Trends Sci. 2026; 23(10): 13192
Antidiabetic and Renoprotective Potential of Sand Sea Cucumber (Holothuria Scabra) Extract in Streptozotocin-Induced Male Rats
Ahmad
Fajri1,
![]() ,
Amiroh Rochimah Imam1,
,
Amiroh Rochimah Imam1,
![]() ,
Thy Ha Huynh2,
,
Thy Ha Huynh2,
Hien
Minh Nguyen3,
![]() and
Yulia
Yusrini Djabir4,*
and
Yulia
Yusrini Djabir4,*
![]()
1Postgraduate Program, Faculty of Pharmacy, Hasanuddin University, Makassar 90245, Indonesia
2Faculty of Pharmacy, Ton Duc Thang University, Ho Chi Minh City, Vietnam
3Research Group in Pharmaceutical and Biomedical Sciences, Faculty of Pharmacy, Ton Duc Thang University,
Ho Chi Minh City 758307, Vietnam
4Drug Efficacy and Medication Safety Research Group, Faculty of Pharmacy, Hasanuddin University,
Makassar 90245, Indonesia
(*Corresponding author’s e-mail: [email protected])
Received: 29 December 2025, Revised: 16 February 2026, Accepted: 26 February 2026, Published: 20 April 2026
Abstract
Introduction: Chronic hyperglycemia in diabetes mellitus promotes progressive renal injury leading to diabetic nephropathy. This study evaluated the antidiabetic and renoprotective effects of the ethanolic extract of Holothuria scabra in streptozotocin-induced diabetic rats. Methods: Twenty-five male rats were divided into five groups (n = 5). Diabetes was induced in the rats with streptozotocin (55 mg/kg), and then treated orally for 28 days with H. scabra ethanolic extract (200 or 400 mg/kg) or metformin. Fasting blood glucose, serum creatinine and urea levels, and histopathological changes in pancreatic and renal tissues were evaluated. Results: Diabetic rats developed sustained hyperglycemia, weight loss, and marked renal structural damage. Treatment with H. scabra ethanolic extract at 200 mg/kg significantly reduced fasting blood glucose (p < 0.001), serum creatinine (p < 0.05), and renal histological injury scores (p < 0.001) compared with untreated diabetic controls. Pancreatic islet morphology showed partial improvement; however, islet number and area were not significantly restored. Phytochemical screening identified flavonoids, saponins, and steroidal compounds. Conclusion: The ethanolic extract of H. scabra exerts integrated antidiabetic and renoprotective effects at 200 mg/kg dose. These benefits likely result from functional metabolic regulation rather than structural regeneration of pancreatic islets. The findings highlight H. scabra as a promising marine-derived candidate for adjunctive diabetes management. Further mechanistic and dose-optimization investigations are warranted.
Keywords: Holothuria scabra, Diabetes mellitus, Diabetic nephropathy, Pancreatic islets, Kidney histology, Streptozotocin
Introduction
Diabetes mellitus (DM) is a common metabolic disorder, characterized by significantly reduced insulin secretion and the presence of insulin resistance in peripheral tissues [1]. The global prevalence of diabetes in 2024 was estimated at 588.7 million, and this figure is projected to rise to 852.5 million by 2050 [2]. Diabetic nephropathy (DN) is known as a kidney disease caused by diabetes, which is a severe microvascular complication of DM and one of the leading causes of end-stage renal disease (ESRD), contributing to increased morbidity and mortality. The disease develops progressively as chronic hyperglycemia triggers structural and functional alterations in kidney tissues [3,4].
DN is a multifactorial condition that results in a complex interaction between immune-inflammatory responses and metabolic disturbances. Kidney damage is triggered by hyperglycemia through various mechanisms, including increased oxidative stress, accumulation of advanced glycation end products (AGEs), and alterations in renal hemodynamics. These mechanisms induce characteristic pathological changes in DN, such as glomerulosclerosis and tubulointerstitial fibrosis [5]. In addition, the activation of vasoactive hormones, including the endothelin and renin angiotensin systems, also contributes to the progression of DN by causing an increase in intraglomerular and systemic pressure [6]. However, the etiology and pathogenesis of diabetic nephropathy, which involve multiple contributing factors, remain incompletely understood [7].
Bioactive compounds derived from marine organisms have attracted increasing attention as promising sources of novel therapeutic agents. Marine natural products possess unique chemical diversity and exhibit considerable pharmacological potential, particularly in enhancing cellular defense mechanisms through the modulation of oxidative stress and inflammatory pathways. Among marine resources widely distributed in Asian coastal regions, especially in China, Holothuria scabra (Sand sea cucumber) has been recognized for both its high nutritional value and diverse biological activities [8-10]. H. scabra is one of the most extensively studied sea cucumber species (Echinodermata: Holothuroidea), owing to its broad geographic distribution and historically high abundance in muddy coastal habitats throughout the Indo-Pacific region. In addition to its high economic importance as a dried seafood commodity in Asian markets, H. scabra is rich in bioactive secondary metabolites, including triterpenoid glycosides such as bivittoside, echinoside A, frondoside A, holothurin B, and magnumosides. These compounds have been widely reported to exert potent anti-inflammatory and antioxidant activities, highlighting the therapeutic potential of H. scabra in the management of metabolic and inflammatory disorders [11,12].
However, despite increasing evidence supporting the antidiabetic and cytoprotective properties of H. scabra, its effectiveness in preventing or attenuating the progression of diabetic nephropathy has not been extensively investigated. Streptozotocin-induced diabetes is a well-established experimental model that mimics hyperglycemia-associated pancreatic β-cell dysfunction and subsequent diabetic complications, including renal injury [13,14]. Therefore, the present study was designed to evaluate the antidiabetic and renoprotective effects of ethanolic extract of H. scabra in streptozotocin-induced diabetic male rats, with particular emphasis on glycemic control and histopathological alterations in pancreatic and renal tissues.
Materials and methods
Drugs and chemicals
Streptozotocin (BioWorld®, USA), metformin (Hexpharm Jaya, Indonesia), distilled water (OneMed, Indonesia), NaCl 0.9% (Otsuka, Indonesia), ethanol 96% (Medika, Indonesia).
Holothuria scabra extract preparation
Samples of H. scabra were collected from the Center for Environmental Education in Puntondo village, Takalar Regency, South Sulawesi, Indonesia (-5.587862965757303”N and 119.48679043070952”E). Species identification was carried out at the Zoology Laboratory, Department of Biology, Faculty of Mathematics and Natural Sciences, Hasanuddin University, Indonesia (Identification No. 001/ZOO/BIO/2025). After collection, the 8 kg of samples were thoroughly washed, and the visceral organs were removed. The body walls were cut into small pieces and macerated in 1,000 mL of 96% ethanol for three days, with the solvent replaced every 24 h. The resulting macerate was filtered using Whatman No. 1 filter paper, and the filtrate was concentrated using a rotary evaporator at 40 °C to obtain 1.34 kg of ethanolic extract, after which the extraction yield was calculated using the following formula [15].
Yield (%) = (Weight of extract (kg)) / (Weight of sample (kg))×100 (1)
Phytochemical screening
Phytochemical testing was performed to identify the presence of several classes of secondary metabolites in the ethanolic extract of H. scabra. The flavonoid test was conducted by adding magnesium and five drops concentrated HCl to 3 mL of the extract, and the result was considered positive when the solution changed yellow, orange, and red color [16-18]. The tannin test was conducted by adding five drops of 5% ferric chloride (FeCl₃) to 3 mL of the extract, and the formation of a greenish black color indicated the presence of tannins [16]. The saponin test was performed by mixing 5 mL of the extract with 5 mL of water, followed by vigorous vertical shaking and the formation of stable foam with a height of 1 - 10 cm lasting for 15 - 60 min indicated a positive result [18]. The triterpenoid/steroid test was performed by adding a mixture of 1 mL chloroform, 2 mL acetic acid, and 2 drops of sulfuric acid (H₂SO₄) to 1 mL of extract. A color change to purple, green or blue color indicates the presence of triterpenoid compounds [16,17,19]. Triterpenoid/steroid test applied the Carr-Price method involves adding a solution of antimony trichloride (SbCl₃) in chloroform to the extract. Cardiac glycosides identification used the Keller-Killiani test method by adding a mixture of 2 mL glacial acetic acid, 1 drop of FeCl3, and 1 mL H2SO4 to 5 mL of extract. Brown rings form, or purple rings form under the brown rings. In addition, Legal's test method involves dissolving the extract with pyridine, then adding a few drops of sodium nitroprusside solution and sodium hydroxide (NaOH). If a pink or red color appears, it indicates a positive result for glycosides. Cardiac glycoside testing also used the Baljet test method by adding several drops of Baljet reagent (a mixture of 95 mL of 1% picric acid with 5 mL of 10% sodium hydroxide) to the extract. A positive result is indicated by the appearance of an orange or reddish orange color. The alkaloid test was performed by adding three drops each of Mayer, Wagner, and Dragendorff reagents to 2 mL of the extract. The formation of brownish-yellow precipitate (Mayer), reddish brown precipitate (Wagner), and brownish or yellowish precipitate (Dragendorff) indicated positive results [17].
Animals
A total of 25 male Wistar rats (body weight > 200 g) were maintained under standard laboratory conditions with controlled temperature and humidity, a 12-hour light/dark cycle, and free access to food and water. The sample size (n = 5 per group) was selected based on previous studies using comparable experimental designs [20]. Male rats were used to ensure consistent streptozotocin-induced hyperglycemia and to minimize hormonal variability [13]. All experimental procedures were approved by the Ethics Committee of the Faculty of Pharmacy, Hasanuddin University (UNHAS), Indonesia (Approval No. 2052/UN4.17/KP.06.05/2025), and were conducted in accordance with internationally recognized ARRIVE guidelines for the care and use of laboratory animals.
Induction of diabetes
Diabetes was induced by a single intraperitoneal injection of freshly prepared Streptozotocin (STZ) (55 mg/kg) dissolved in 0.1 M citrate buffer (pH 4.5). Following STZ administration, the rats were given 5% glucose solution for 24 h. Subsequently, on day 3 post-STZ induction, blood samples were collected from the tail vein to measure fasting blood glucose levels. Rats with fasting glucose levels > 250 mg/dL accompanied by signs of polyuria and polydipsia were classified as diabetic and included in the study [14,21]. Administration of ethanolic extract of H. scabra was initiated after confirmation of the diabetic condition.
Experimental protocols
Following acclimatization, rats were assigned identification codes and randomly allocated into five groups (n = 5 per group) by manual random selection of the codes. The normal group received 1% sodium carboxymethyl cellulose (Na-CMC) orally. Diabetes was induced in the remaining groups by a single intraperitoneal injection of STZ at a dose of 55 mg/kg. Following diabetes induction, the diabetic group was administered 1% Na-CMC orally. The treatment groups received the ethanolic extract of H. scabra, suspended in 1% Na-CMC, at doses of 200 or 400 mg/kg body weight, administered orally once daily. The doses of the ethanolic extract of H. scabra were selected based on previous studies demonstrating their efficacy and safety on renal function in rats [22]. The metformin group received metformin at a dose of 100 mg/kg orally. All treatments are administered for 28 days. At the end of the experimental period, rats were anesthetized with diethyl ether prior to blood collection. Subsequently, euthanasia was performed using an overdose of diethyl ether in accordance with the approved institutional animal ethics protocol (Approval No. 2052/UN4.17/KP.06.05/2025). The kidney and pancreas tissues were then immediately collected, rinsed with physiological saline solution, and processed for histopathological examination.
Biochemical analysis
Blood samples were collected from rats under light anesthesia induced by diethyl ether to minimize stress and pain during the procedure. Retro-orbital blood collection was performed by trained personnel. Following collection, blood was placed into plain tubes (Onemed, Indonesia) and centrifuged at 4,000 rpm for 15 min to obtain serum for biochemical analysis. Creatinine and urea levels were measured in duplicate using a Humalyzer 3500 clinical chemistry analyzer (Human Diagnostics).
Histopathological examination
The kidneys and pancreas were rinsed with 0.9% physiological saline, and relative organ weight (kidney and pancreatic index) was calculated using the following formula [23].
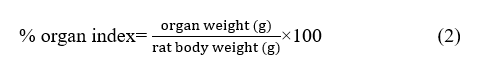
The organs were then fixed in 10% neutral buffered formalin, trimmed into approximately 1 cm sections, and subjected to routine histological processing. Paraffin-embedded tissues were sectioned at a thickness of 4 µm using a microtome and stained with hematoxylin and eosin (H&E). Histopathological evaluation of renal tissue and pancreatic islets was performed using a light microscope (Olympus CX33) at 400× magnification by two blinded observers using the same scoring system.
Renal injury was assessed using a semi-quantitative scoring system evaluating glomerular and tubular damage, interstitial inflammatory infiltration, and structural disorganization. Each parameter was graded on a five-point scale (0 - 4) according to the percentage of tissue involvement: 0 = normal; 1 = < 10%; 2 = 10 - 25%; 3 = 26 - 50%; and 4 = > 50%. Tubular necrosis was defined by cytoplasmic swelling, vacuolization, pyknotic or karyorrhectic nuclei, and luminal accumulation of cellular debris. Interstitial inflammatory infiltration was characterized by the presence of inflammatory cells, predominantly lymphocytes, within the interstitial space. Glomerular sclerosis was identified by capillary lumen occlusion, mesangial matrix expansion, glomerular shrinkage, or hyalinization. The total renal injury score was obtained by summing the individual parameter scores [23].
Pancreatic histomorphometric analysis was conducted following the method of Djabir et al. (2021) with slight modifications [24]. The total number of pancreatic islets (N) was determined at 100× magnification for each rat. The diameter of each islet was measured three times across its center along the vertical, horizontal, and diagonal axes. Five islets per rat, including both the largest and smallest, were selected to calculate the mean islet diameter at 40× magnification. The islet area was then estimated using the equation A = π(D/2)².
Statistical analysis
The numerical data were first tested for normality using the Shapiro Wilk test in GraphPad Prism 10.5.0. Values that were normally distributed were presented as mean ± standard deviation and analyzed using one-way ANOVA. When significant differences among groups were detected, Tukey’s test was applied to identify which group pairs differed significantly.
Results
The extraction yield obtained in this study was 16.75%. Qualitative phytochemical screening of the ethanolic extract of H. scabra demonstrated the presence of flavonoids, saponins, and steroids, while tannins, cardiac glycosides, and alkaloids were absent (Table 1).
Table 1 Phytochemical screening of ethanolic extract of Holothuria scabra.
Phytochemical Group |
Target compound |
Method |
Color for positive test |
Result |
Flavonoids |
Pyrone ring |
Shinoda test |
Yellow, red color |
() |
Tannins |
Polyphenolic |
Ferric chloride test |
Greenish black |
( ) |
Saponins |
Saponins |
Foam test |
Stable foam persists for 15 min |
() |
Triterpenoids & Steroids |
Triterpenoid/Steroid nucleus |
Lieberman-Burchard’s test |
- A red-brown ring forms at the interface - Upper layer: Dark yellow-brown (Triterpene), or green/moss-green (Steroid) |
() (Steroid) |
Carr-Price test |
- Blue fluorescence: Triterpene - Yellow fluorescence: Steroid |
() (Steroid) |
||
Steroid aglycone |
Lieberman-Burchard’s test
|
- A reddish-purple ring appears at the liquid interface - Lower layer: Pink - Upper layer: Green |
( ) |
|
Cardiac Glycoside |
5-membered lactone ring |
Baljet test |
Red or pink color |
( ) |
Legal’s test |
Orange or reddish orange color |
( ) |
||
2-deoxy sugar
|
Kellar-Kiliani test |
- A reddish-brown ring forms at the interface - Upper layer turns bluish-green |
( ) |
|
Alkaloids |
|
Borchardt reagent |
Brown precipitate |
( ) |
Dragendorff reagent |
Red-orange precipitate |
|||
Valse-Mayer reagent |
Yellowish-white precipitate |
|||
Bertrand reagent |
White precipitate |
As shown in Figure 1, body weight did not differ significantly among groups at baseline (day 0). Following streptozotocin (STZ) induction, body weight changes remained statistically comparable across groups through day 14. By day 29, the untreated diabetic group exhibited a significant reduction in body weight compared with the normal control group (p = 0.0017). In contrast, rats treated with H. scabra ethanolic extract or metformin showed less pronounced weight loss; however, these differences did not reach statistical significance when compared with the diabetic control group.
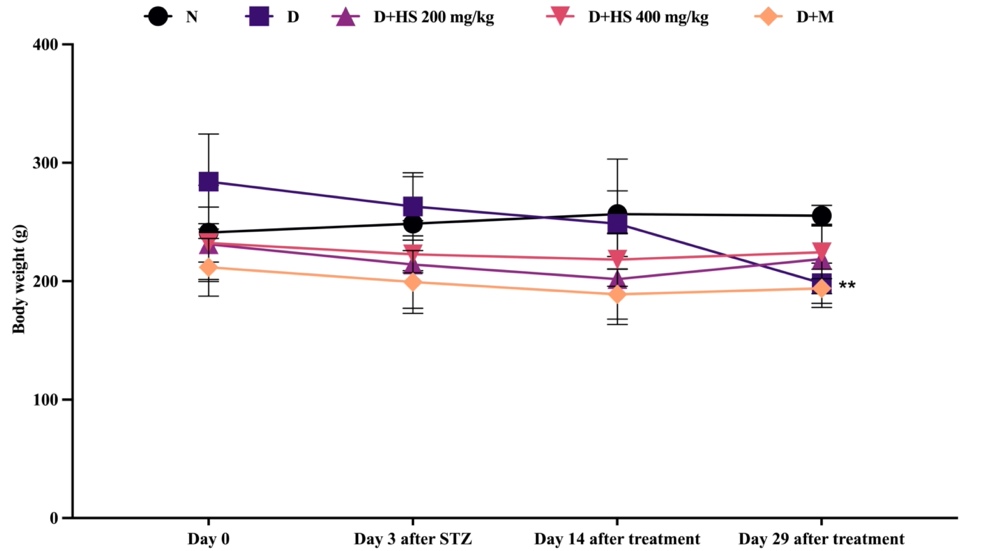
Figure 1 Changes in body weight (g) in normal (N), diabetic (D), ethanolic extract of H. scabra at a dose of 200 mg/kg (D+HS 200) and 400 mg/kg (D+HS 400), and metformin-treated (D+M) groups. Data are presented as mean ± SD. **p < 0.01 vs normal group.
Fasting blood glucose (FBG) levels (Figure 2) did not differ significantly among groups on day 0. After STZ induction, all diabetic groups demonstrated a marked increase in FBG compared with the normal group (p < 0.0001), which persisted until day 14, indicating sustained hyperglycemia in the diabetic groups, whereas the normal group maintained low FBG levels. No significant differences were observed between the treated groups and the diabetic control at this time point. On day 29, the diabetic control exhibited the highest FBG levels, significantly higher than those of the normal group (p < 0.0001). Administration of the ethanolic extract of H. scabra at 200 mg/kg significantly reduced FBG compared with the diabetic control (p = 0.0008); similarly, the 400 mg/kg dose (p = 0.0104) and metformin treatment (p = 0.0112) also resulted in significant reductions.
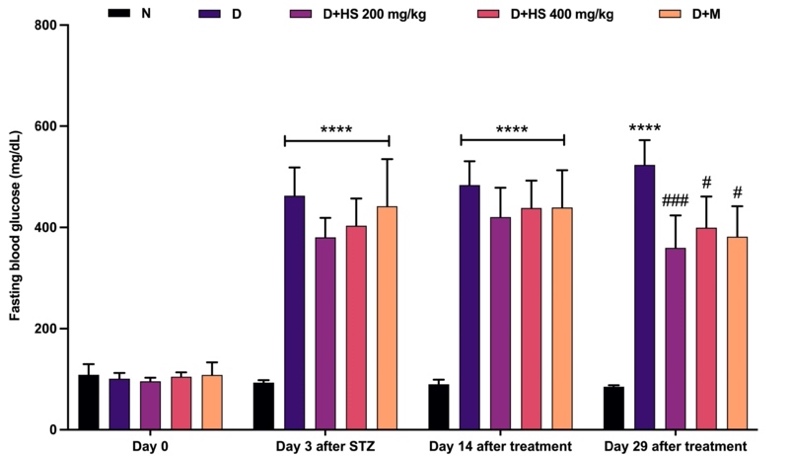
Figure 2 Changes in fasting blood glucose levels (mg/dL) in the normal (N), diabetic (D), H. scabra extract-treated (200 mg/kg [D+HS 200] and 400 mg/kg [D+HS 400]), and metformin-treated (D+M) groups. Data are presented as mean ± SD. ****p < 0.0001 vs normal group; #p < 0.05, ###p < 0.001, and ####p < 0.0001 vs diabetic group.
Figure 3 shows that renal biochemical parameters were similar among all groups at baseline (day 0), with no significant differences in serum creatinine or urea levels. At the end of the study, the diabetic group demonstrated significant renal impairment, indicated by elevated creatinine (p = 0.0193) and urea (p < 0.0001) levels compared with the normal group. Treatment with the ethanolic extract of H. scabra (200 mg/kg) significantly decreased creatinine levels compared with the diabetic group (p = 0.0427). On the other hand, metformin administration significantly reduced urea levels relative to the diabetic group (p = 0.0007).
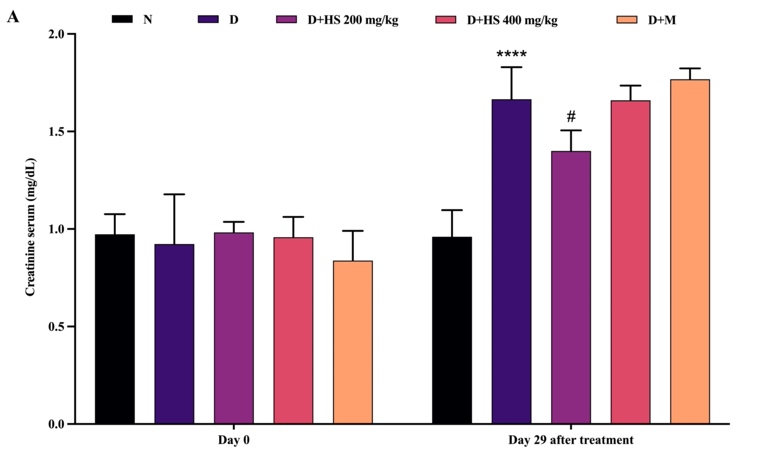
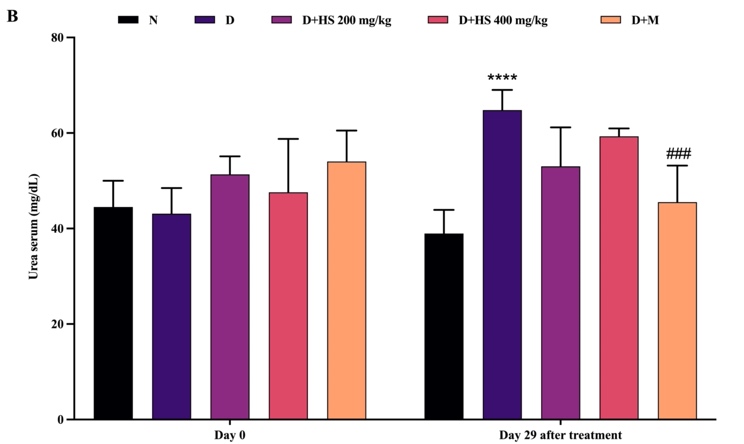
Figure 3 Serum creatinine (mg/dL) (A) and urea (mg/dL) (B) levels in the normal (N), diabetic (D), H. scabra extract-treated (200 mg/kg [D+HS 200] and 400 mg/kg [D+HS 400]), and metformin-treated (D+M) groups. Data are presented as mean ± SD. ****p < 0.0001 vs normal group; #p < 0.05 and ###p < 0.001 vs diabetic group.
Figures 4 and 5 depict rat kidney histopathology and the corresponding kidney damage parameters, respectively. Figure 4(B) demonstrates marked morphological alterations in the diabetic group, including dilation of Bowman’s space, glomerular deformity, tubular damage, and inflammatory cell infiltration. These findings were consistent with a significant increase in kidney damage scores (p < 0.001) and kidney index (p < 0.05) compared with the normal group (Figure 5). Administration of the ethanolic extract of H. scabra at 200 mg/kg (Figure 4(C)) and metformin (Figure 4(E)) resulted in histological improvement, characterized by more regular glomerular architecture, narrowing of Bowman’s space, tubular regeneration, and reduced inflammatory cell infiltration. At 400 mg/kg (Figure 4(D)), the glomeruli appeared relatively normal; however, tubular repair was less optimal than that observed at 200 mg/kg.
Treatment with the extract at 200 mg/kg (p = 0.0002) and 400 mg/kg (p = 0.0117), as well as metformin (p = 0.0034), significantly reduced kidney damage scores compared with the diabetic group. Nevertheless, the kidney index did not fully return to normal levels in any treatment group, as no significant difference was observed compared with the diabetic group (p > 0.05).
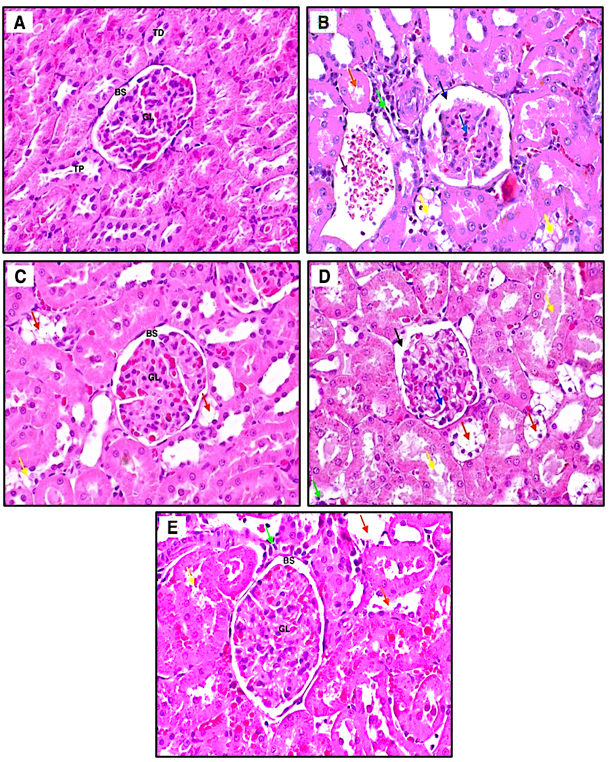
Figure 4 Representative photomicrographs of rat kidney histopathology in the normal control (A), diabetic control (B), H. scabra extract 200 mg/kg (C), H. scabra extract 400 mg/kg (D), and metformin-treated (E) groups. Arrows indicate distal tubular damage (orange), inflammatory infiltration (green), intratubular hemorrhage (purple), proximal tubular degeneration (yellow), glomerular alterations (blue), and widened Bowman’s space (black). Original magnification ×400.
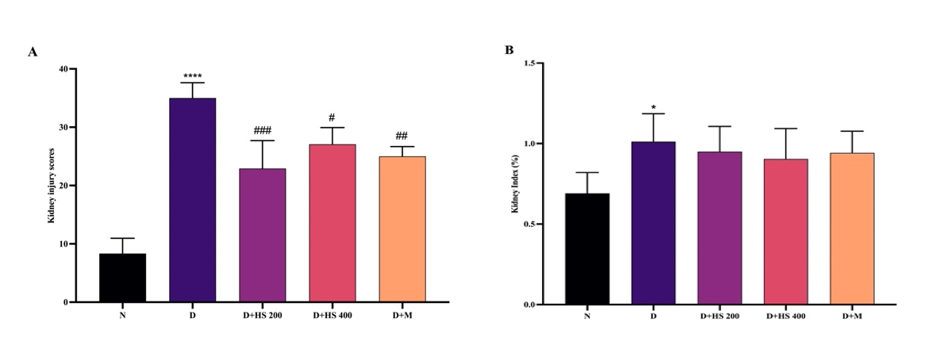
Figure 5 Kidney injury scores (A) and kidney index (B) in the normal (N), diabetic (D), H. scabra extract-treated (200 mg/kg [D+HS 200] and 400 mg/kg [D+HS 400]), and metformin-treated (D+M) groups. Data are presented as mean ± SD. *p < 0.05 and ****p < 0.0001 vs normal group; #p < 0.05, ##p < 0.01, ###p < 0.001 vs diabetic group.
For pancreatic histology, the normal group (Figure 6(A)) exhibited well-defined, large islets of Langerhans with clear boundaries and densely packed β-cells, consistent with intact pancreatic architecture. In contrast, the diabetic group (Figure 6(B)) demonstrated marked reduction in islet size with indistinct margins, accompanied by cytoplasmic degeneration, nuclear fading, acinar cell degeneration, and prominent vacuolization. Administration of H. scabra ethanolic extract at 200 mg/kg (Figure 6(C)) resulted in histological improvement to some degree, characterized by relatively larger islets, increased cellular density, and more distinct islet borders, although mild vacuolization persisted. Similarly, the metformin-treated group (Figure 6(E)) showed improved islet architecture with clearer boundaries and reduced cellular degeneration, despite the presence of mild interstitial edema. Quantitative analysis revealed significant differences between the normal and diabetic groups in islet number (p = 0.0003) and pancreatic index (p < 0.001). However, no significant differences were observed in islet number, islet area, or pancreatic index between the extract-treated or metformin-treated groups and the diabetic group (p > 0.05).
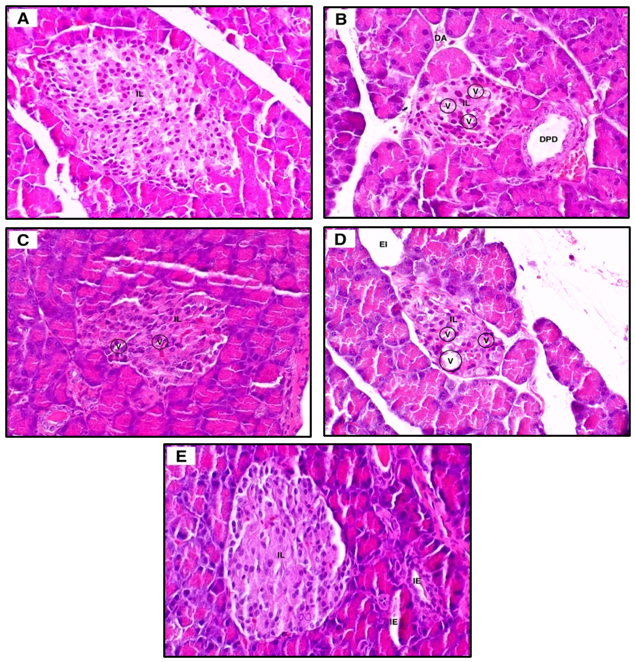
Figure 6 Representative pancreatic histology of normal (A), diabetic (B), Holothuria scabra extract-treated (200 mg/kg, C; 400 mg/kg, D), and metformin-treated (E) rats. IL, islet of Langerhans; DA, acinar degeneration; V, vacuoles; DPD, dilated pancreatic duct; EI, expanded interstitium; IE, interstitial edema. Original magnification ×100.
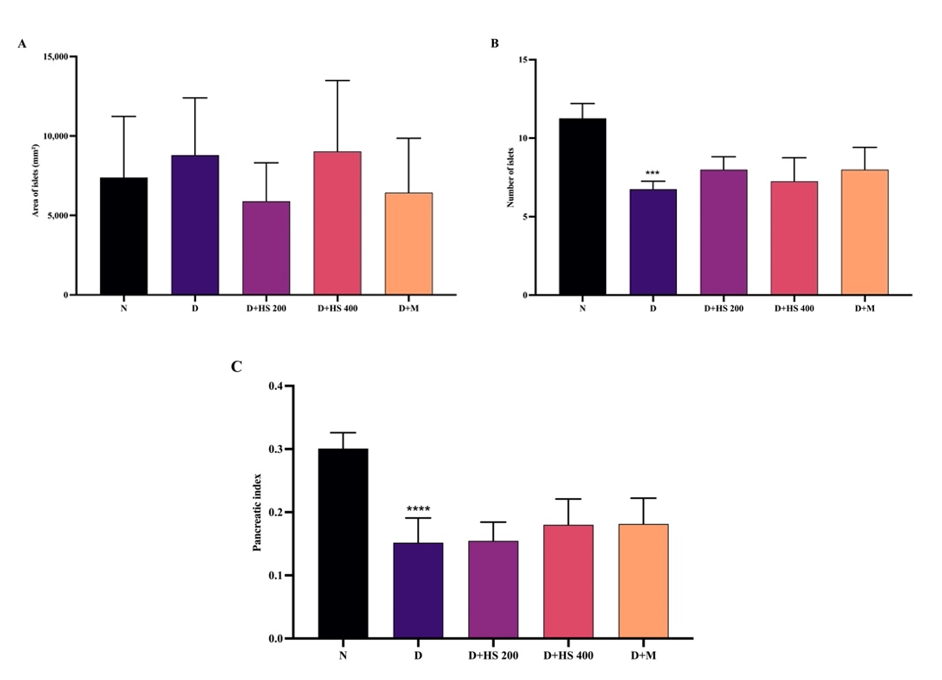
Figure 7 Pancreatic parameters: (A) islet number, (B) islet area, and (C) pancreatic index in normal (N), diabetic (D), H. scabra extract-treated (200 and 400 mg/kg), and metformin-treated groups. Data are mean ± SD. ***p < 0.001 and ****p < 0.0001 vs N.
Discussion
STZ induces diabetes by targeting pancreatic β-cells through its high affinity for the GLUT2 transporter, allowing the compound to readily enter the cells. Inside β-cells, STZ is cleaved into a glucose moiety and methyl nitrosourea; the alkylating methyl nitrosourea then modifies biological macromolecules, damages DNA, and triggers β-cell death. This process is further amplified by DNA methylation, increased production of nitric oxide and reactive oxygen species, and depletion of NAD⁺, which is essential for cellular energy metabolism. Damage also occurs to mitochondrial DNA, disrupting metabolic signaling and inhibiting glucose-stimulated insulin secretion. The combination of these mechanisms leads to severe cellular stress, resulting in β-cell destruction and the development of insulin-dependent diabetes [25,26].
Diabetic rats typically experience progressive body weight loss as a consequence of sustained hyperglycemia and insulin deficiency. Impaired glucose utilization promotes a shift toward catabolic metabolism, characterized by increased lipolysis and proteolysis to meet energy demands [27]. Consistent with this mechanism, the untreated diabetic group in the present study exhibited significant body weight reduction. However, despite significant reductions in fasting blood glucose levels, administration of the ethanolic extract of H. scabra for 28 days did not fully restore body weight. This finding suggests that while the extract improved glycemic control, the duration of treatment may have been insufficient to reverse the established catabolic state or to normalize metabolic balance.
The antidiabetic effects observed in this study may be attributed to the bioactive constituents present in Holothuria species. Saponins and triterpenoid/steroidal compounds have been reported to improve glycemic control by enhancing insulin secretion and sensitivity, suppressing hepatic gluconeogenesis, and promoting peripheral glucose utilization. In addition, they also exert antioxidant and anti-inflammatory activities that help protecting pancreatic β-cells [28,29]. Phytochemical screening also identified flavonoids and tannins, which are known to inhibit intestinal glucose absorption, improve insulin signaling, and mitigate oxidative stress–induced β-cell damage [30,31].
Although significant reductions in fasting blood glucose were observed, quantitative pancreatic parameters, including islet number and area, were not significantly different from the diabetic control group. This discrepancy suggests that glycemic improvement may not depend solely on structural regeneration of β-cells. Instead, the glucose-lowering effect of H. scabra ethanolic extract may predominantly involve extra-pancreatic mechanisms, such as suppression of hepatic gluconeogenesis and modulation of insulin signaling pathways [32]. Thus, despite only partial histological recovery of pancreatic tissue, the extract appears to exert functional metabolic regulation that translates into systemic improvement.
Improved metabolic control may, in turn, contribute to attenuation of diabetes-induced renal injury. In the present study, extract-treated groups showed better preservation of glomerular and tubular architecture, accompanied by reduced injury scores and lower serum creatinine levels. Interestingly, the 200 mg/kg dose demonstrated more consistent histological improvement than the 400 mg/kg dose, particularly in tubular regeneration. This pattern suggests a possible non-linear dose–response relationship. In phytopharmacology, higher doses do not necessarily confer greater efficacy, as excessive concentrations of bioactive compounds may lead to reduced bioavailability, metabolic burden, or counter-regulatory effects [33]. Therefore, further dose-optimization studies are warranted to define the therapeutic window of the extract.
Limitations
This study has several limitations that should be acknowledged. First, the relatively small sample size (n = 5 per group), without a priori power calculation, may limit the statistical strength and generalizability of the findings. Second, the treatment duration was relatively short, which may not fully capture long-term metabolic and renal effects. Third, acute and sub-chronic toxicity evaluation was not performed and should be conducted in future studies to establish the safety profile and therapeutic window of the extract. Fourth, the absence of detailed cellular and molecular analyses restricts mechanistic insight into the pathways underlying the observed antidiabetic and renoprotective activities. Future studies incorporating comprehensive molecular investigations, oxidative stress markers, inflammatory mediators, and insulin signaling pathways, are warranted to better elucidate the mechanisms of action of the ethanolic extract of H. scabra.
Conclusions
This study demonstrates that the ethanolic extract of Holothuria scabra exerts integrated antidiabetic and renoprotective effects in streptozotocin-induced diabetic rats. Beyond lowering fasting blood glucose, the extract attenuated renal structural damage and reduced injury scores. The 200 mg/kg dose showing more consistent histological improvement, suggesting a potential non-linear dose–response relationship. Importantly, glycemic reduction occurred despite the absence of significant restoration of pancreatic islet number and area, indicating that the metabolic benefits may involve functional enhancement of residual β-cells and/or extra-pancreatic mechanisms rather than structural regeneration alone. The identification of flavonoids, saponins, and steroidal compounds provides a plausible phytochemical basis for the observed antioxidant and metabolic regulatory effects. These findings support H. scabra as a promising marine-derived candidate for adjunctive diabetes therapy and warrant further mechanistic and dose-optimization studies.
Acknowledgements
The authors would like to thank the staff of the Laboratory of Biopharmaceutics and Clinical Pharmacy, Faculty of Pharmacy, Hasanuddin University, for their technical assistance during the experimental procedures. This study was partially supported by funding obtained through Drug Efficacy and Medication Safety Thematic Research Group, Hasanuddin University (contract number:00518/UN4.22/PT.01.03/2025).
Declaration of Interest
The authors declare no known competing interests or personal relationships that could have appeared to influence the work reported in this paper.
Declaration of generative AI in scientific writing
The authors declare that AI tools were used solely to improve language clarity and manuscript presentation and did not influence the scientific content, data analysis, interpretation, or conclusions of the study. ChatGPT was utilized for language refinement and structural editing, and Grammarly was used for grammar and spelling correction.
Credit author statement
Ahmad Fajri: Writing Original draft, Investigation, Formal analysis, Visualization. Amiroh Rochimah Imam: Investigation, Data analysis. Hien Minh Nguyen: Conceptualization, Supervision, and Writing-Review & Editing. Huyn Ha Thy: Data curation. Yulia Yusrini Djabir: Conceptualization, Resources, Methodology, Supervision, and Writing-Review & Editing.
References
[1] AME Elkhalifa, M Nazar, SI Ali, I Khursheed, S Taifa, MA Mir, IH Shah, M Malik, Z Ramzan, S Ahad, N Bashir, E Elamin, EA Bazie, EM Ahmed, MM Alruwaili, AW Baltoyour, AS Alarfaj, IA Al Bataj, AMA Arabe and SU Nabi. Novel therapeutic agents for management of diabetes mellitus: A hope for drug designing against Diabetes Mellitus. Life 2024; 14(1), 99.
[2] H Sun, P Saeedi, S Karuranga, M Pinkepank, K Ogurtsova, BB Duncan, C Stein, A Basit, JCN Chan, JC Mbanya, ME Pavkov, A Ramachandaran, SH Wild, S James, WH Herman, P Zhang, C Bommer, S Kuo, EJ Boyko and DJ Magliano. IDF Diabetes Atlas: Global, regional and country-level diabetes prevalence estimates for 2021 and projections for 2045. Diabetes Research and Clinical Practice 2022; 183, 109119.
[3] Y Luo, W Zhang and G Qin. Metabolomics in diabetic nephropathy: Unveiling novel biomarkers for diagnosis (Review). Molecular Medicine Reports 2024; 30(3), 156.
[4] P Paul, L Chacko, TK Dua, P Chakraborty, U Paul, VV Phulchand, NK Jha, SK Jha, R Kandimalla and S Dewanjee. Nanomedicines for the management of diabetic nephropathy: Present progress and prospects. Frontiers in Endocrinology 2023; 14, 1236686.
[5] G Hou, Y Dong, Y Jiang, W Zhao, L Zhou, S Cao and W Li. Immune inflammation and metabolic interactions in the pathogenesis of diabetic nephropathy. Frontiers in Endocrinology 2025; 16, 1602594.
[6] Y Ratan, A Rajput, A Pareek, A Pareek and G Singh. Comprehending the role of metabolic and hemodynamic factors alongside different signaling pathways in the pathogenesis of diabetic nephropathy. International Journal of Molecular Sciences 2025; 26(7), 3330.
[7] J Jin and M Zhang. Exploring the role of NLRP3 inflammasome in diabetic nephropathy and the advancements in herbal therapeutics. Frontiers in Endocrinology 2024; 15, 1397301.
[8] MF Nisar, M Li J Xu and C Wan. Anti-diabetic effects of marine natural products through redox modulation via Nrf2/HO-1 cytoprotective pathways. Frontiers in Marine Science 2024; 11, 1438955.
[9] DAAS Laksemi, PAA Damayanti, IM Sudarmaja, IK Tunas, LPR Sundari, NL Rustini and PIB Apsari. In-vivo antimalarial activity of Holothuria scabra simplicia in Plasmodium berghei-infected mice. Universa Medicina 2024; 43, 195-201.
[10] S Xia, M Li, T Shen, Y Gao, X Ma and X Yuan. Chapter 5 - Sea cucumbers in Chinese daily life. In: A Mercier, JF Hamel, AD Suhrbier and CM Pearce (Eds.). The World of Sea Cucumbers. Elsevier, Amsterdam, 2024, p. 65-75.
[11] TL Wargasetia, H Ratnawati, N Widodo and MH Widyananda. Antioxidant and anti-inflammatory activity of sea cucumber (Holothuria Scabra) active compounds against KEAP1 and iNOS protein. Bioinformatics and Biology Insights 2023; 17, 1-10.
[12] JF Hamel, I Eeckhaut, C Conand, J Sun, G Caulier and A Mercier. Global knowledge on the commercial sea cucumber Holothuria scabra. Advances in Marine Biology 2022; 91, 1-286.
[13] OC Eleazu, KC Eleazu, S Chukwuma and UN Essien. Review of the mechanism of cell death resulting from streptozotocin challenge in experimental animals, its practical use and potential risk to humans. Journal of Diabetes and Metabolic Disorder 2013; 12, 60.
[14] A Ghasemi and S Jeddi. Streptozotocin as a tool for induction of rat models of diabetes: A practical guide. EXCLI Journal 2023; 22, 274-294.
[15] A Fiqardina, YY Djabir, A Santoso, SN Salsabil and I Ismail. The nephroprotective effect of clove oil (Oleum caryophylli) against levofloxacin toxicity in rats. Iranian Journal of Toxicology 2022; 16, 27-34.
[16] K Godlewska, P Pacyga, A Najda and I Michalak. Investigation of chemical constituents and antioxidant activity of biologically active plant-derived natural products. Molecules 2023; 28, 5572.
[17] L Maheshwaran, L Nadarajah, SPNN Senadeera, CB Ranaweera, AK Chandana and RN Pathirana. Phytochemical testing methodologies and principles for preliminary screening/ qualitative testing. Asian Plant Research Journal 2024; 12(5), 11-38.
[18] A Alqethami and AY Aldhebiani. Medicinal plants used in Jeddah, Saudi Arabia: Phytochemical screening. Saudi Journal Biological Sciences 2021; 28(1), 805-812.
[19] L Traore, YA Bekro, JL Pirat, and JA Mamyrbekova. Study of crude extracts from Cassia sieberiana root bark and Khaya grandifoliola trunk bark: Phytochemical screening, quantitative analysis and radical scavenging activity. International Journal of Current Pharmaceutical Research 2015; 7(3), 22-26.
[20] MB Ekong, FN Odinukaeze, AC Nwonu, CC Mbadugha, and AA Nwakanma. Brain activities of streptozotocin-induced diabetic wistar rats treated with gliclazide: Behavioural, biochemical and histomorphology studies. IBRO Neuroscience Reports 2022; 12, 271-279.
[21] D Cheng, B Liang and Y Li. Antihyperglycemic effect of Ginkgo biloba extract in streptozotocin-induced diabetes in rats. Biomed Research Internasional 2013; 2013(7), 162724.
[22] AD Anugrah, A Asmara, Y Leman and YY Djabir. Sandfish (Holothuria scabra) ethanolic extract mitigates ethylene glycol-induced acute kidney injury in rats. Natural Resources for Human Health 2025; 5, 300-309.
[23] S Usman, RI Juhardi, SA Iraekawati, NY Linthin, A Arsyad and YY Djabir. Protective effects of astaxanthin on triple whammy-induced acute kidney injury via antioxidant, anti-inflammatory and anti-apoptotic mechanisms. Trends in Sciences 2026; 23(2),11484.
[24] YY Djabir; H Hardi, H Setiawati, S Lallo, Risfah Yulianty, MH Cangara and H Veni. Artocarpus altilis leaf extract protects pancreatic islets and improves glycemic control in alloxan-induced diabetic rats. Journal of Reports in Pharmaceutical Sciences 2021; 10(1), 87-92.
[25] AKS Brito, AVS Mendes, BT Acha, ASSS Oliveira, JL Macedo, AS Cruzio, MDG Prianti, RR Abreu, M Lucarini, A Durazzo, MDCC Martins and DDR Arcanjo. Experimental models of type 2 diabetes mellitus induced by combining hyperlipidemic diet (HFD) and streptozotocin administration in rats: An integrative review. Biomedicines 2025; 13(5), 1158.
[26] S Lenzen. The mechanisms of alloxan- and streptozotocin-induced diabetes. Diabetologia 2008; 51(2), 216-226
[27] W Fang, X Yuan, W Li, S Seery, G Chen, Z Cai, Z Huang, X Wang, W Wu, Z Chen, Y Li, S Wu and Y Chen. Excessive weight gain onset-age and risk of developing diabetes mellitus: A large, prospective Chinese cohort study. Frontier in Endocrinology 2023; 14, 1281203.
[28] A El Barky, SA Hussein, AAE Alm-Eldeen, A Hafez and TM Mohamed. Diabetes management saponins and their potential role in diabetes mellitus. Diabetes Management 2017; 7(1), 148-158.
[29] YE Puspitasari, E Tuenter, K Foubert, H Herawati, A Hariati, A Aulanni’am, L Pieters, TD Bruyne and N Hermans. Saponin and fatty acid profiling of the sea cucumber Holothuria atra, α-Glucosidase inhibitory activity and the identification of a novel triterpene glycoside. Nutrients 2023; 15, 1-20.
[30] A Hossain, D Dave and F Shahidi. Antioxidant Potential of sea cucumbers and their beneficial effects on human health. Marine Drugs 2022; 20, 521.
[31] RF Sinuhaji, S Ramadhani, VK Setiawan and U Baroroh. Targeting diabetes with flavonoids from Indonesian medicinal plants: A review on mechanisms and drug discovery. Naunyn Schmiedebergs Arch Pharmacol 2025; 398, 11683-11703.
[32] M Li, X Chi, Y Wang, S Setrerrahmane, W Xie and H Xu. Trends in insulin resistance: Insights into mechanisms and therapeutic strategy. Signal Transduction and Targeted Therapy 2022; 7, 216.
[33] L Yang, J Wang, RA Cheke and S Tang. A Universal delayed difference model fitting dose-response curves. Dose Response 2021; 19(4), 1-18.