with healthy controls [32], whereas miR-15b-5p is significantly downregulated in KRAS-mutated tissues compared to wild-type counterparts [18]. Interestingly, plasma samples from KRAS-mutated CRC have shown increased miR-15b-5p levels [23]. This apparent discrepancy may reflect differences between intracellular and circulating miRNA dynamics. While miR-15b-5p appears to be downregulated in KRAS-mutated tumor tissues, tumor cells may actively release miRNAs into the circulation through extracellular vesicles (EV) [33] or RNA-binding protein complexes [34]. In addition, evidence suggests that miRNA loading into EV is not a passive process but may involve selective sorting mechanisms. Consistent with this, several diseases, including cancer, exhibit dysregulated EV-miRNA profiles, suggesting selective miRNA packaging into extracellular vesicles during pathogenesis [34]. This may contribute to the elevated circulating levels observed in plasma or serum. However, this relationship needs further investigation, particularly regarding the correlation between miR-15b-5p expression in tumor tissues and plasma. Studies analyzing paired tissue and plasma samples collected at the same time point would be important to better elucidate this relationship in CRC. Furthermore, given that KRAS mutations drive constitutive activation of the EGFR downstream signaling cascade, miR-15b-5p may participate in KRAS-dependent transcriptional and post-transcriptional networks, potentially functioning within a dynamic feedback regulatory loop.
However, variations in sample types (e.g., tissue vs plasma or serum), patient characteristics, and experimental methodologies across studies may also contribute to these discrepancies. Furthermore, many mechanistic insights into miRNA secretion are derived from preclinical models, and their relevance in clinical settings remains to be fully elucidated. Collectively, these contrasting patterns suggest that miR-15b-5p regulation is highly context-dependent and influenced by tumor origin, microenvironment, and molecular characteristics.
Table 1 Summary of studies investigating the profile of miR-15b-5p in human cancer. The table shows that miR-15b-5p exhibits differential expression, either upregulated or downregulated, depending on the type of cancer.
miR-15b-5p is also frequently upregulated in early-stage tumors. This indicates that miR-15b-5p is likely involved in the initiation and early development of cancer. Elevated levels have been reported in the early stages of NSCLC [15,20,36], melanoma [37], CRC and advanced adenomas [32], as well as early breast lesions such as ductal carcinoma in situ (DCIS) and stage I breast cancer [24]. Therefore, miR-15b-5p can be developed as a promising non-invasive biomarker for early cancer detection.
miR-15b-5p exhibits a broad and reproducible deregulation profile across various malignancies. Its frequent upregulation, detectability in early disease stages, and robust cross-platform consistency underscore its potential as a clinically relevant biomarker for cancer detection and monitoring [14]. Conversely, miR-15b-5p downregulation in specific tumor types reflects the complexity of its biological mechanisms, suggesting that it may act as an oncomiR or a TSM depending on the molecular context. These dual roles highlight the importance of evaluating miR-15b-5p within the unique biological framework of each tumor type when interpreting its mechanistic role and potential clinical applications.
Upstream regulation of miR-15b-5p
Genetic variation of miRNA
Several studies have demonstrated that mutations in miRNA sequences can disrupt their biogenesis. Research by Duan et al. [46] showed that a Single Nucleotide Polymorphism (SNP) of miR-125a sequence in Human HEK293 cell lines decreases the processing of pri-miRNA to pre-miRNA, thereby reducing the miRNA’s ability to regulate its target genes. Similar findings were reported for the SNP of miR-126 in chronic lymphocytic leukemia (CLL) patient samples, diminishing the activity of the Drosha-DGCR8 complex in cleaving its pri-miRNA [47]. Sun et al. [48] also provided evidence of polymorphism-induced effects on miR-15b-5p. Although conducted in porcine, the study demonstrated that mutations within miR-15b host gene can disrupt the transition from pri- to pre-miRNA and alter strand selection, shifting the dominant mature miRNA from miR-15b-5p to miR-15b-3p loaded onto AGO. Collectively, these findings underscore that polymorphisms of miRNA genes may substantially disrupt their biogenesis and alter mature miRNA expression. Additionally, it suggests a potential impact on the regulation of downstream targets that still requires further study.
Long non-coding RNA
Long non-coding RNAs (lncRNAs), which are transcripts longer than 200 nucleotides, represent key regulatory molecules that modulate miRNA activity through multiple interconnected molecular mechanisms [49]. The well-established function of lncRNA is to act as competing endogenous RNAs (ceRNAs) by sponging miRNA through complementary base pairing of miRNA recognition elements (MREs), thereby preventing miRNA interaction with its target mRNAs [50,51]. Moreover, lncRNA also influences transcriptional and post-transcriptional miRNA biogenesis, including epigenetic regulation, miRNA maturation, and miRNA stability [49,52]. In the case of miR-15b-5p, the biological consequences are highly contextual, depending on its function as a tumor suppressor miR (TSM) or as an oncomiR. Consequently, the biological outcomes of lncRNA can function either as oncogenic or tumor-suppressive lncRNA, depending on the genes targeted by miRNA, as shown in Table 2.
Table 2 lncRNA/miR-15b-5p regulatory interactions and associated biological activities. This highlights the involvement of lncRNA/miR-15b-5p interactions in regulating cancer progression, including proliferation, migration, invasion, and apoptosis.
|
Cancer type |
lncRNA |
Target gene(s) |
Biological activity |
Citation |
Oncogenic Activity |
PCa |
TTN-AS1, TRPM2-AS, and AFAP-AS1 |
CHRM3 |
↑ cell proliferation and aggressiveness |
[53] |
Hypopharyngeal Squamous Cell Carcinoma (HPSCC) |
Linc00662 |
Not mentioned |
↑ cell
proliferation, invasion, and migration |
[35] |
|
Osteosarcoma |
TRPM2-AS |
PPM1D |
↑ cell
viability, proliferation, and migration |
[54] |
|
CRC |
CERS6‐AS1 |
SPTBN2 |
↑ cell proliferation, invasion, migration, EMT process, and stemness |
[55] |
|
Hepatocellular Carcinoma (HCC) |
H19 |
CDC42 |
↑ proliferation,
invasion, and migration |
[56] |
|
Breast Cancer (BC) |
MEG3 and FAM66E |
PIK3R1 and JUN |
↑ proliferation ↓ apoptosis |
[42] |
|
Osteosarcoma |
PVT1 |
CCND1 |
↑ cell proliferation and chemotherapy resistance |
[57] |
|
Tumor Suppressive Activity |
Bladder Cancer |
MAGI2-AS3 |
CCDC-19 |
↓cell proliferation, colony formation, migration, and invasion |
[51,58] |
HBV-induced Hepatocellular Carcinoma |
SSTR5-AS1 and FAM138B |
CA2 and CCNE2 |
↑ cell proliferation |
[50] |
|
Cervical Cancer (CC) |
FENDRR |
TUBA1A |
↓ cell
viability, proliferation, invasion, and migration |
[59] |
Recent studies consistently demonstrate that lncRNA/miR-15b-5p interactions regulate the expression of target genes involved in major oncogenic pathways. In prostate cancer, the lncRNA AFAP-AS1 decreases miR-15b-5p activity through sequence-specific interactions, resulting in overexpression of CHRM3 and activating the Hippo-YAP pathway, thereby promoting tumor proliferation, cell viability, and aggressiveness [53]. A similar upstream interaction is observed in HPSCC, where Linc00662 suppresses miR-15b-5p activity and supports cancer stem cell-like phenotypes [35]. In triple-negative breast cancer (TNBC), TCGA-based bioinformatic analyses suggest that MEG3 and FAM66E are significant upstream regulators of miR-15b-5p. Both lncRNAs show strong statistical associations with altered expression of PI3KR1 and JUN, key components of PI3K/Akt and AP-1 signalling, respectively. Contributing to the heterogeneity of proliferative and stress-response pathways characteristic of TNBC [42].
A more complex pattern is observed in osteosarcoma, where 2 different lncRNAs, TRPM2-AS and PVT1, simultaneously suppress the tumor-suppressive activity of miR-15b-5p [54,57]. TRPM2-AS upregulates PPM1D, a negative regulator of p53-mediated apoptosis, thus promoting proliferation and survival [54]. Whereas PVT1 derepresses CCND1, enhancing the G1/S transition and accelerating cell proliferation [57]. Furthermore, in HCC, miR-15b-5p demonstrates opposing biological roles depending on its specific target genes, which influence the functional activity of the lncRNA involved. H19 acts as an oncogenic lncRNA that suppresses miR-15b-5p and activates the CDC42/PAK1 axis to promote tumor proliferation, invasion, and migration [56]. Whereas SSTR5-AS1 and FAM138B exert the opposite effect by neutralizing the oncomiR activity of miR-15b-5p, maintaining CA2 and CCNE2 expression and ultimately promoting tumor suppressive signaling [50].
Overall, although lncRNAs exhibit diverse regulatory activities, available experimental evidence consistently indicates that most lncRNA/miR-15b-5p interactions occur via sponging or ceRNA mechanisms, thereby preserving the expression of target mRNAs. This predominance pattern suggests that downstream regulatory changes are driven not only by alterations in miR-15b-5p expression levels, but also by lncRNA-mediated inhibition of its biological activity. Moreover, cross-malignancy findings show that the final biological outcome is determined not solely by the presence of sponging interactions, but also by the functional context of miR-15b-5p and its target genes in each cancer type.
Downstream regulation of miR-15b-5p
miR-15b-5p targets genes across multiple signaling pathways, leading to context-dependent biological effects across various cancer types. Its predicted (bioinformatic and expression profiling) and validated (in vitro and in vivo) targets are distributed across major oncogenic networks, as illustrated in Figure 1 and summarized in Table 3. Because each target gene contributes to a distinct signaling cascade, miR-15b-5p repression results in pathway-specific effects that influence the proliferation, survival, inflammation, or migration of cancer cells. Consequently, the functional role of miR-15b-5p, whether TSM or oncomiR, is determined by which signaling pathway is predominantly affected in a given cancer type. The following section provides an integrated overview of these signaling mechanisms by illustrating the relationships between miRNA target genes and their positions within each pathway.
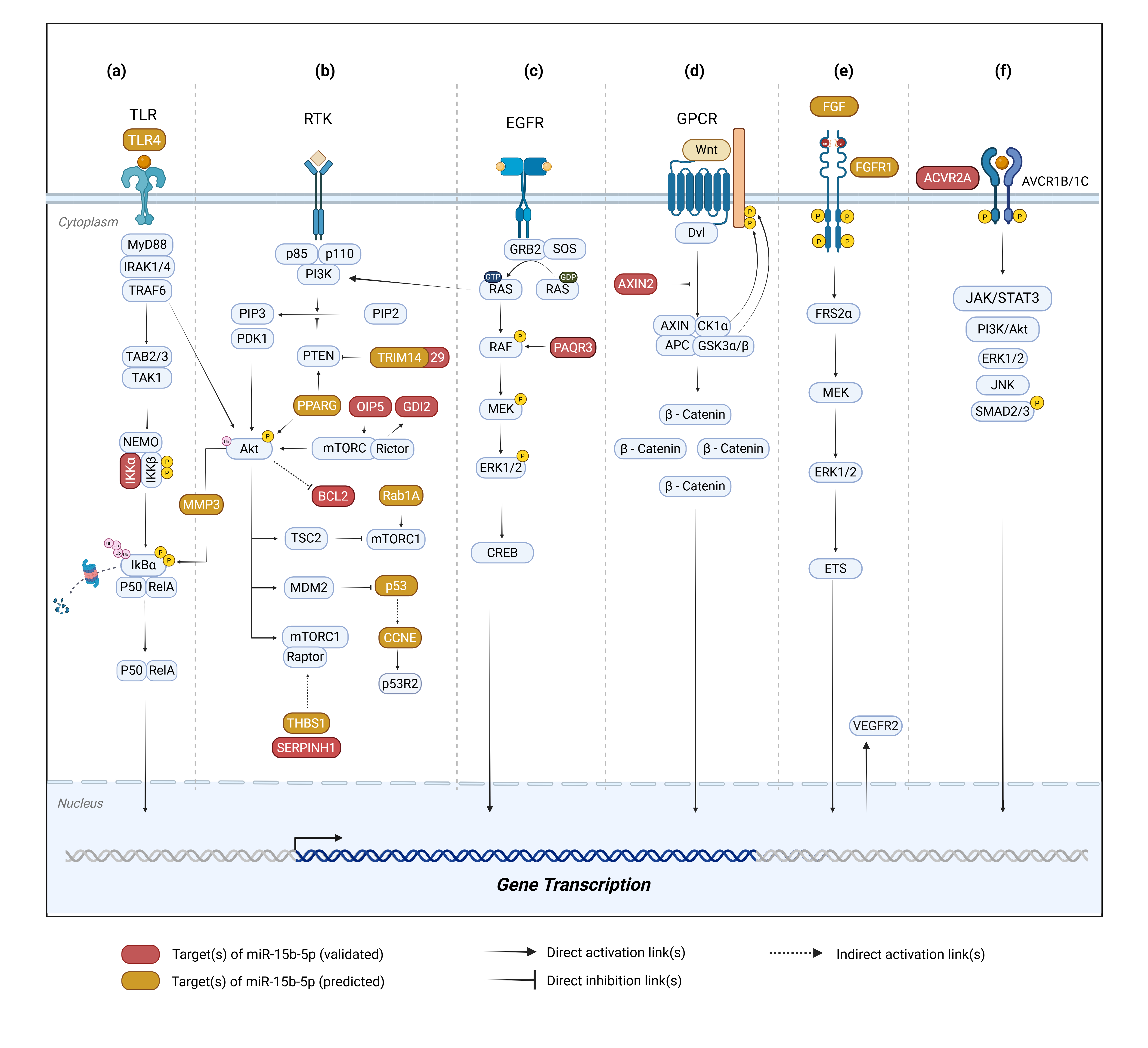
Figure 1 Downstream signaling pathways modulated by miR-15b-5p across multiple cancer-related molecular networks. (a) NF-κB pathway, (b) PI3K/Akt signaling with downstream crosstalk to the p53 regulatory axis, (c) MAPK/ERK pathway, (d) Wnt/β-catenin pathway, (e) VEGF signaling pathway, and (f) TGF-β/SMAD pathway. Created in BioRender. Dewi, D. (2025) https://BioRender.com/eernija.
Table 3 Summary of pathways involved in miR-15b-5p regulation. This highlights the target genes and signaling pathways associated with miR-15b-5p. In addition, miR-15b-5p may function as either an oncogenic miRNA or tumor suppressor depending on the molecular context.
Cancer type |
Sample type |
Methods |
Target genes |
Pathway |
Validation Status |
Potential role of miRNA |
Citation |
PCa |
- Human prostate epithelial cell lines - LNCaP (ATCC, CRL-1740) - PC3 cells |
- qRT-PCR - In cell Western
|
ACVRA2 |
TGF-β |
Validated |
oncomiR ↓ miR-15b-5p in LNCaP cells vs HC ↓ miR-15b-5p ↑ ACVR2A cell cycle arrest occured |
[60] |
PCa |
Data sets of mRNA and miRNA expression and clinical information from the University of California Santa Cruz Xena database |
Bioinformatics |
NUSAP1 and AURKB |
- Sphingosine 1-phosphate - Liver Kinase B1 signaling - TRAIL signaling pathway - ErbB receptor signaling network - IFN-gamma pathway - Alpha9 beta1 integrin signaling |
Predicted |
oncomiR ↑ miR-15b-5p poor prognosis |
[61] |
PCa |
- PCa cell lines (22RV1, PC3) - PCa tissues - Adjacent normal tissues (TCGA data) - Xenograft mouse model |
- qRT-PCR - Western blot - Dual-luciferase reporter assay - In vitro - In vivo
|
RECK |
RECK/ MMP |
Validated |
oncomiR ↓ miR-15b-5p ⟞ RECK ↑ cell growth ↑ invasiveness ↑ tumor recurrence
|
[8] |
HCC |
- HepG2, Hep3B, SKHEP-1, Alexander cells, Chang liver, THLE2, Huh7, HLK2, HKK2, HLK4, HLK1, HLK5, HKK1, HLK6, HLK3, SH-J1 cell lines - BALB/c nude mice (BALB/cByJ-Hfh11nu) |
- In vitro - In vivo - qRT-PCR - Dual-luciferase assay IHC - Immunoblotting |
OIP5 |
AKT/mTORC1 and β-catenin |
Validated |
TSM ↑ miR-15b-5p ↓ OIP5 ↓ tumor growth ↓ metastasis |
[62] |
HCC |
- Liver cancer tissues (n = 69) - Adjacent non-cancerous tissues - Cell lines (HepG2, Huh7, Hep3B, HCCLM3) |
- qRT-PCR - Western blot -Dual-luciferase reporter assay - In vitro - In vivo
|
Axin2 |
Wnt/β‑catenin
|
Validated |
oncomiR ↑ miR-15b-5p ↓ Axin2 ↑ proliferation ↑ invasion ↑ TNM stage ↑ tumor capsular infiltration ↓ overall survival |
[7] |
CRC |
- CRC cell lines (SW620, HCT116, DLD1, SW1116, NCM460) - CRC tissues and adjacent normal mucosa (23 pairs) - CRC patient tissues (TCGA data) - CAC and xenograft mouse model |
- qRT-PCR - Western blot - Dual-luciferase reporter assay - ChIP - IHC - In vitro - In vivo
|
NF-KB1, IKK-α |
NF-KB/XIAP |
Validated
|
TSM ↑ miR-15b-5p ↓ NF-KB1 ↓ IKK-α ↑ miR-15b-5p ⟞ 5-FU resistance ↓ tumor progression
|
[63] |
CRC |
- CRC tissues (n = 94) HCT116, SW480, - SW620, LoVo, Caco-2, HT-29 cell lines - Xenograft mouse model |
- qRT-PCR - Western blot - Luciferase reporter assay - IHC - In vitro - In vivo |
ACOX1 |
Peroxisomal β-oxidation |
Validated |
oncomiR ↑ miR-15b-5p ⟞ ACOX1 ↑ metastasis
|
[9] |
Microsatellite Stable Colorectal Cancer (MSS CRC) |
- MSS CRC tissues (human, n = 160 - 261) - MSS CRC cell lines (CT26, MC38, SW1116, HT29, SW480, SW620); Colitis-Associated Cancer (CAC) - Murine models; Syngeneic subcutaneous CT26 - MC38 tumor-bearing mice |
- qRT-PCR - Luciferase reporter assay - Western blot - ChIP - IHC - In vitro - In vivo
|
PD-L1 |
Immune Response Checkpoint (PD1-PDL1) |
Validated |
TSM ↑ miR-15b-5p ↓ PD-L1
↑ IL-17A
↑ anti-PD-1
resistance IL-17A
blockade |
[64] |
NB |
- SK-N-AS, NB-19, and SK-N-BE, NB975, K562.mbIL21.4-1BBL cell lines co-cultured with CD8+T cell and NK cells - C57/BL6 mice |
- In vitro - In vivo - Western blot - qRT-PCR - Luciferase reporter assay - IHC |
PD-L1 |
Immune Response Checkpoint (PD1-PDL1) |
Validated |
TSM ↓ miR-15b-5p ↑ PD-L1 ↑ TIL exhaustion in NB patients and PDX tumors
|
[65] |
NB |
- NB patient-derived xenografts - NB cell lines (SK-N-BE (2), NB-19, CHLA-136, SH-EP Tet21N) - Xenograft model in NSG mice |
- qRT-PCR - Western blot - Luciferase reporter assay - In vitro - In vivo
|
MYCN |
Post-transcriptional repression (RISC/Ago2/3'UTR) of MYCN |
Validated |
TSM ↑ miR-15b-5p ↓ MYCN ↓ proliferation ↓ migration ↓ invasion ↓ colony formation ↓tumorigenesis |
[66] |
NPC |
- 5-8F, 6-10B, S-18, S-26, CNE1, CNE2 and SUNE2 cell lines - Fresh NPC tissues (n = 25), - HC tissues (n = 17) - FFPE of primary NPC tissues (n = 69) |
- qRT-PCR - Western blot - IHC - In vitro - In vivo - Luciferase reporter assay |
TRIM29 |
PTEN/AKT/mTOR |
Validated |
TSM ↓ miR-15b-5p ↑ TRIM29 ↑ proliferation ↑ EMT ↑ metastasis
|
[67] |
LGG |
- Glioma tissues (n = 5) - Normal cerebral tissues (n = 3) |
- Bioinformatics - Western blot
|
SPTBN2 |
- Neuroactive ligand‑receptor interaction -Cytokine‑cytokine receptor interaction - Calcium signaling pathway - cAMP signaling pathway |
Predicted |
oncomiR ↑ miR-15b-5p ↓ SPTBN2 poor prognosis
|
[68] |
NF- κB pathway
NF-κB is a central transcription factor complex that maintains cellular homeostasis by regulating inflammation, survival, and immune responses. Its dimer remains inactive by binding to IκB until upstream signals trigger IκB phosphorylation and degradation, enabling NF-κB nuclear translocation and promoting gene transcription [69]. Within the NF-κB axis, miR-15b-5p modulates several key components across different cancer types. In HCV-induced hepatocellular carcinoma, miR-15b-5p is predicted to target TLR4, suggesting a potential regulation in inflammatory signaling based on bioinformatic [44]. In colorectal cancer, its downregulation leads to upregulation of IKKα expression, thereby decreasing sensitivity to 5-fluorouracil chemotherapy [63]. Whereas in oral cancer, miR-15b-5p is predicted to target MMP3 and its overexpression enhances cell proliferation, migration and epithelial-mesenchymal transition (EMT) in vitro [41,70].
PI3K/Akt pathway
The PI3K/Akt pathway is one of the major regulators that plays a crucial role in cell survival, proliferation, and metabolism. Activated primarily through receptor tyrosine kinases (RTKs) or G-protein coupled receptors (GPCRs). PI3K activation generates PIP3, which recruits and activates Akt, enabling downstream regulation of targets involved in cell-cycle progression, apoptosis suppression, and motility. Dysregulation of these pathways is common in cancer and frequently associated with enhanced tumor growth and therapeutic resistance [71].
In head and neck squamous cell carcinoma (HNSCC), miR-15b-5p downregulation contributes to the upregulation of TRIM14 and Rab1A. TRIM14 suppresses PTEN, thereby enhancing PI3K/Akt activity and promoting tumor progression. Whereas Rab1A amplification further strengthens proliferative signaling [72]. In nasopharyngeal carcinoma (NPC), elevated TRIM29 similarly promotes proliferation and metastasis by inhibiting PTEN, reinforcing sustained Akt activation [67,73]. In oral cancer, bioinformatic analyses predict that miR-15b-5p targets PPARG [41], a transcription factor that exerts dual effects on Akt signalling, particularly in HNSCC [74]. PPARG is typically elevated in stromal or tumor-adjacent normal cells within the tumor microenvironment (TME), where it can activate PTEN to enhance apoptosis, yet simultaneously stimulates Akt as part of anti-inflammatory and tissue-repair responses. In certain contexts, PPARG can increase chemosensitivity rather than promote tumor growth [74]. More broadly, bioinformatic studies in oral cancer indicate miR-15b-5p is predicted to target SERPINH1 [41]. Furthermore, evidence from colorectal cancer, however, confirms SERPINH1 as a direct miR-15b-5p target, which is associated with advanced T stage, lymph node involvement, distant metastasis, and poor overall survival [75].
In hepatocellular carcinoma (HCC), miR-15b-5p is linked to OIP5-driven Akt activation and increased migration. Thus, suggesting a direct regulatory axis that enhances metastatic behavior through augmented Akt signaling [62]. Moreover, miR-15b-5p directly targets GDI2 in thyroid cancer. Increased GDI2 furthermore induces MMP2/9, leading to enhanced proliferation and invasion [76]. In lung cancer, miR-15b-5p directly represses BCL2 expression, thereby reducing apoptosis and promoting cell viability, migration, and EMT, driven by survival signaling [77]. Collectively, these findings indicate that the PI3K/Akt pathway is more frequently implicated across various cancer types, suggesting its prominent role compared with other signaling pathways.
p53 pathway
The Tumor Protein p53 (p53) axis integrates stress responses to regulate apoptosis, cell-cycle arrest, and DNA repair, and miR-15b-5p intersects with this pathway primarily through genes that regulate proliferation and apoptosis properties [78]. In penile cancer and glioblastoma, profiling data show that tumor protein p53 (TP53), the gene encoding p53, is directly suppressed by miR-15b-5p. This downregulation of p53 leads to reduced apoptosis and enhanced proliferative capacity[40,79]. In HBV-induced HCC, miR-15b-5p represses CCNE, a key regulator of the G1/S transition. This reduced CCNE expression paradoxically associated with a poorer prognosis, reflecting a complex interplay among viral oncogenesis, disrupted p53 signaling, and compensatory alterations in cell-cycle regulators [50].
MAPK pathway
Mitogen-activated protein kinase (MAPK) regulates various cellular processes, including proliferation, differentiation, apoptosis, and stress responses [80]. miR-15b-5p functions as an oncomiR in gastric cancer (GC). Overexpression of miR-15b-5p in GC cells promotes proliferation, migration, invasion, and epithelial-mesenchymal transition (EMT) through direct repression of progestin and adipoQ receptor family member 3 (PAQR3) [13]. PAQR3 functions as a tumor suppressor that constrains cellular proliferation, migration, tumorigenicity, epithelial-mesenchymal transition (EMT), and metastatic progression [13,81]. By binding to Raf-1 and sequestering it within the Golgi apparatus, PAQR3 prevents its translocation to the cytoplasm and thereby blocks the activation of downstream effectors, ultimately suppressing MAPK pathway signaling [82].
Wnt/β-catenin pathway
In the Wnt/β-Catenin pathway, dysregulation of the transcription factor β-catenin significantly contributes to the initiation and progression of cancer [83–85]. miR-15b-5p is known to regulate the Wnt/β-Catenin pathway. Dong et al. [7] reported that miR-15b-5p was overexpressed in liver cancer tissue compared with normal and was validated to target the axis formation inhibitor 2 (Axin2). Axin2 functions as a TSG that attenuates Wnt/β-catenin signaling activity [7,86]. Axin2 serves as a scaffold component of the β-catenin destruction complex under both Wnt-active and Wnt-inactive conditions. Notably, Axin2 is a direct transcriptional target of β-catenin, such that activation of the Wnt/β-catenin cascade leads to a concurrent rise in Axin2 expression. This induction establishes a negative feedback loop that restrains excessive activation of the Wnt/β-catenin pathway [87], positioning Axin2 as a key negative regulator of the pathway. In this context, the downregulation of Axin2 by miR-15b-5p in liver cancer supports its oncomiR role, as suppression of Axin2 facilitates sustained Wnt/β-catenin activation, thereby enhancing tumor cell proliferation and invasion.
VEGF pathway
The VEGF signaling cascade is a principal regulator of tumor angiogenesis via VEGFR2-dependent activation of the PI3K/AKT, Ras-Raf-MEK-ERK, and PLCγ/PKC pathways, promoting endothelial proliferation, migration, and survival [88]. Hypoxia-induced stabilization of HIF-1α further enhances VEGF-A transcription, sustaining pathological angiogenesis, while crosstalk with fibroblast growth factor (FGF) signaling amplifies the angiogenic response [89,90]. In renal cell carcinoma, miR-15b-5p is markedly downregulated and inversely associated with tumor grade, indicating a tumor-suppressive and anti-angiogenic function [31]. Mechanistically, miR-15b-5p has been reported to target FGF ligands and FGFR1/2, which are central upstream regulators of the Fibroblast Growth Factor- Fibroblast Growth Factor Receptor (FGF-FGFR) axis responsible for activating Ras-Raf-MAPK, PI3K-AKT, and PLCγ signaling pathways essential for angiogenesis and endothelial activation. Downregulation of miR-15b-5p may therefore release FGF/FGFR signaling from post-transcriptional repression, consequently amplifying VEGF-mediated angiogenesis through synergistic pathway activation [91].
TGF-β pathway
The TGF-β pathway is initiated when active TGF-β binds to type II receptors, leading to recruitment and phosphorylation of type I receptors and subsequent activation of the canonical SMAD2/3-SMAD4 complex, which translocates to the nucleus to regulate genes controlling proliferation, differentiation, and tissue homeostasis [92,93]. In canonical pathway, TGF-β activates several non-SMAD pathways including MAPK, PI3K/AKT, and Rho GTPase signaling, which control cell migration, epithelial-mesenchymal transition (EMT), and tumor progression. Within the activin subfamily of TGF-β ligands, activin binds to type II receptors such as ACVR2A (Activin A Receptor Type 2A), which subsequently recruit type I receptors (e.g., ACVR1B) to initiate both SMAD-dependent and SMAD-independent signaling cascades [93]. Activin A has been shown to downregulate miR-15b-5p and increases ACVR2A expression in prostate cancer cells, suggesting an inverse regulatory relationship [60]. Upregulation of ACVR2A may therefore enhance activin/TGF-β signaling by phosphorylating SMAD2/3 and activating non-SMAD pathways, promoting proliferation, EMT, apoptosis resistance, and tumor aggressiveness [94].
Future prospect of miR-15b-5p in cancer studies
Defining molecular heterogeneity and the dual roles
Literature profiling reveals the unique behavior of miR-15b-5p, which exhibits a dual role expression pattern determined by tumor origin, molecular subtype, and microenvironment. Although its differential expression has been well documented across various tissues, the specific molecular factors underlying this heterogeneity remain incompletely understood. Research must now focus on identifying the regulatory factors causing this variation, particularly upstream transcription regulators, epigenetic modifications, and non-coding RNA networks (such as the lncRNA-miRNA-mRNA axis). It is also important to conduct pan-cancer and cross-tissue analyses to determine whether this dual role stems from distinct biological programs or from shared regulatory motifs. Limited pan-cancer miRNA analyses have been reported, mainly to identify miRNAs that are consistently upregulated or downregulated across multiple tumor types compared to normal tissues [95]. This study has highlighted miRNAs that act as broad tumor suppressors or oncogenic regulators in either cancer-type–specific or pan-cancer contexts. However, miR-15b-5p has rarely been a primary focus in these investigations and has been only incidentally included in a limited number of studies. Given that miR-15b-5p shows context-dependent expression patterns across different cancers, dedicated pan-cancer and integrated multi-omics analyses centered on this miRNA remain largely unexplored and represent an important direction for future research. Discovering these mechanisms is key to establishing miR-15b-5p as a promising and context-specific biomarker for diagnosis and prognosis.
Clarifying mechanistic networks and pathway intersections
Our pathway-based analysis indicates that miR-15b-5p is involved in multiple cancer processes, from proliferation to apoptosis. Most of these effects converge on the PI3K/Akt pathway, a central hub for cell survival, which interacts extensively with the p53 signaling axis to control the cell cycle. In addition to direct mRNA targeting, miR-15b-5p activity is likely shaped by multilayered regulation involving lncRNA-mediated sponging and epigenetic modulation, which may shift pathway dominance across different tumor contexts and contribute to its context-dependent dual role. Based on the literature we reviewed, upstream lncRNA regulators of miR-15b-5p remain limited and are not yet well characterized, indicating an important gap for further investigation.
While many targets cluster within the PI3K/Akt network, we also identified predicted and validated targets like PAQR3, AXIN2, and ACVR2A. These genes may have had a significant effect because they became key nodes with other major oncogenic pathways, including Wnt/β-catenin, MAPK/ERK, and TGF-β/SMAD. It’s likely to act as context-dependent modulators, broadening the functional impact of miR-15b-5p on specific tumor phenotypes. To validate these connections, future work must prioritize gain- and loss-of-function studies and luciferase reporter assays to verify direct miRNA-mRNA binding. Furthermore, using in vivo models is essential to confirm their physiological relevance. Further understanding of this deeper mechanistic insight would properly evaluate miR-15b-5p’s potential as a candidate for pathway-guided therapy.
Driving translational applications and therapeutic strategies
It is important to distinguish between experimentally validated findings and emerging hypotheses when considering the translational potential of miR-15b-5p. Due to the large number of targets that have not been experimentally validated yet, the interpretation bioinformatics predictions require careful consideration when discussing therapeutic implications. The initial discovery-driven phase should be followed by a more rigorous and systematic validation. Using human miRNA mimics and in vivo xenograft models may serve as a practical starting point for confirming target interactions. This step is essential, particularly for assessing toxicity and bioavailability before any therapy can move toward human clinical trials.
One of the most likely directions of translation at this stage is the potential role of miR-15b-5p in modulating therapeutic resistance. Preclinical evidence indicates that miR-15b-5p may reverse or modulate 5-fluorouracil resistance by disrupting the NF-κB/IKKα/XIAP axis. Some predicted targets, such as PPARG, may also influence the tumor microenvironment, potentially affecting treatment response, although these indirect effects remain to be experimentally confirmed. Taken together, these findings suggest potential directions for future clinical translation, in which modulation of miR-15b-5p could be explored as a potential chemosensitizing strategy to enhance the effectiveness of standard treatment regimens, although current evidence remains largely preclinical.
The potential of miR-15b-5p in immunotherapy is also becoming an increasingly interesting area of exploration. Several reports suggest that this miRNA modulates PD-L1 expression in neuroblastoma and CRC, suggesting a potential immunoregulatory role in specific experimental contexts. In limited preclinical models, delivery of miR-15b-5p mimics has been associated with increased responsiveness to anti-PD-1 therapy by lowering the threshold for immune activation. Even so, the effectiveness of this approach still depends heavily on how the molecule is delivered. This is why the development of nanoparticle- or exosome-based carriers that can protect and safely transport RNA remains a key requirement for advancing miRNA-based immunotherapy toward clinical use.
Overall, these results suggest that miR-15b-5p may be useful for more than just regulating a single pathway, especially in approaches aimed at overcoming treatment resistance. MiR-15b-5p may function as a context-dependent regulator of pathways associated with treatment resistance and immune modulation, as it controls NF-κB signaling, which is involved in chemotherapy resistance to agents such as 5-fluorouracil, and modulates immune-related pathways, including PD-L1 expression and T cell responses [65,96]. These characteristics raise the possibility that miR-15b-5p–targeted approaches could be investigated in combination with immunotherapeutic modalities, including cell-based strategies; Such applications remain theoretical and require substantial preclinical validation. In addition, because RNA molecules are naturally unstable and context-dependent, delivery issues still limit the therapeutic application of miRNAs. Although there are still significant challenges related to safety, specificity, and scalability, delivery platforms such as exosomes or nanoparticle-based carriers have been proposed to improve stability and targeting accuracy. Importantly, any combination strategy requires spatial and temporal control to minimize off-target or oncogenic effects due to the dual nature and context-dependent behavior of miR-15b-5p. This perspective highlights the potential significance of miR-15b-5p in influencing intrinsic tumor signaling and the tumor immune microenvironment, although further mechanistic and translational studies are required.
Conclusions
The context-dependent nature and regulatory complexity of miR-15b-5p across various cancer types indicate that its oncogenic or tumor-suppressive functions are dictated by tumor-specific molecular features and microenvironmental cues. miR-15b-5p biogenesis is also influenced by upstream regulators such as genetic polymorphisms, thereby reshaping mature miRNA expression and potentially conferring downstream regulatory consequences. The role of miR-15b-5p in cancer is further shaped by lncRNAs acting as upstream regulators, whose cancer-specific regulatory patterns govern the context-dependent functions of miR-15b-5p across malignancies. By targeting genes within distinct signaling pathways, miR-15b-5p exerts pathway-specific effects that ultimately determine its tumor-suppressive or oncogenic role in a cancer type-dependent manner. It should also be noted that existing studies exhibit substantial heterogeneity in sample sources, experimental platforms, and patient populations, which may contribute to the inconsistent expression patterns reported across various cancers. Furthermore, many mechanistic insights are derived from preclinical models, highlighting the need for further validation in well-characterized clinical cohorts. Overall, miR-15b-5p is a promising candidate for a non-invasive biomarker and therapeutic target in cancer. However, further integrative and mechanistic studies are essential in order to define the determinants of its divergent roles and advance its use in precision oncology.
Acknowledgements
This work was supported by the Directorate General of Higher Education, Research, and Technology, Ministry of Education, Culture, Research, and Technology, Republic of Indonesia (contract number: 067/C3/DT.05.00/PL/2025;2480/ UN1/DITLIT/Dit-Lit/PT.01.03/2025).
Declaration of Generative AI in Scientific Writing
The authors acknowledge the use of generative AI tools (e.g., Grammarly and ChatGPT by OpenAI) in the preparation of this manuscript, specifically for grammar correction and language editing. AI tools were not utilized for generating scientific content or interpreting research data. The authors remain fully responsible for the accuracy, content, and conclusions presented in this study.
CRediT Author Statement
Kharisma Arethusa Maisaroh, Putri Rohmatul Laili, Reza Adhi Pratama, and Auraga Dewantoro: Contributed equally to this work: Conceptualization; writing the original draft; visualization. Dyah Laksmi Dewi: Conceptualization; writing; review and editing; supervision; funding acquisition. Teguh Aryandono: Supervision; review and editing. Irianiwati Widodo: Supervision; review and editing; funding acquisition.
References
[1] HH Wu, S Leng, C Sergi and R Leng. How MicroRNAs command the battle against cancer. International Journal of Molecular Sciences 2024; 25(11), 5865.
[2] T Tomankova, M Petrek and E Kriegova. Involvement of microRNAs in physiological and pathological processes in the lung. Respiratory Research 2010; 11(1), 159.
[3] MI Almeida, RM Reis and GA Calin. MicroRNA history: Discovery, recent applications, and next frontiers. Mutation Research/Fundamental and Molecular Mechanisms of Mutagenesis 2011; 717(1-2), 1-8.
[4] Y Peng and CM Croce. The role of MicroRNAs in human cancer. Signal Transduction and Targeted Therapy 2016; 1(1), 15004.
[5] J O’Brien, H Hayder, Y Zayed and C Peng. Overview of microRNA biogenesis, mechanisms of actions, and circulation. Frontiers in Endocrinology 2018; 9, 402.
[6] Z Wang, H Wang, S Zhou, J Mao, Z Zhan and S Duan. miRNA interplay: Mechanisms and therapeutic interventions in cancer. MedComm-Oncology 2024; 3(4), e93.
[7] Y Dong, N Zhang, S Zhao, X Chen, F Li and X Tao. miR-221-3p and miR-15b-5p promote cell proliferation and invasion by targeting Axin2 in liver cancer. Oncology Letters 2019; 18(6), 6491-6500.
[8] R Chen, L Sheng, HJ Zhang, M Ji and WQ Qian. miR-15b-5p facilitates the tumorigenicity by targeting RECK and predicts tumour recurrence in prostate cancer. Journal of Cellular and Molecular Medicine 2018; 22(3), 1855-1863.
[9] LN Sun, Z Zhi, LY Chen, Q Zhou, XM Li, WJ Gan, S Chen, M Yang, Y Liu, T Shen, Y Xu and JM Li. SIRT1 suppresses colorectal cancer metastasis by transcriptional repression of miR-15b-5p. Cancer Letters 2017; 409, 104-115.
[10] GR Chen, YB Zhang, SF Zheng, YW Xu, P Lin, HC Shang-Guan, YX Lin, DZ Kang and PS Yao. Decreased SPTBN2 expression regulated by the ceRNA network is associated with poor prognosis and immune infiltration in low-grade glioma. Experimental and Therapeutic Medicine 2023; 25(6), 253.
[11] MTD Martino, P Tagliaferri and P Tassone. MicroRNA in cancer therapy: Breakthroughs and challenges in early clinical applications. Journal of Experimental & Clinical Cancer Research 2025; 44(1), 126.
[12] MJ Ramaiah. Functions and epigenetic aspects of miR-15/16: Possible future cancer therapeutics. Gene Reports 2018; 12, 149-164.
[13] C Zhao, Y Li, G Chen, F Wang, Z Shen and R Zhou. Overexpression of miR-15b-5p promotes gastric cancer metastasis by regulating PAQR3. Oncology Reports 2017; 38(1), 352-358.
[14] S Ghafouri-Fard, T Khoshbakht, BM Hussen, HH Jamal, M Taheri and M Hajiesmaeili. A comprehensive review on function of miR-15b-5p in malignant and non-malignant disorders. Frontiers in Oncology 2022; 12, 870996.
[15] X Jin, Y Chen, H Chen, S Fei, D Chen, X Cai and C Xie. Evaluation of tumor-derived exosomal miRNA as potential diagnostic biomarkers for early-stage non-small cell lung cancer using next-generation sequencing. Clinical Cancer Research 2017; 23(17), 5311-5319.
[16] X Jin, Y Chen, H Chen, S Fei, D Chen, X Cai, L Liu, B Lin, H Su, L Zhao, M Su, H Pan, L Shen, D Xie and C Xie. Identifying specific miRNAs and associated mRNAs in CD44 and CD90 cancer stem cell subtypes in gastric cancer cell line SNU-5. International Journal of Clinical and Experimental Pathology 2020; 13(6), 1313-1323.
[17] X Li, C Sun, Z Ge, Y Li, H Zhou, Y Wu, S Lin, P Zhang, X Wu and Y Lai. Evaluation of the diagnostic value of a three-miRNA panel in prostate cancer: A discovery and validation study. Discover Oncology 2025; 16(1), 611.
[18] E Milanesi, M Dobre, AI Bucuroiu, V Herlea, TE Manuc, A Salvi, GD Petro, M Manuc and G Becheanu. miRNAs-based molecular signature for KRAS mutated and wild type colorectal cancer: An explorative study. Journal of Immunology Research 2020; 2020, 4927120.
[19] S Paydas, A Acikalin, M Ergin, H Celik, B Yavuz and K Tanriverdi. Micro-RNA (miRNA) profile in Hodgkin lymphoma: Association between clinical and pathological variables. Medical Oncology 2016; 33(4), 34.
[20] L Fan, H Qi, J Teng, B Su, H Chen, C Wang and Q Xia. Identification of serum miRNAs by nano-quantum dots microarray as diagnostic biomarkers for early detection of non-small cell lung cancer. Tumor Biology 2016; 37(6), 7777-7784.
[21] J Xia, M Luo, L Dai, L Wang, L Wang and J Zhu. Serum exosomal microRNAs as predictive markers for EGFR mutations in non-small cell lung cancer. Journal of Clinical Laboratory Analysis 2021; 35(5), e23743.
[22] CO Pintea, M Pricop, E Seclaman, NC Balica, K Guran, DI Horhat and CI Mot. A pilot study on the analysis of circulating miRNA upregulation in laryngeal cancer. Diseases 2025; 13(4), 101.
[23] J Gasparello, C Papi, M Allegretti, E Giordani, F Carboni, S Zazza, E Pescarmona, P Romania, P Giacomini, C Scapoli, R Gambari and A Finotti. A distinctive microRNA (miRNA) signature in the blood of colorectal cancer (CRC) patients at surgery. Cancers 2020; 12(9), 2410.
[24] GB Spychalski, AA Lin, SJ Yang, H Shen, J Rosario, K Tien, K French, M Ghali, S Yee, M Yin, MD Feldman, EF Conant, SP Weinstein, EL Carpenter, D Issadore and A Nayak. miRNA panel from HER2+ and CD24+ plasma extracellular vesicle subpopulations as biomarkers of early-stage breast cancer. Breast Cancer Research 2025; 27(1), 90.
[25] M Sromek, G Rymkiewicz, A Paziewska, LM Szafron, M Kulecka, M Zajdel, M Kulinczak, M Dabrowska, A Balabas, Z Bystydzienski, M Chechlinska and JK Siwicki. A set of 17 microRNAs common for brain and cerebrospinal fluid differentiates primary central nervous system lymphoma from non-malignant brain tumors. Biomolecules 2021; 11(9), 1395.
[26] T Sugai, M Osakabe, T Niinuma, R Sugimoto, M Eizuka, Y Tanaka, N Yanagawa, K Otsuka, A Sasaki, T Matsumoto and H Suzuki. Genome-wide analysis of microRNA and mRNA expression in colorectal intramucosal neoplasia and colorectal cancer with a microsatellite-stable phenotype based on adenoma-carcinoma sequences. Frontiers in Oncology 2022; 12, 831100.
[27] R Weissman, EL Diamond, J Haroche, N Pillar, G Shapira, BH Durham, J Buthorn, F Cohen, M Ki and O Hershkovitz-Rokah. The contribution of microRNAs to the inflammatory and neoplastic characteristics of Erdheim-Chester disease. Cancers 2020; 12(11), 3240.
[28] DA Armstrong, BB Green, JD Seigne, AR Schned and CJ Marsit. MicroRNA molecular profiling from matched tumor and bio-fluids in bladder cancer. Molecular Cancer 2015; 14(1), 194.
[29] A Molfino, M Beltra, MI Amabile, R Belli, G Birolo, E Belloni, S De Lucia, L Garcia-Castillo, F Penna, G Imbimbo, G Nigri, B Pardini, P Costelli and M Muscaritoli. Small non-coding RNA profiling in patients with gastrointestinal cancer. Journal of Cachexia, Sarcopenia and Muscle 2023; 14(6), 2692-2702.
[30] W Xu, S Liu, L Ma, L Cheng, Q Li, L Qing, Y Yang and Z Dong. Identification of miRNA signature in cancer-associated fibroblast to predict recurrent prostate cancer. Computers in Biology and Medicine 2024; 180, 108989.
[31] J Kiraly, E Szabo, P Fodor, A Vass, M Choudhury, R Gesztelyi, C Szasz, T Flasko, N Dobos, B Zsebik, AJ Steli, G Halmos and Z Szabo. Expression of hsa-miRNA-15b,-99b,-181a and their relationship to angiogenesis in renal cell carcinoma. Biomedicines 2024; 12(7), 1441.
[32] M Marcuello, S Duran-Sanchon, L Moreno, JJ Lozano, L Bujanda, A Castells and M Gironella. Analysis of a 6-miRNA signature in serum from colorectal cancer screening participants as non-invasive biomarkers for advanced adenoma and colorectal cancer detection. Cancers 2019; 11(10), 1542.
[33] J Mills, M Capece, E Cocucci, A Tessari and D Palmieri. Cancer-derived extracellular vesicle-associated microRNAs in intercellular communication: one cell's trash is another cell's treasure. International Journal of Molecular Sciences 2019; 20(24), 6109.
[34] M Groot and H Lee. Sorting mechanisms for microRNAs into extracellular vesicles and their associated diseases. Cells 2020; 9(4), 1044.
[35] B Zhang and Q Ye. Linc00662 sponges miR-15b-5p to promote hypopharyngeal squamous cell carcinoma progression by facilitating cancer stem cell-like phenotypes. Journal of Cancer 2024; 15(12), 3781.
[36] Y Ashirbekov, N Khamitova, K Satken, A Abaildayev, I Pinskiy, A Yeleussizov, L Yegenova, A Kairanbayeva, D Kadirshe, G Utegenova, N Jainakbayev and K Sharipov. Circulating microRNAs as biomarkers for the early diagnosis of lung cancer and its differentiation from tuberculosis. Diagnostics 2024; 14(23), 2684.
[37] S Fogli, B Polini, S Carpi, B Pardini, A Naccarati, N Dubbini, M Lanza, MC Breschi, A Romanini and P Nieri. Identification of plasma microRNAs as new potential biomarkers with high diagnostic power in human cutaneous melanoma. Tumor Biology 2017; 39(5), 1-8.
[38] D Hanniford, J Zhong, L Koetz, A Gaziel-Sovran, DJ Lackaye, S Shang, A Pavlick, R Shapiro, R Berman, F Darvishian, Y Shao, I Osman and E Hernando. A miRNA-based signature detected in primary melanoma tissue predicts development of brain metastasis. Clinical Cancer Research 2015; 21(21), 4903-4912.
[39] CY Li, WW Zhang, JL Xiang, XH Wang, J Li and JL Wang. Identification of microRNAs as novel biomarkers for esophageal squamous cell carcinoma: A study based on The Cancer Genome Atlas (TCGA) and bioinformatics. Chinese Medical Journal 2019; 132(18), 2213-2222.
[40] J da Silva, CC da Costa, IF Ramos, AC Laus, L Sussuchi, RM Reis, AS Khayat, LR Cavalli and SR Pereira. Upregulated miRNAs on the TP53 and RB1 binding seedless regions in high-risk HPV-associated penile cancer. Frontiers in Genetics 2022; 13, 875939.
[41] Y Yang, X Xin and X Xu. Identification of potential miRNA-mRNA regulatory network in the development of oral cancer. Disease Markers 2022; 2022, 9376608.
[42] Y Zhao, Y Song, Y Zhang, M Ji, P Hou and F Sui. Screening protective miRNAs and constructing novel lncRNAs/miRNAs/mRNAs networks and prognostic models for triple-negative breast cancer. Molecular and Cellular Probes 2023; 72, 101940.
[43] G Cochetti, L Cari, V Maula, R Cagnani, A Paladini, M Del Zingaro, G Nocentini and E Mearini. Validation in an independent cohort of MiR-122, MiR-1271, and MiR-15b as urinary biomarkers for the potential early diagnosis of clear cell renal cell carcinoma. Cancers 2022; 14(5), 1112.
[44] AA Mohamed, NN Amer, N Osama, W Hafez, AEA Ali, MM Shaheen, AA Alkhalegy, EA Abouahmed, SM Soaida, LA Samy, A El-Kassas, I Cherrez-Ojeda and RR El-Awady. Expression of miR-15b-5p and toll-like receptor4 as potential novel diagnostic biomarkers for hepatitis C virus-induced hepatocellular carcinoma. Non-Coding RNA Research 2025; 10, 262-268.
[45] M Dobre, RC Poenaru, AM Niculae, C Vladut, V Herlea, E Milanesi and ME Hinescu. Increased levels of miR-15b-5p and miR-20b-5p in pancreatic ductal adenocarcinoma with hepatic metastases. Genes 2023; 14(8), 1577.
[46] R Duan, C Pak and P Jin. Single nucleotide polymorphism associated with mature miR-125a alters the processing of pri-miRNA. Human Molecular Genetics 2007; 16(9), 1124-1131.
[47] R Harnprasopwat, D Ha, T Toyoshima, H Lodish, A Tojo and A Kotani. Alteration of processing induced by a single nucleotide polymorphism in pri-miR-126. Biochemical and Biophysical Research Communications 2010; 399(2), 117-122.
[48] W Sun, J Lan, L Chen, J Qiu, Z Luo, M Li, J Wang, J Zhao, T Zhang, X Long, J Chai, Z Yan, Z Guo and S Gun. A mutation in porcine pre-miR-15b alters the biogenesis of miR-15b\16-1 cluster and strand selection of miR-15b. PLoS One 2017; 12(5), e0178045.
[49] JS Mattick, PP Amaral, P Carninci, S Carpenter, HY Chang, LL Chen, R Chen, C Dean, ME Dinger, KA Fitzgerald, TR Gingeras, M Guttman, T Hirose, M Huarte, R Johnson, C Kanduri, P Kapranov, JB Lawrence, JT Lee, …, M Wu. Long non-coding RNAs: Definitions, functions, challenges and recommendations. Nature Reviews Molecular Cell Biology 2023; 24(6), 430-447.
[50] J Xu, J Zhang, F Shan, J Wen and Y Wang. SSTR5-AS1 functions as a ceRNA to regulate CA2 by sponging miR-15b-5p for the development and prognosis of HBV-related hepatocellular carcinoma. Molecular Medicine Reports 2019; 20(6), 5021-5031.
[51] F Wang, Y Zu, S Zhu, Y Yang, W Huang, H Xie and Y Zu. Long noncoding RNA MAGI2-AS3 regulates CCDC19 expression by sponging miR-15b-5p and suppresses bladder cancer progression. Biochemical and Biophysical Research Communications 2018; 507, 231-235.
[52] B Ma, S Wang, W Wu, P Shan, Y Chen, J Meng, L Xing, J Yun, L Hao, X Wang, S Li and Y Guo. Mechanisms of circRNA/lncRNA-miRNA interactions and applications in disease and drug research. Biomedicine & Pharmacotherapy 2023; 162, 114672.
[53] S Asai, Y Goto, K Tanigawa, Y Tomioka, M Kato, K Mizuno, S Sakamoto and N Seki. MiR-15b-5p inhibits castration-resistant growth of prostate cancer cells by targeting the muscarinic cholinergic receptor CHRM3. FEBS Letters 2023; 597(8), 1164-1175.
[54] Y Cai, Y Yang, X Zhang, Q Ma and M Li. TRPM2-AS promotes the malignancy of osteosarcoma cells by targeting miR-15b-5p/PPM1D axis. Cell Cycle 2022; 21(8), 835-850.
[55] SY Zhao, Z Wang, XB Wu, S Zhang, Q Chen, DD Wang and QF Tan. CERS6-AS1 contributes to the malignant phenotypes of colorectal cancer cells by interacting with miR-15b-5p to regulate SPTBN2. The Kaohsiung Journal of Medical Sciences 2022; 38(5), 403-414.
[56] Y Zhou, RG Fan, CL Qin, J Jia, XD Wu and WZ Zha. LncRNA-H19 activates CDC42/PAK1 pathway to promote cell proliferation, migration and invasion by targeting miR-15b in hepatocellular carcinoma. Genomics 2019; 111(6), 1862-1872.
[57] Z Li, JM Tian, Y Chu, HY Zhu, JJ Wang and J Huang. Long non-coding RNA PVT1 (PVT1) affects the expression of CCND1 and promotes doxorubicin resistance in osteosarcoma cells. Journal of Bone Oncology 2023; 43, 100512.
[58] A Tolle, L Buckendahl and K Jung. Plasma miR-15b-5p and miR-590-5p for distinguishing patients with bladder cancer from healthy individuals. Oncology Reports 2019; 42(4), 1609-1620.
[59] Y Zhu, X Zhang, L Wang, X Zhu, Z Xia, L Xu and J Xu. FENDRR suppresses cervical cancer proliferation and invasion by targeting miR-15a/b-5p and regulating TUBA1A expression. Cancer Cell International 2020; 20(1), 152.
[60] EC Ottley, HD Nicholson and EJ Gold. Activin A regulates microRNAs and gene expression in LNCaP cells. The Prostate 2016; 76(11), 951-963.
[61] NB Zhang, JJ Jin and XH Ye. Integrated analysis of the potential roles of microRNA-messenger RNA networks in prostate adenocarcinoma. Heliyon 2023; 9(12), e22383.
[62] H Li, J Zhang, MJ Lee, GR Yu, X Han and DG Kim. OIP5, a target of miR-15b-5p, regulates hepatocellular carcinoma growth and metastasis through the AKT/mTORC1 and β-catenin signaling pathways. Oncotarget 2017; 8(11), 18129.
[63] C Zhao, Q Zhao, C Zhang, G Wang, Y Yao, X Huang, F Zhan, Y Zhu, J Shi, J Chen, F Yan and Y Zhang. miR-15b-5p resensitizes colon cancer cells to 5-fluorouracil by promoting apoptosis via the NF-kB/XIAP axis. Scientific Reports 2017; 7(1), 4194.
[64] C Liu, R Liu, B Wang, J Lian, Y Yao, H Sun, C Zhang, L Fang, X Guan, J Shi, S Han, F Zhan, S Luo, Y Yao, T Zheng and Y Zhang. Blocking IL-17A enhances tumor response to anti-PD-1 immunotherapy in microsatellite stable colorectal cancer. Journal for Immunotherapy of Cancer 2021; 9(1), e001895.
[65] AS Pathania, P Prathipati, OA Olwenyi, S Chava, OVS Smith, SC Gupta, NK Chaturvedi, SN Byrareddy, DW Coulter and KB Challagundla. miR-15a and miR-15b modulate natural killer and CD8+T-cell activation and anti-tumor immune response by targeting PD-L1 in neuroblastoma. Molecular Therapy-Oncolytics 2022; 25, 308-329.
[66] S Chava, CP Reynolds, AS Pathania, S Gorantla, LY Poluektova, DW Coulter, SC Gupta, MK Pandey and KB Challagundla. miR-15a-5p, miR-15b-5p, and miR-16-5p inhibit tumor progression by directly targeting MYCN in neuroblastoma. Molecular Oncology 2020; 14(1), 180-196.
[67] XM Zhou, R Sun, DH Luo, J Sun, MY Zhang, MH Wang, Y Yang, HY Wang and SJ Mai. Upregulated TRIM29 promotes proliferation and metastasis of nasopharyngeal carcinoma via PTEN/AKT/mTOR signal pathway. Oncotarget 2016; 7(12), 13634.
[68] GR Chen, YB Zhang, SF Zheng, YW Xu, P Lin, HC Shang-Guan, YX Lin, DZ Kang and PS Yao. Decreased SPTBN2 expression regulated by the ceRNA network is associated with poor prognosis and immune infiltration in low-grade glioma. Experimental and Therapeutic Medicine 2023; 25(6), 253.
[69] H Mao, X Zhao and SC Sun. NF-kB in inflammation and cancer. Cellular & Molecular Immunology 2025; 22(8), 811-839.
[70] Y Zhu, L Yan, W Zhu, X Song, G Yang and S Wang. MMP2/3 promote the growth and migration of laryngeal squamous cell carcinoma via PI3K/Akt-NF-kB-mediated epithelial-mesenchymal transformation. Journal of Cellular Physiology 2019; 234(9), 15847-15855.
[71] Y He, MM Sun, GG Zhang, J Yang, KS Chen, WW Xu and B Li. Targeting PI3K/Akt signal transduction for cancer therapy. Signal Transduction and Targeted Therapy 2021; 6(1), 425.
[72] P Ahmad, J Sana, M Slavik, D Gurin, L Radova, NA Gablo, T Kazda, P Smilek, Z Horakova, B Gal, M Hermanova, P Slampa and O Slaby. MicroRNA-15b-5p predicts locoregional relapse in head and neck carcinoma patients treated with intensity-modulated radiotherapy. Cancer Genomics & Proteomics 2019; 16(2), 139-146.
[73] Q Wu, D Nandi and D Sharma. TRIM-endous functional network of tripartite motif 29 (TRIM29) in cancer progression and beyond. Cancer and Metastasis Reviews 2025; 44(1), 16.
[74] B Han, J Chen, S Chen, X Shen, L Hou, J Fang and M Lian. PPARG and the PTEN-PI3K/AKT signaling axis may cofunction in promoting chemosensitivity in hypopharyngeal squamous cell carcinoma. PPAR Research 2024; 2024(1), 2271214.
[75] XS Jin, LX Chen, TT Ji and RZ Li. SERPINH1 promoted the proliferation and metastasis of colorectal cancer by activating PI3K/Akt/mTOR signaling pathway. World Journal of Gastrointestinal Oncology 2024; 16(5), 1890.
[76] J Zou, J Qian, H Fu, F Yin, W Zhao and L Xu. MicroRNA-15b-5p exerts its tumor repressive role via targeting GDI2: A novel insight into the pathogenesis of thyroid carcinoma. Molecular Medicine Reports 2020; 22(4), 2723-2732.
[77] J Wang, S Yao, Y Diao, Y Geng, Y Bi and G Liu. miR-15b enhances the proliferation and migration of lung adenocarcinoma by targeting BCL2. Thoracic Cancer 2020; 11(6), 1396-1405.
[78] M Gasco, S Shami and T Crook. The p53 pathway in breast cancer. Breast Cancer Research 2002; 4(2), 70.
[79] J Gasparello, C Papi, M Zurlo, L Gambari, A Rozzi, A Manicardi, R Corradini, R Gambari and A Finotti. Treatment of human glioblastoma U251 cells with sulforaphane and a peptide nucleic acid (PNA) targeting miR-15b-5p: Synergistic effects on induction of apoptosis. Molecules 2022; 27(4), 1299.
[80] YJ Guo, WW Pan, SB Liu, ZF Shen, Y Xu and LL Hu. ERK/MAPK signalling pathway and tumorigenesis. Experimental and Therapeutic Medicine 2020; 19(3), 1997-2007.
[81] X Yu, Z Li, MTV Chan and WWK Wu. PAQR3: A novel tumor suppressor gene. American Journal of Cancer Research 2015; 5(9), 2562-2568.
[82] Q Guo, XL Liu, K Zhai, C Chen, XX Ke, J Zhang and G Xu. The emerging roles and mechanisms of PAQR3 in human cancer: Pathophysiology and therapeutic implications. International Journal of General Medicine 2023; 16, 4321-4328.
[83] K He and WJ Gan. Wnt/β-catenin signaling pathway in the development and progression of colorectal cancer. Cancer Management and Research 2023; 15, 435-448.
[84] BD White, AJ Chien and DW Dawson. Dysregulation of Wnt/β-catenin signaling in gastrointestinal cancers. Gastroenterology 2012; 142(2), 219-232.
[85] Y Zhang and X Wang. Targeting the Wnt/β-catenin signaling pathway in cancer. Journal of Hematology & Oncology 2020; 13(1), 165.
[86] E Jho, T Zhang, C Domon, CK Joo, JN Freund and F Costantini. Wnt/β-catenin/Tcf signaling induces the transcription of Axin2, a negative regulator of the signaling pathway. Molecular and Cellular Biology 2002; 22(4), 1172-1183.
[87] DB Bernkopf, M Bruckner, MV Hadjihannas and J Behrens. An aggregon in conductin/axin2 regulates Wnt/β-catenin signaling and holds potential for cancer therapy. Nature Communications 2019; 10(1), 4251.
[88] M Simons, E Gordon and L Claesson-Welsh. Mechanisms and regulation of endothelial VEGF receptor signalling. Nature Reviews Molecular Cell Biology 2016; 17(10), 611-625.
[89] H Mahaki, S Nobari, H Tanzadehpanah, A Babaeizad, G Kazemzadeh, M Mehrabzadeh, A Valipour, N Yazdinezhad, H Manoochehri, P Yang and M Sheykhhasan. Targeting VEGF signaling for tumor microenvironment remodeling and metastasis inhibition: Therapeutic strategies and insights. Biomedicine & Pharmacotherapy 2025; 186, 118023.
[90] Z Shi, M Kuai, B Li, CF Akowuah, Z Wang, Y Pan, M Tang, X Yang and P Lu. The role of VEGF in cancer angiogenesis and tumorigenesis: Insights for anti-VEGF therapy. Cytokine 2025; 189, 156908.
[91] G Liu, T Chen, Z Ding, Y Wang, Y Wei and X Wei. Inhibition of FGF-FGFR and VEGF-VEGFR signalling in cancer treatment. Cell Proliferation 2021; 54(4), e13009.
[92] J Liao, R Chen, B Lin, R Deng, Y Liang, J Zeng, S Ma and X Qiu. Cross-talk between the TGF-β and cell adhesion signaling pathways in cancer. International Journal of Medical Sciences 2024; 21(7), 1307.
[93] R Du, L Wen, M Niu, L Zhao, X Guan, J Yang, C Zhang and H Liu. Activin receptors in human cancer: Functions, mechanisms, and potential clinical applications. Biochemical Pharmacology 2024; 222, 116061.
[94] FA Alabi, IF Okpalanwaka, A Oyegbesan, E Okoyeocha, M Oladejo and SO Imodoye. Transforming growth factor-beta signaling in cancer: Therapeutic implications, challenges, and pathways to progress. MedComm-Oncology 2025; 4(3), e70027.
[95] S Moradi, A Kamal, H Aboulkheyr Es, F Farhadi, M Ebrahimi, H Chitsaz, A Sharifi-Zarchi and H Baharvand. Pan-cancer analysis of microRNA expression profiles highlights microRNAs enriched in normal body cells as effective suppressors of multiple tumor types: A study based on TCGA database. PLoS One 2022; 17(4), e0267291.
[96] JD Gagnon, R Kageyama, HM Shehata, MS Fassett, DJ Mar, EJ Wigton, K Johansson, AJ Litterman, P Odorizzi, D Simeonov, BJ Laidlaw, M Panduro, S Patel, LT Jeker, ME Feeney, MT McManus, A Marson, M Matloubian, S Sanjabi and KM Ansel. miR-15/16 restrain memory T cell differentiation, cell cycle, and survival. Cell Reports 2019; 28(8), 2169-2181.