Trends
Sci.
2026;
23(10):
13692
Histopathology of the Coffee Berry Borer Hypothenemus hampei (Ferrari) Infected by the Entomopathogenic Fungi Beauveria bassiana
Sinlapachai Senarat1,2, Narit Thaochan2,3,*, Kodeeyah Thoawan3,
Peerasak Bunsap3, Kitipong Angsujinda4, Pisit Poolprasert5,
Anjaree Inchan6, Natthawut Charoenphon7,
Supapong Imsonpang8 and Gen Kaneko9
1Division of Biological Sciences, Faculty of Science, Prince of Songkla University, Songkhla 90110, Thailand
2Center for Integrative Animal Tissue Biology and Innovation (iTissue-PSU), Prince of Songkla University,
Songkhla 90110, Thailand
3Agricultural Innovation and Management Division (Pest Management), Faculty of Natural Resources, Prince of Songkla University, Songkhla 90110, Thailand
4Aquatic Resources Research Institute, Chulalongkorn University, Bangkok 10330, Thailand
5Department of Entomology, Faculty of Agriculture, Kasetsart University, Bangkok 10900, Thailand
6Faculty of Medicine, Praboromarajchanok Institute, Ministry of Public Health, Nonthaburi 11000, Thailand
7Department of Anatomy, Faculty of Medical Science, Naresuan University, Phitsanulok 65000, Thailand
8Division of Health and Applied Science, Faculty of Science, Prince of Songkla University, Songkhla 90110, Thailand
9College of Natural and Applied Science, University of Houston-Victoria, Victoria, Texas 77901, USA
(*Corresponding author’s e-mail: [email protected])
Received: 25 February 2026, Revised: 29 March 2026, Accepted: 10 April 2026, Published: 30 April 2026
Abstract
The coffee berry borer (Hypothenemus hampei) is one of the most destructive pests of coffee worldwide, causing severe economic losses. The entomopathogenic fungus Beauveria bassiana has been widely used as a biological control agent; however, detailed histopathological responses of H. hampei to fungal infection remain poorly understood. This study investigated the morpho-histopathological alterations and apoptosis in adult H. hampei following exposure to B. bassiana. Beetles were immersed in a spore suspension (1×10⁹ conidia/mL) and sampled at multiple time points up to 144 h post-exposure. Histological analyses were performed using hematoxylin-eosin, PAS, Masson’s trichrome, and Grocott’s methenamine silver staining. Apoptotic cells were detected using TUNEL assay, and tissue damage was evaluated using the histological alteration index (HAI). Fungal hyphae were detected in the integument and adipose tissue as early as 12 h post-exposure. From 24 to 144 h, progressive fungal proliferation, cuticle thinning, muscle degeneration, adipose tissue degeneration, and gut epithelial erosion were observed. Quantitative analysis revealed a significant increase in fungal area and a marked reduction in cuticle thickness (p < 0.001). Apoptotic cell density increased in a time-dependent manner, with strong immunoreactivity observed after 60 h. These findings demonstrate that B. bassiana rapidly penetrates and disrupts host tissues, leading to extensive structural damage and apoptosis. Understanding the infection dynamics of B. bassiana provides important insights for optimizing biological control strategies and enhancing sustainable pest management in coffee agroecosystems.
Keywords: Beauveria bassiana, Coffee berry borer, Entomopathogenic fungi, Histopathology, Apoptosis, Biological control, Cuticle degradation, Thailand
Introduction
The coffee berry borer Hypothenemus hampei, a member of the Curculionidae family, is a tiny insect of great concern to the coffee industry. It threatens coffee cultivation worldwide on large-scale commercial plantations as well as smaller family-owned plantations [1]. This beetle of 1.4 - 1.7 mm infests coffee cherries at all stages of maturity. The infestation cycle begins with a female insect drilling into the coffee cherry, introducing fungi to establish a ‘fungus farm’ within the fruit. The female then lays 30 to 70 eggs inside the cherry, developing into adult beetles through larval and pupal stages. These symbiotic microorganisms serve as a food source for the developing larvae. Their proliferation, together with the feeding activity of the beetle, contributes to the degradation of coffee berry tissues and may facilitate secondary fungal infections. Infestation by the coffee berry borer impairs coffee quality and increases premature bean drop, particularly in Arabica and Robusta varieties [1]. Barrera [2] noted that coffee bean drops increase 30% - 35% after borer infestation and highlighted the significant impacts of the pest. The economic impacts include reduced coffee prices and reduced conformity to international trade standards, which control coffee exports with stringent regulations. Even a 1.5% infestation trace on the bean surface is potentially a barrier to global markets [3].
Due to the severe repercussions of coffee borer infestation, various control measures have been implemented. Buranapanitchaphan and Chanbang [4] introduced pheromone traps to attract and capture the pest, showing promising results in reducing populations of coffee berry borers. Still, biological control methods that utilize infectious fungi have become increasingly favored. Several entomopathogenic fungi, including Beauveria bassiana, have effectively managed coffee berry borers and other pests [5-7]. More recently, De La Rosa et al. [8] applied B. bassiana to control coffee berry borer infection, finding a spore density of 1×109 conidia/mL the most effective control. Another fungus species, Metarhizium anisopliae, has been formulated into a ready-to-use powder that was highly effective against the coconut rhinoceros beetle (Oryctes rhinoceros) [9].
Histopathology studies cellular and tissue changes that indicate animal health status and physiological responses [10-12]. Histopathological studies can elucidate the effects of fungal pathogens on the coleopterans. Bava et al. [11] initially observed degenerative changes in the Malpighian tubules of the corn planthopper, Peregrinus maidis, following exposure to B. bassiana and M. anisopliae. Duan et al. [12] then found that B. bassiana could penetrate the midgut of the colorado potato beetle, Leptinotarsa decemlineata, within 24 h, causing the degeneration and eventual death of integument. Ibrahim et al. [13] also noted histopathological alterations in the skin and hemocytes of Zeuzera pyrina larvae after a 3 - 4-day exposure to B. bassiana. However, there is still limited information about the histological dynamics of fungal invasion and host response, including the case of B. bassiana and the coffee berry borer.
The objective of this study was to investigate the histopathological alterations in the coffee berry borer following exposure to B. bassiana to elucidate fungal invasion and host physiological responses. The findings, including rapid penetration, progressive tissue damage, and increased apoptosis, provide a basis for optimizing application timing and improving formulation strategies, thereby supporting the development of more effective ready-to-use biocontrol products.
Materials and methods
Sampling of coffee berry borer
Coffee cherries infested by the coffee berry borer were collected in the dry season (March to April 2023) from 2 sites in Thailand: Thepha Research Station at the Faculty of Natural Resources in Thepha District, Songkhla Province, and the Agricultural Career Promotion and Development Center in Mueang Yala District, Yala Province. Infested cherries were randomly selected in the field to avoid sampling bias and then transported in plastic containers to the laboratory at the Department of Agricultural Innovation and Management, Prince of Songkla University, Hat Yai. The coffee berry borers were removed from the sampled coffee cherries and placed in 5 cm Petri dishes for species identification following the taxonomic guidelines of Smith et al. [14]. A Leica S8 APO stereo microscope was used for observation and photography.
Fungus cultivation
The B. bassiana fungal strain PSUB01 was kept according to a specialized preservation method from our previous publication [15] at the Laboratory of Pest Management, Faculty of Natural Resources, Prince of Songkhla University. For cultivation, the fungus was transferred onto SDAY agar plates and incubated for 14 days at 28.0 ± 0.2 °C without light. The duration of incubation was selected to ensure the complete development of fungal spores on the surface of the agar (Figures 1(A) and 1(B)). After this period, the spores were collected through filtration, preparing a spore suspension in distilled water with a concentration of 1×109 spores/mL for further experiments. This concentration is a high-density suspension and highly achieved effective pest control [15].
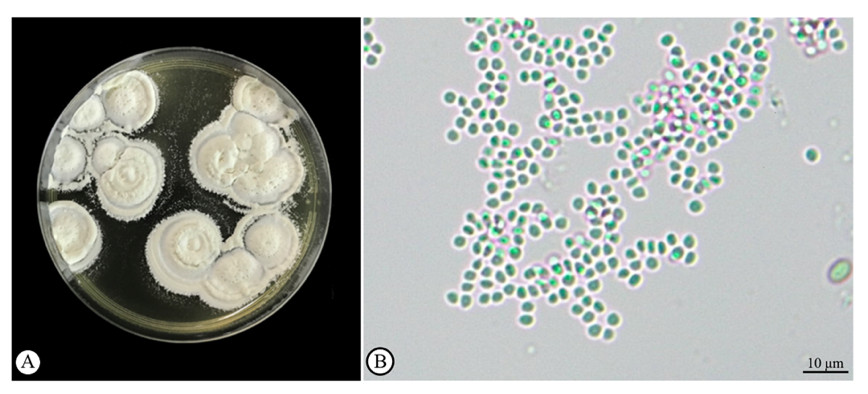
Figure 1 Beauveria bassiana spores on the surface of agar (A) and high magnification showing its spore (arrow) under a light microscopic observation (B).
Preparation of B. bassiana powdered formulation
Partially cooked rice was used as the cultivation substrate for B. bassiana. A heat-sealable bag measuring 7 11 inches was filled with 200 g of cooked rice. Beuveria bassiana spores were sprayed onto the rice, the bag was vigorously shaken to ensure the spores were uniformly distributed on the rice, and a rubber band was used to seal the bag. To facilitate air exchange, 20 holes were made in the bag with a pin. Beuveria bassiana was incubated for 14 days at 28.0 ± 0.2 °C under natural light conditions. After incubation, the spores were collected from the rice using a sieve. The spore powder was then dried in a desiccator for 5 days at 28.0 ± 0.2 °C. The dry spore powder was thoroughly mixed with a mixture of talcum and kaolin powders at a 1:9 ratio. The homogeneous powder was subsequently stored in zip-lock foil bags until used.
Infection of H. hampei with B. bassiana
To minimize contamination from handling, a small brush was always used to transfer H. hampei. For the control group, 30 beetles were submerged in sterilized distilled water for 1 min (N = 10, triplicate), whereas the experimental group was submerged in the spore suspension (1×109 spores/mL) for 1 min. After submersion, the insects were transferred and placed in the plastic containers and maintained under laboratory conditions (28 ± 1 °C, 80% - 90% RH) until sampling. H.hampei (N = 45) were sampled from the experimental group at 0, 6, 12, 18, 24, 30, 36, 48, 54, 60, 72, 84, 96, 122, and 144 h after submersion in the spore suspension. For each time point, randomly chosen specimens (N = 3) were dissected into sections along a horizontal axis. Three replications were performed throughout the study. Both control and experimental groups were preserved in Davidson’s fixative solution for about 24 h to prepare them for structural examination. The tissues of other beetles were fixed in 100 µL of fixative solution in a 1.5 mL microcentrifuge tube for 36 h to preserve tissue structure and prevent degradation. The samples were then transferred to 70% ethanol and stored at 4 °C until further histological analysis.
Histological and histopathological examination of beetle specimens
The fixed H. hampei samples were prepared for histological observation following Suvarna et al. [16], producing three tissue sections approximately 4 μm in thickness, which were stained with hematoxylin and eosin to reveal the structural and histopathological alterations within the tissues. The extent of tissue alterations was evaluated using the Histological Alteration Index (HAI) [17]. Three levels of lesion were classified including 0 = no change, 1 = mild, 2 = moderate, and 3 = severe. The histological alteration was evaluated and calculated by multiplying the severity grade by the percentage of the affected area (0% - 100%). The resulting scores ranged from 0 to 300.
Various additional staining methods were employed to enhance visualization of tissue structures and fungal elements. Masson’s Trichrome was used to detect the muscle and connective tissues, Periodic Acid-Schiff (PAS) staining was applied to detect polysaccharides and fungal cell wall components, and Grocott’s Methenamine Silver (GMS) staining was used to specifically visualize fungal hyphae [15,18].
The stained sections were observed using a conventional light microscope, and high-definition images were captured with a 3D Histech Pannoramic SCAN 150 scanner (Budapest, Hungary). Six distinct layers of the cuticle were measured for thickness, and the area of fungus coverage was determined. The ImageJ (Version 18.0) software was used for quantitative measurements.
Cell death analysis by TUNEL staining
Following the method of Rojo and Gonzalez [19], the terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assay was subjected using in situ TUNEL detection kit (Abcam, UK., Cat No: ab206386) in accordance with the manufacturer’s instructions to identify apoptotic cell death in unstained H. hampei specimens. Consecutive sections from previous topic were deparaffinized, rehydrated, and kept in the permeabilization in a solution containing dilated proteinase K. The sections were then treated with 30% hydrogen peroxide in methanol and subsequently incubated in TdT buffer at 37 °C for 1 h. TUNEL staining was subsequently carried out. A standard light microscope was used to observe the labeled sections, and cell death maps were created from images to show the spatial distribution of apoptotic cells within tissues. The number of apoptotic cells in 10 specimens was determined from 3 permanent slides of each specimen (N = 30) at a 40× magnification. The ImageJ software was employed to ensure accurate cell counting and to measure areas of interest. Additionally, the density of apoptotic cells in the tissues was evaluated by a semiquantitative method, assigning values based on the level of TUNEL-positive signal. No TUNEL-positive signal was represented by the symbol ‘-’, weak TUNEL-positive signal by ‘+’, moderate TUNEL-positive signal by ‘++’, and strong TUNEL-positive signal by ‘+++’.
Statistical evaluation of experimental data
The normality of all data was checked using the Shapiro-Wilk normality test. One-way analysis of variance (ANOVA) followed by Dunnett’s post-hoc test to compare the control group with the experimental groups, using IBM SPSS 22.0 and the R version 4.3.1 multcomp package 1.4 - 25. The significant threshold level was set at 95% (p < 0.05).
Results and discussion
Histopathology of Hypothenemus hampei infected by Beauveria bassiana
Before fungal infection, the external morphology of H. hampei was normal (Figure 2(A)), and no fungal population was observed in the structure of the integument (Figures 2(B) and 2(C)). Twelve h after exposure, no fungi were observed externally (Figure 2(D)). Still, the development of B. bassiana hyphae was observed in almost all adipose tissues (Figure 2(E)). In the outer integument (Figure 2(F)). The presence of hyphae dramatically increased in the host tissue from 24 to 120 h post-exposure (Figures 2(G) - 2(R)). Significant cellular alterations were observed in the integument system and adipose tissue with prominent hyphae (Figures 2(P) - 2(R)).

Figure 2 Light microscope images of the representative abdomen position in the adult Hypothenemus hampei infected with Beauveria bassiana - (A): Adult Hypothenemus hampei without infection; (B): Normal integument structure after H and E staining; (C): Normal integument structure after Grocott’s methenamine silver stain (GMS); (D-F): At 12 h post-exposure, hyphae had started to invade the adipose tissue beneath the cuticle (E, H&E; F, GMS); (G-I): At 24 h post-exposure, hyphae were spread out in the adipose tissue (H, H&E; I, GMS); (J-L): At 48 h post-exposure, further expansion of hyphae was observed (K, H&E; L, GMS); (M): At 84 h post-exposure, prominent hyphae were observed in the external morphology (N, H&E; O, GMS); (P-R): At 120 h post-exposure (Q, H&E; R GMS). Abbreviations: Bb = Beauveria bassiana, H&E = hematoxylin and eosin, GMS = Grocott’s methenamine silver stain, Ig = integument, Th = thorax, Ad = adipose tissue, Ms = muscle, Ep = epithelium.
We also prepared longitudinal sections to observe B. bassiana throughout the length of H. hampei. Several fungal features, including hyphae, conidiophores and conidia, were widely distributed, and hyphal penetration of the integument was also observed (Figures 3(A) - 3(C)). In parallel to a specialized infection structure of appressorium was successfully observed, which it was produced the penetration peg together with the penetration hyphae (Figures 3(D) - 3(F)).
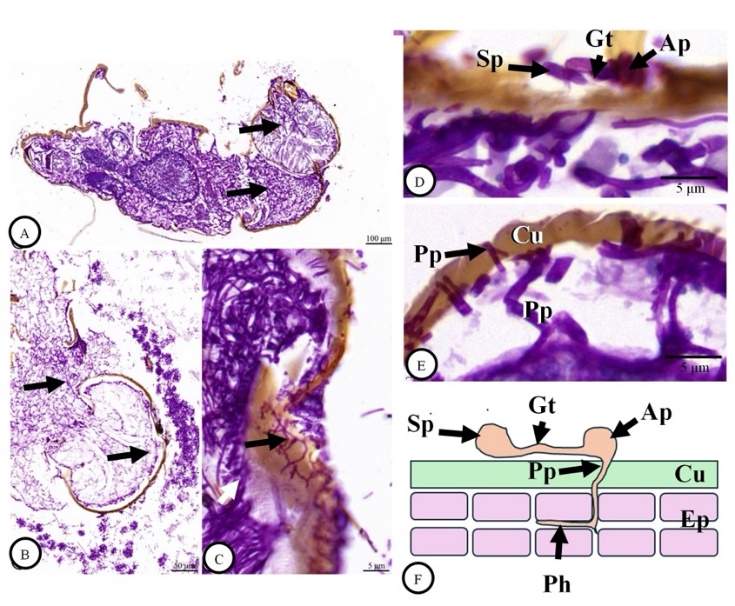
Figure 3 Light microscope images of a longitudinal section of an adult Hypothenemus hampei infected with Beauveria bassiana at 120 h post-exposure. The positive reaction of fungal features (arrows) with Periodic Acid-Schiff (PAS) reagent was observed throughout the body (A-B). High magnification revealed the development of fungal hyphae via the integument (black arrows) and the body area (white arrow). PAS staining figures and illustrated figure revealed that a specialized infection structure of appressorium (Ap) produced the penetration peg (Pp) together with the penetration hyphae (Ph) (D-F). Abbreviations: Cu = cuticle, Ep = epidermis, Gt = germinal tube, Ph = Penetration of hyphae.
Fungal area and cuticle thickness of Hypothenemus hampei
Grocott’s methenamine silver stain (GMS) clearly distinguished the area of B. bassiana within tissues (Figure 2), allowing us to quantify the extent of fungus growth within the body cavity. A substantial increase in the area of the fungus was observed from 48 to 144 h post-exposure (Figure 4(A)). In contrast, the thickness of the cuticle showed a significant reduction (p < 0.001) from 12 to 144 h post-exposure (Figure 4((B)). Cuticle thickness was significantly reduced at 12 h post-exposure but decreased more slowly at subsequent time points.
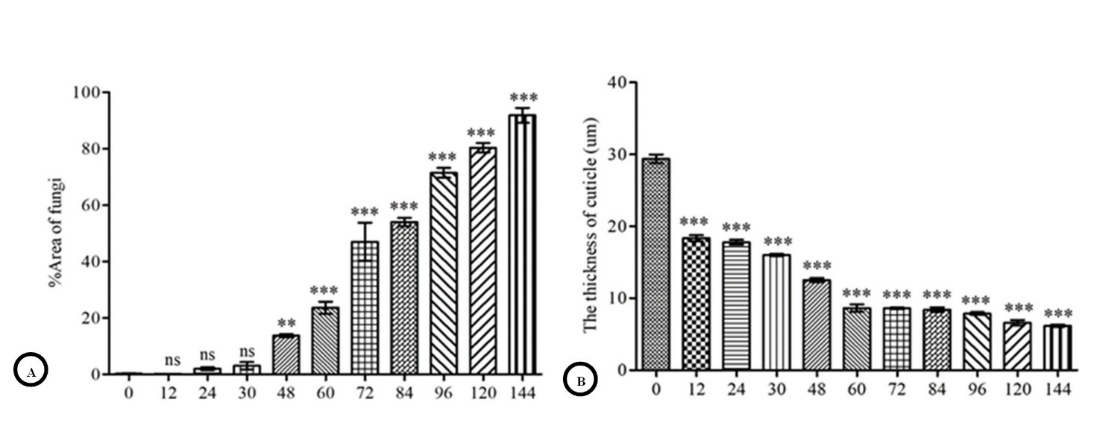
Figure 4 Quantitative evaluation of Beauveria bassiana infection. Values are represented as means ± SE. The significant difference was determined by Dunnett’s test (**p < 0.01, ***p < 0.001). (A): percentage areas of fungal infection. One-way ANOVA detected the significant effect of time (F10, 22 = 200.85, p < 2.2×10−16). (B): The thickness of the cuticle after Beauveria bassiana exposure. One-way ANOVA detected the significant effect of time (F10, 99 = 612.48, p < 2.2×10−16).
Histological Alteration Index (HAI)
The histological alteration index (HAI) was determined to evaluate the effect of fungal infection. Compared to the normal tissue of H. hampei (Figure 5(A)), three major histopathological alterations were observed, including thinning of the adipose layer, degeneration of muscle fibers, and erosion of the gut epithelium (Figures 5(B) and 5(C)). All three alterations showed a time-dependent increase. The most severe thinning of adipose tissue, disintegration of muscle fibers, and compromise of gut epithelial integrity were noted at 144 h post-exposure, reaching the highest severity score of 3 (Figure 5).
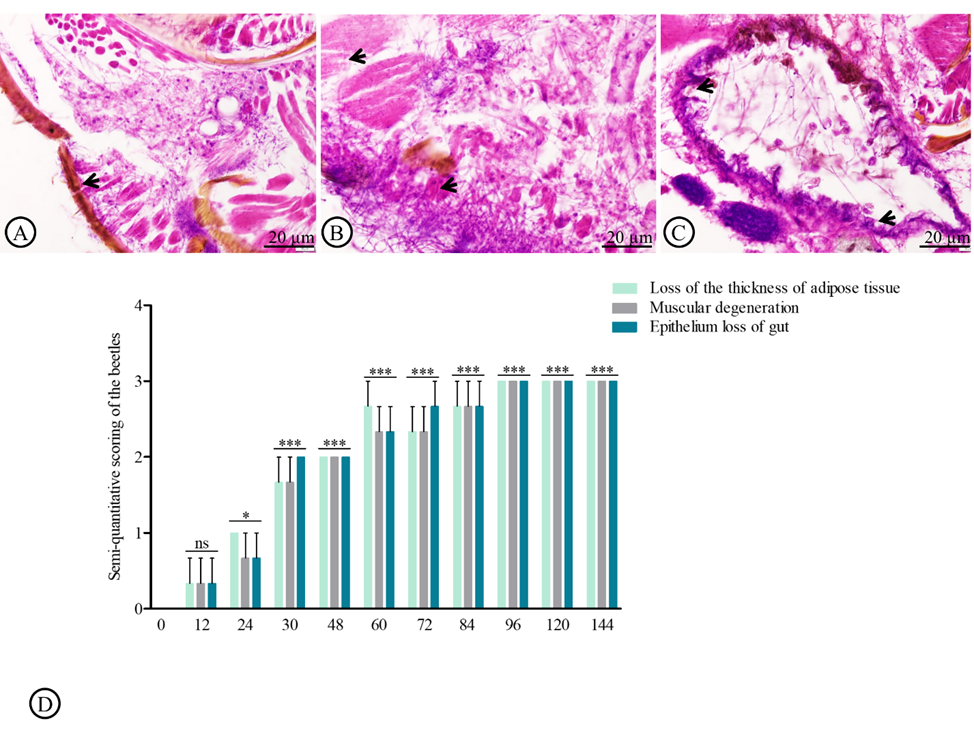
Figure 5 Histopathological alterations index (HAI) in adult Hypothenemus hampei infected with Beauveria bassiana. The normal structure of integument (arrow, A), muscular degeneration (arrow, B), and gut epithelium loss (arrow, C) were assessed in a semiquantitative manner. Values are represented as means ± SE.
Semiquantitative evaluation of the TUNEL-positive cells
Immediately after exposure to B. bassiana, the tissue of H. hampei was primarily unaltered with a sparse presence of apoptotic cells, which indicated a minimal immune response (Figure 6(A)). At 12 h post-exposure, there was a slight increase in the number of TUNEL-positive apoptotic cells (Figure 6(B) and Table 1). At 48 h post-exposure, a marked increase in apoptotic cells was detected (Figure 6(C) and Table 1), followed by observation of high immunoreactivity levels from 60 to 144 h post-exposure (Figures 6(D) - 6(E) and Table 1).
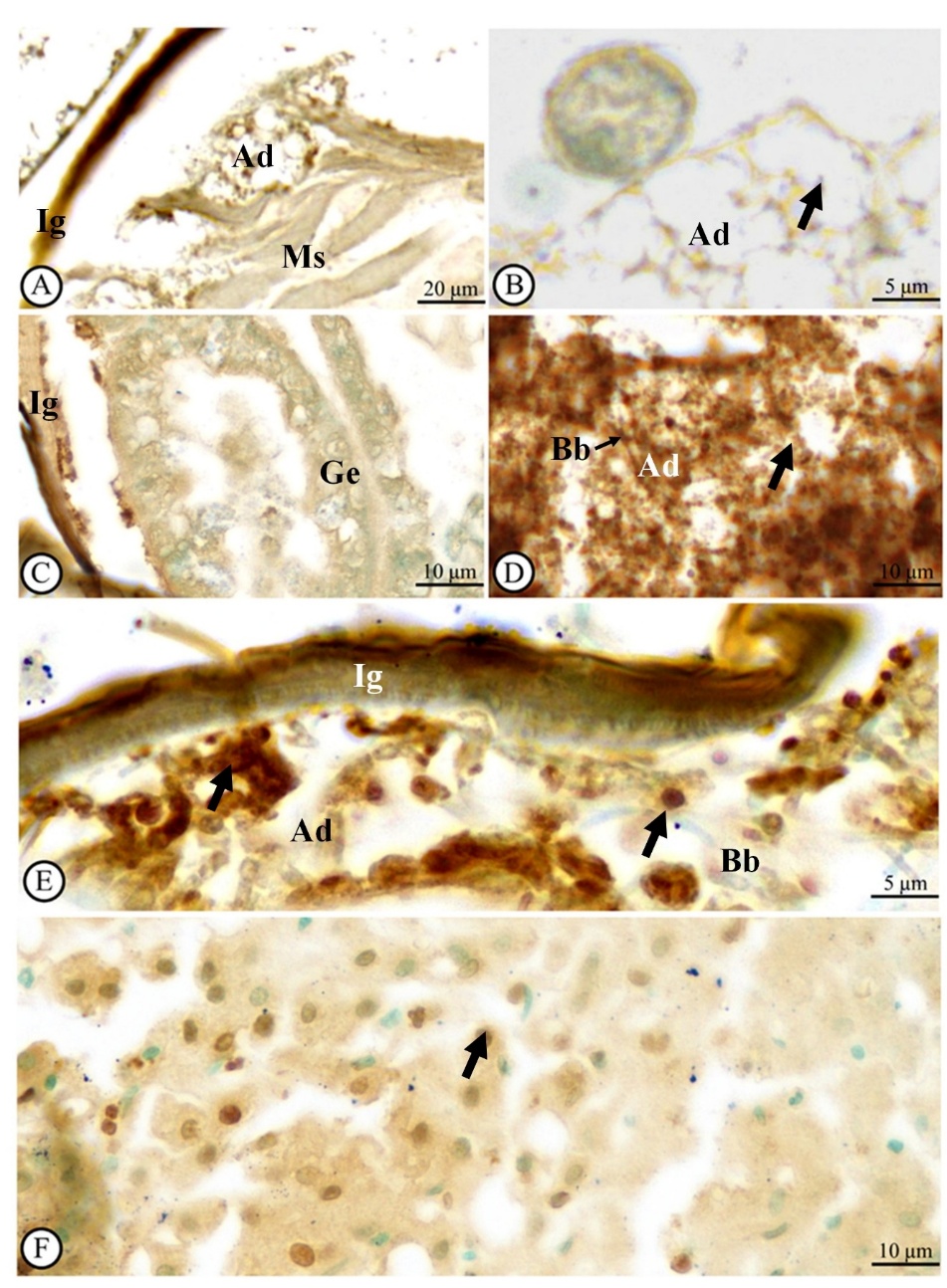
Figure 6 Light microscope observations of apoptotic cells in an adult Hypothenemus hampei infected with Beauveria bassiana (Bb) at 0 (A), 12 (B), 48 (C), 84 (D), and 120 h (E) post-exposure. Results from TUNEL staining method showed the brown signals as well as the TUNEL-positive cells [black arrows indicate the apoptotic cell]. Note: Liver tissue from human body as positive tissue (F). Abbreviations: Ad = adipose tissue, Ig = integument, Ms = muscle, Ge = gut epithelium.
Our observation revealed that B. bassiana exhibited rapid infection dynamics in H. hampei, with fungal penetration observed as early as 12 h post-exposure. This early invasion of the cuticle and adipose tissue indicates a high infectivity potential, which is critical for effective biological control. It is possible for a better understanding of the interaction between the specialized adipose tissue of sampled insects and EPFs. Therefore, this question should be investigated about the fungal tropism and adipose tissue in further observation.
Table 1 Semiquantitative analytical score of TUNEL-positive cell numbers at each time point in hours.
Title 1 |
0 |
12 |
48 |
60 |
84 |
96 |
120 |
144 |
TUNEL positive cells |
- |
+ |
++ |
+++ |
+++ |
+++ |
+++ |
+++ |
On the semiquantitative analytical scale, ‘−’ indicates no TUNEL-positive signal, ‘+’ weak TUNEL-positive signal, ‘++’ moderate TUNEL-positive signal, and ‘+++’ strong TUNEL-positive signal.
Progressive tissue degradation, including cuticle thinning, muscle disintegration, and gut epithelial erosion, together with increased apoptosis in H. hampei, suggests that fungal infection disrupts multiple physiological systems and immune responses of the sampled insects under a defensive host response, ultimately leading to host mortality. Compared with previous studies reporting fungal germination and infection at later time points [23], the earlier penetration observed in this study highlights the potential of this fungal strain for rapid action against the coffee berry borer. Such rapid infection dynamics are advantageous for field application, where timely suppression of pest populations is essential. These findings provide important insights into the mechanisms underlying the pathogenicity of B. bassiana and support its use as a biological control agent. Understanding the timing of fungal invasion and tissue damage can help optimize application strategies, such as timing and formulation, to enhance efficacy under field conditions. However, environmental factors such as temperature, humidity, and UV exposure may influence fungal performance, and should be considered when applying this biocontrol agent in coffee-based agroecosystems [24].
In this study, fungal conidia were easily identifiable within the cuticle layer of H. hampei. The penetration of the germ tube into the insect’s cuticle is primarily facilitated by the secretion of hydrolytic enzymes, including lipases, proteases, and chitinases. These extracellular cuticle-degrading enzymes play a crucial role in breaking down the insect’s protective barrier [25]. In addition to these enzymes, secondary metabolites such as destruxin, bavericin, and efrapeptins, secreted by Beauveria species, contribute to the pathogenic process by aiding in tissue destruction and immune suppression within the host. These compounds not only degrade the cuticle but also impair the physiological functions of the insect, eventually leading to its death [26].
We hypothesize that the enzymatic breakdown of the cuticle and subsequent invasion of the insect body are critical factors in inducing mortality. This aligns with previous research showing that mechanical damage to the insect’s cuticle, in combination with enzymatic action, is often a key contributor to insect death [25]. Additionally, Khachatourians [27]. suggested that entomopathogenic fungi have components capable of chemically digesting the insect cuticle, further facilitating fungal invasion. This combination of mechanical and enzymatic degradation likely overwhelms the insect’s defenses, leading to rapid decline and eventual death.
In the context of the host’s cellular response, while apoptosis is traditionally viewed as a defensive mechanism to sequester pathogens, the massive induction of cell death observed in this study may actually facilitate fungal colonization by rapidly breaking down host tissues and reducing biological resistance, a strategy also observed in other hypocrealean fungi attacking H. hampei [28]. Thus, B. bassiana not only penetrates the insect cuticle but also disrupts vital physiological processes, providing a comprehensive mechanism for pest control. The insights from this study could enhance the development of more effective biocontrol strategies, particularly in pest management systems where chemical resistance is becoming an issue. Understanding the infection dynamics of B. bassiana can aid in optimizing environmental conditions to maximize fungal efficacy, ultimately contributing to the sustainable management of agricultural pests like H. hampei.
Conclusions
The findings of this study provide compelling evidence of the effectiveness of Beauveria bassiana as a biocontrol agent against H. hampei. The fungus rapidly invades and degrades internal tissues, leading to cuticle thinning, extensive tissue damage, and increased apoptosis, which are key factors contributing to insect mortality. These insights into infection dynamics, including the timing of fungal penetration and tissue disruption, can be used to optimize application strategies and improve the efficacy of biocontrol formulations. Consequently, this approach may reduce reliance on chemical insecticides, lower environmental contamination, and support ecological balance in coffee-based agroecosystems. This aligns with the study’s initial rationale of promoting sustainable pest management through the use of entomopathogenic fungi.
Acknowledgements
The authors thank the Microtechnique Laboratory (MIC-LAB), Division of Biological Sciences, Faculty of Science, Prince of Songkla University, Thailand for technical support and equipment use in this experiment. We would like to thank Ms. Kitaya Kongtong for her laboratory support and for preparing the histological figures and thank Dr. Tommy Coyne for his check and correction of the grammatical structure. This research is supported by the Center of Excellence on Agricultural Biotechnology, Office of the Permanent Secretary, Ministry of Higher Education, Science, Research and Innovation (AG-BIO/MHESI). The project was granted by the Foundation for Research in Agricultural and Natural Resources, Biotechnology Phase 3, Faculty of Natural Resources, Prince of Songkla University. Also, this research was funded by Thailand Science Research and Innovation (TSRI) grant No. 672A01014 from the Fundamental Fund for Basic Research, National Research Council of Thailand.
Declaration of generative AI in scientific writing
The authors acknowledge the use of generative AI tools to check the referencing style of this manuscript. No content generation or data interpretation was performed by AI. The authors take full responsibility for the content and conclusions of this work.
CRediT author statement
Sinlapachai Senarat: Conceptualization; Methodology; Validation; Writing - Original Draft; Writing - Review & Editing. Narit Thaochan: Conceptualization; Methodology; Validation; Writing - Original Draft; Writing - Review & Editing; Supervision; Project administration. Kodeeyah Thoawan: Investigation. Peerasak Bunsap: Software; Investigation. Kitipong Angsujinda: Investigation. Pisit Poolprasert: Investigation; Writing - Original Draft. Anjaree Inchan: Investigation. Natthawut Charoenphon: Investigation; Writing - Original Draft. Supapong Imsonpang: Investigation. Gen Kaneko: Original Draft; Writing - Review & Editing.
References
[1] MA Johnson, CP Ruiz-Diaz, NC Manoukis and JCV Rodrigues. Coffee berry borer (Hypothenemus hampei), a global pest of coffee: Perspectives from historical and recent invasions, and future priorities. Insects 2020; 11(12), 882.
[2] JF Barrera. Coffee pests and their management. In: JL Capinera (Ed.). Encyclopedia of entomology. Springer, Dordrecht, Netherlands, 2008, p. 961-998.
[3] P Benavides, C Góngora and A Bustillo. IPM program to control coffee berry borer Hypothenemus hampei, with emphasis on highly pathogenic mixed strains of Beauveria bassiana, to overcome insecticide resistance in Colombia. In: F Perveen (Ed.). Insecticides - advances in integrated pest management. IntechOpen, London, 2012, p. 511-540.
[4] A Buranapanitchaphan and C Yaowaluk. Infestation of coffee berry borer and controlling efficiency of attractants. Journal of Agricultural Science 2014; 30, 223-231.
[5] N Thaochan and A Chandrapatya. The phenotypic and metabolic properties of Metarhizium guizhouense on Corcyra cephalonica. Mycosphere 2016; 7(2), 214-225.
[6] N Thaochan and A Ngampongsai. Effects of autodisseminated Metarhizium guizhouense PSUM02 on mating propensity and mating competitiveness of Bactrocera cucurbitae (Diptera: Tephritidae). Biocontrol Science and Technology 2015; 25(6), 629-644.
[7] N Thaochan, A Ngampongsai, CS Prabhakar and Q Hu. Beauveria bassiana PSUB01 simultaneously displays biocontrol activity against Lipaphis erysimi (Kalt.) (Hemiptera: Aphididae) and promotes plant growth in Chinese kale under hydroponic growing conditions. Biocontrol Science and Technology 2021; 31(10), 997-1015.
[8] W De LaRosa, R Alatorre, JF Barrera and C Toriello. Effect of Beauveria bassiana and Metarhizium anisopliae (Deuteromycetes) upon the coffee berry borer (Coleoptera: Scolytidae) under field conditions. Journal of Economic Entomology 2000; 93(5), 1409-1414.
[9] S Paudel, S Mansfield, LF Villamizar, TA Jackson and SDG Marshall. Can biological control overcome the threat from newly invasive coconut rhinoceros beetle populations (Coleoptera: Scarabaeidae)? A review. Annual of the Entomological Society of America 2021; 114(2), 247-256.
[10] LFC Ribeiro, J Tavares, SAV Silva, LFAA Alvez, EA Loth and RMC Brancalhão. Infection of silkworm larvae by the entomopathogenic fungus Metarhizium anisopliae. Crop Protection 2017; 47(4), e20151485.
[11] Y Duan, H Wu, Z Ma, L Yang and D Ma. Scanning electron microscopy and histopathological observations of Beauveria bassiana infection of Colorado potato beetle larvae. Microbial Pathogenesis 2017; 111, 435-439.
[12] CM Lacey, LA Lacey and DR Roberts. Route of invasion and histopathology of Metarhizium anisopliae in Culex quinquefasciatus. Journal of Invertebrate Pathology 1988; 52(1), 108-118.
[13] R Ibrahim, S Alahmadi, YS Binnaser and D Shawer. Seasonal prevalence and histopathology of Beauveria bassiana infecting larvae of the leopard moth, Zeuzera pyrina L. (Lepidoptera: Cossidae). Egyptian Journal of Biological Pest Control 2019; 29(1), 65.
[14] SM Smith, RA Beaver and AI Cognato. A monograph of the Xyleborini (Coleoptera, Curculionidae, Scolytinae) of the Indochinese Peninsula (except Malaysia) and China. ZooKeys 2020; 983, 1-442.
[15] Thaochan, A Ngampongsai, CS Prabhakar and Q Hu Q. Beauveria bassiana PSUB01 simultaneously displays biocontrol activity against Lipaphis erysimi (Kalt.) (Hemiptera: Aphididae) and promotes plant growth in Chinese kale under hydroponic growing conditions. Biocontrol Science and Technology 2021; 31(10), 997-1015.
[16] SK Suvarna, C Layton and JD Bancroft. Bancroft’s theory and practice of histological techniques. Elsevier, Amsterdam, Netherlands, 2013, p. 40-95.
[17] A Pintong, S Ampawong, N Komalamisra, P Sriwichai, S Popruk and J Ruangsittichai. Insecticidal and histopathological effects of Ageratum conyzoides weed extracts against dengue vector, Aedes aegypti. Insects 2020; 11(4), 224.
[18] DR Dietrich and HO Krieger. Histological analysis of endocrine disruptive effects in small laboratory fish. John Wiley & Sons, New Jersey, United States, 2009, p. 254-265.
[19] MC Rojo and ME Gonzalez. In situ detection of apoptotic cells by TUNEL in the gill epithelium of the developing brown trout (Salmo trutta). Journal of Anatomy 1998; 193(3), 391-398.
[20] DW Roberts and RA Humber. Etomogenous fungi. In: GT Cole and WB Kendrick (Eds.). The biology of conidial fungi. Academic Press, New York, 1981, p. 201-236.
[21] RA de Souza, D Pratissoli, LM de A Junior, J de A Pinheiro, JO Ataíde, LAG Tamashiro, MA dos Santos and AP Damascena. Mortality of coffee berry borer, Hypothenemus hampei in field, with pre and post application of entomopathogenic fungus Beauveria bassiana (Balsamo) associated to emulsifiers. International Journal of Advanced Engineering Research and Science 2020; 7(5), 80-85.
[22] FM Chang, HL Lu and YS Nai. Evaluation of potential entomopathogenic fungus, Beauveria bassiana, for controlling the coffee berry borer Hypothenemus hampei (Ferrari) (Coleoptera: Curculionidae) in Taiwan. Journal of Asia-Pacific Entomology 2023; 26(4), 102118.
[23] Y Liu, Y Yang and B Wang. Entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae play roles of maize (Zea mays) growth promoter. Scientific Reports 2022; 12(1), 15706.
[24] AV Toledo, AMM de Remes Lenicov and CC López Lastra. Histopathology caused by the entomopathogenic fungi, Beauveria bassiana and Metarhizium anisopliae, in the adult planthopper, Peregrinus maidis, a maize virus vector. Journal of Insect Science 2010; 10(1), 35.
[25] B Deka, C Baruah and A Babu. Entomopathogenic microorganisms: their role in insect pest management. Egyptian Journal of Biological Pest Control 2021; 31(1), 121.
[26] G Xiao, SH Ying, P Zheng, ZL Wang, S Zhang, XQ Xie, Y Shang, RJ Leger, GP Zhao, C Wang and MG Feng. Genomic perspectives on the evolution of fungal entomopathogenicity in Beauveria bassiana. Scientific Reports 2012; 2(1), 483.
[27] AE Hajek and RJ St Leger. Interactions between fungal pathogens and insect hosts. Annual Review of Entomology 1994; 39(1), 293-322.
[28] S Senarat, P Bunsap, P Poolprasert, A Inchan, N Charoenphon, P Sornying and N Thaochan. Histopathological study of host-pathogen interactions between Cordyceps javanica PSUC002 and Hypothenemus hampei. Journal of Fungi 2025; 11(6), 423.