Trends
Sci.
2026;
23(10):
13265
Phytochemical Screening and Antioxidant Activities of Baeckea frutescens Branches Ethanolic Extract on RAW 264.7 Cells
Nurhanani Ayub1, Salfarina Ramli1,2, Ihsan Safwan Kamarazaman3,
Evana Kamarudin4, Richard Johari James1,2, Nurul Badriah5 and Hasseri Halim1,2,*
1Faculty of Pharmacy, Universiti Teknologi MARA Selangor, Puncak Alam 42300, Malaysia
2Integrative Pharmacogenomic Institute (iPROMISE), Universiti Teknologi MARA Selangor,
Puncak Alam 42300, Malaysia
3Forest Research Institute Malaysia, Selangor Darul Ehsan 52109, Malaysia
4Centre for Medical Laboratory Technology Studies, Faculty of Health Sciences, Universiti Teknologi MARA,
Puncak Alam Campus, Selangor 42300, Malaysia
5Department of Biomedical Science, Faculty of Medicine, Universitas Andalas, Padang 25175, Indonesia
(*Corresponding author’s e-mail: hasseri2945@uitm.edu.my)
Received: 5 January 2026, Revised: 15 March 2026, Accepted: 22 March 2026, Published: 30 April 2026
Reactive oxygen species (ROS) at low concentrations play an important role in cellular signaling, immune function, and cell proliferation. However, excessive ROS can damage cellular components and induce inflammation, contributing to the progression of diseases such as atherosclerosis, cancer, and rheumatoid arthritis. This study examines the phytochemical composition and antioxidant potential of an ethanolic extract from Baeckea frutescens L. branches (BFBE), using RAW 264.7 murine macrophages as the experimental model. The phytochemical screening was conducted using standard tests while the antioxidant activity of the ethanolic extract was evaluated using the DPPH and FRAP assays. Additionally, the study measured intracellular ROS levels and conducted antioxidant enzyme assays to determine the antioxidant capacity of the Baeckea frutescens branches ethanolic extract. Phytochemical screening revealed that the extract contains high levels of saponins and hydrolysable tannins, moderate levels of flavonoids, and a lower concentration of steroids. Alkaloids and triterpenoids were not detected. In the DPPH assay, the extract exhibited concentration-dependent free radical scavenging activity, with an IC₅₀ value of 86.16 μg/mL. The FRAP assay further supported its antioxidant potential, showing a gallic acid equivalent of 31.91 μM at 1,000 μg/mL of extract. Beyond these chemical assays, BFBE demonstrated a potential dual antioxidant effect within the macrophage model. It not only directly scavenged ROS but also helped maintain superoxide dismutase (SOD) activity under elevated ROS generation induced by hydrogen peroxide (H₂O₂), while no significant changes were observed in catalase and glutathione peroxidase activities. These findings suggest that the antioxidant effects of BFBE may involve both chemical antioxidant activity and modulation of cellular antioxidant responses. The study supports the potential of BFBE as a natural therapeutic agent for managing elevated ROS generation and oxidative stress. Further research is recommended to elucidate its molecular mechanisms and evaluate its in vivo efficacy.
Keywords: Baeckea frutescens, Phytochemical, Antioxidant, Hydrogen peroxide, Superoxide dismutase, DCFH-DA, RAW 264.7
Introduction
Oxidative stress, characterized by an imbalance between the generation of reactive oxygen species
(ROS) and the capacity of the antioxidant defense system, plays a pivotal role in the pathogenesis of numerous chronic and degenerative diseases. Excessive ROS production can damage cellular components such as proteins, lipids, and nucleic acids, ultimately leading to inflammation, cellular dysfunction, and apoptosis. Conditions such as cancer, cardiovascular diseases, diabetes, and neurodegenerative disorders have all been linked to oxidative stress-mediated mechanisms [1,2].
Given the harmful effects of ROS, there is increasing interest in identifying and utilizing natural compounds with potent antioxidant properties to mitigate oxidative damage. Plant-derived bioactive compounds, particularly those rich in polyphenols, flavonoids, tannins, and saponins, have shown promising protective effects against elevated ROS generation and oxidative stress-related cellular damage. These secondary metabolites exert their activity through free radical scavenging, metal chelation, and enhancement of endogenous antioxidant enzyme systems such as superoxide dismutase (SOD), catalase, and glutathione peroxidase (GPx) [3,4].
Baeckea frutescens, known as cucur atap in Malay, is a member of the Myrtaceae family and a shrub native to Southeast Asia, Australia, and New Guinea. In Malaysia, B. frutescens can be found on mountain tops, quartz ridges, and the sandy coasts of the eastern parts of the Peninsula. Known for its traditional medicinal uses, B. frutescens has been employed in various cultures to treat ailments such as fever, headache, rheumatism, and dysentery. B. frutescens has been traditionally utilized in Malaysia for postpartum care, including during confinement and in massages to alleviate body aches and numbness. Additionally, it has been employed for managing rheumatism [5]. In China, it is brewed as a herbal tea to alleviate fever and sunstroke [6]. B. frutescens is rich in a variety of bioactive compounds, including flavonoids, phloroglucinols, sesquiterpenoids, and essential oils. Approximately 130 metabolites have been isolated from this plant, with phloroglucinol derivatives and tasmanone being notable for their pharmacological activities. These compounds contribute to the plant’s antioxidant, anti-inflammatory, antimicrobial, and cytotoxic properties, making it a valuable resource for therapeutic applications [6].
Macrophages play a central role in the immune response and are major contributors to inflammation-associated oxidative stress through the production of reactive oxygen species (ROS) and reactive nitrogen species (RNS) during immune activation [7-9]. RAW 264.7 murine macrophage cells are widely used as an in vitro model for studying ROS generation, oxidative stress and inflammation because they are capable of generating intracellular ROS and modulating antioxidant defense mechanisms in response to various stimuli, such as lipopolysaccharide (LPS) or hydrogen peroxide [10,11]. Therefore, this cell line provides a physiologically relevant model to evaluate the antioxidant and cytoprotective effects of natural compounds under conditions that mimic inflammation-associated oxidative stress [12,13].
This study aims to investigate the antioxidant potential of ethanolic extracts from B. frutescens branches through both chemical and in vitro cellular assays. The extract’s ability to scavenge free radicals and enhance antioxidant enzyme activities was assessed using DPPH and FRAP assays, as well as intracellular ROS and enzyme modulation studies in RAW 264.7 murine macrophages. By utilizing RAW 264.7 macrophages as a model of immune-related ROS generation, this study seeks to provide further scientific validation for the potential of B. frutescens as a natural therapeutic agent against ROS-induced cellular damage.
Material and methods
Reagents and chemicals
We used 95% ethanol, ethyl ether, hydrochloric acid (HCl), 70% ethanol, ferric chloride (FeCl3), absolute ethanol, magnesium ribbon, chloroform (CHCl3), acetic anhydride (C4H6O3), sulfuric acid (H2SO4), ammoniacal chloroform, 2,2-diphenyl-1-picrylhydrazyl (DPPH), gallic acid (C7H6O5), sodium acetate trihydrate (C2H9NaO5), glacial acetic acid, 2,4,6-tripyridyl-s-triazine (TPTZ), ferric chloride hexahydrate (FeCl₃·6H₂O), Dulbecco’s Modified Eagle Medium (DMEM), fetal bovine serum (FBS), phosphate buffered saline (PBS), penicillin-streptomycin, dimethyl sulfoxide (DMSO), 2′,7′-dichlorodihydrofluorescein diacetate (DCFH-DA), Superoxide Dismutase Kit (Cayman Chemical Company, cat. #706002), Glutathione Peroxidase Assay Kit (Cayman Chemical Company, cat. #703102) and Catalase Assay Kit (Cayman Chemical Company, cat. #707002) and hydrogen peroxide (H₂O₂).
Plant material collection and extraction
Branches of B. frutescens, collected from Setiu, Kuala Terengganu, Terengganu, Malaysia, were taxonomically verified by a botanist from the Forest Research Institute Malaysia. A voucher specimen (SBID:044/20) was catalogued and stored at the Faculty of Pharmacy, Universiti Teknologi MARA, Selangor. Post-authentication, the branches were air-dried and ground into fine powder. The extraction process followed a modified protocol [14]. Specifically, 500 g of the powdered material was extracted three times, each for 72 h, using 2.5 L of 95% ethanol. The pooled extracts were filtered and concentrated under reduced pressure using a rotary evaporator at 40 °C and approximately 150 - 200 mbar until complete solvent removal. The resulting crude ethanolic extract was kept at –80 °C for subsequent experiments. The extraction yield was calculated for each iteration. The yield of each extraction was calculated using the following formula:
For the non-cell-based antioxidant assays (DPPH and FRAP), the crude extract was dissolved in distilled water prior to analysis. For the cell-based experiments, the crude extract was dissolved in Dulbecco’s Modified Eagle Medium (DMEM) before treatment. A vehicle control consisting of the respective solvent (distilled water or DMEM) alone was included in all experiments to ensure that the solvent did not influence the measured outcomes.
Phytochemical analysis
Phytochemical screening of B. frutescens branches ethanolic extract (BFBE) was conducted to detect major classes of secondary metabolites which include alkaloid, saponin, tannin, triterpenoid and steroid content, accordingly [15]. All tests were performed in four independent biological replicates (n = 4).
Antioxidant analysis
DPPH assay
The antioxidant capacity of BFBE was evaluated by measuring its DPPH radical scavenging activity. The assay was performed according to the protocol [14]. Briefly, 50 µL of branch extract at various concentrations was added to 150 µL of 0.3 mM DPPH ethanolic solution in a 96-well plate. Following a 30-minute incubation in the dark at room temperature, the absorbance of each well was measured at 515 nm using a microplate reader. The assay was performed in four independent biological replicates (n = 4). Scavenging activity was then calculated, and IC50 values were determined using GraphPad Prism 9 software. Ethanol served as the blank, and ascorbic acid was used as a positive control. DPPH scavenging activity was calculated using this formula:

FRAP assay
The ferric reducing antioxidant power of BFBE was determined according to the previous study [16]. The FRAP reagent was freshly prepared by combining 300 mM acetate buffer (pH 3.6), 10 mM TPTZ solution, 20 mM ferric chloride solution, and distilled water in a 10:1:1:1 ratio. For the assay, 3 µL of branch extract or gallic acid (at various concentrations) was added to a 96-well plate, followed by 100 µL of FRAP reagent. After a 30-minute incubation at 37 °C in the dark, the absorbance was measured at 593 nm using a microplate reader. All measurements were performed in four independent biological replicates (n = 4). Gallic acid served as a standard, and the results were expressed as gallic acid equivalents, calculated using GraphPad Prism 9 software. Ascorbic acid (vitamin C) was used as the positive control for the chemical antioxidant assays (DPPH and FRAP) because it is a well-established water-soluble radical scavenger commonly used as a reference standard in these assays.
Cell culture
RAW 264.7 murine macrophages (ATCC® TIB-71™, American Type Culture Collection, USA) were grown in Dulbecco’s Modified Eagle Medium (DMEM) enriched with 10% fetal bovine serum (FBS) and 1% penicillin-streptomycin. The cells were incubated at 37 °C in a humidified environment with 5% CO₂, following the protocol outlined [17]. Cells between passages 5 - 15 were used for all experiments. Routine sub-culturing was carried out once the cells reached approximately 80% - 90% confluency.
Cell viability assay
Cell viability was assessed through the MTT assay, which measures the metabolic activity of viable cells via the reduction of the yellow tetrazolium salt MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) into insoluble purple formazan by mitochondrial enzymes. RAW 264.7 cells were seeded in a 96-well plate at a density of 10,000 cells/well and incubated overnight at 37 °C, after which the cells were exposed to their respective treatment conditions and the culture medium was carefully removed. The crude extract was dissolved in Dulbecco’s Modified Eagle Medium (DMEM) prior to treatment. A vehicle control consisting of DMEM alone was included in the all cell-based experiments to ensure that the solvent did not affect cell viability or intracellular ROS levels. The cells were then incubated in serum-free DMEM containing 0.5 mg/mL MTT at 37 °C for 4 h. Following incubation, the MTT solution was discarded, and dimethyl sulfoxide (DMSO) was added to dissolve the formed formazan crystals. This assay was performed in four independent biological replicates (n = 4). The absorbance was recorded at 540 nm using a microplate reader, and cell viability was expressed as a percentage of the untreated control using the calculation formula below.
Cell Viability (%) = (Absorbance of treated cells / Absorbance of control cells) × 100 (3)
The concentrations for subsequent cell-based assays (3.125 - 100 µg/mL) were selected based on the MTT results, utilizing subtoxic doses that maintained cell viability above 90% to ensure the observed physiological responses were not influenced by cytotoxicity.
Antioxidant capacity
Intracellular reactive oxygen species
Intracellular reactive oxygen species levels were measured according to the method [18]. RAW 264.7 cells were seeded in a 96-well black plate at a density of 10,000 cells/well and incubated overnight at 37 °C. Cells were then treated with varying concentrations of BFBE (3.125 - 100 µg/mL) in serum-free media for 24 h. After washing with serum-free media, cells were incubated with 20 µM DCFH-DA for 30 min at 37 °C. Following two washes with PBS, 200 µL of 1 mM H₂O₂ in PBS was added, and the cells were incubated for an additional 20 min at 37 °C. Fluorescence intensity was measured at an excitation wavelength of 485 nm and an emission wavelength of 530 nm using a fluorescence microplate reader. Data analysis was performed using GraphPad Prism 9 software. Data were obtained from four independent biological replicates (n = 4).
Antioxidant enzyme assay
The activities of the antioxidant enzymes superoxide dismutase, glutathione peroxidase, and catalase were measured using commercially available kits (Catalase Assay Kit (Cayman Chemical, cat. no. 707002), Superoxide Dismutase Assay Kit (cat. no. 706002), and Glutathione Peroxidase Assay Kit (cat. no. 703102). RAW 264.7 cells were seeded in 6-well plates and incubated overnight at 37 °C. Following a 24-hour treatment with various concentrations of BFBE (12.5 - 50 µg/mL), cells were exposed to 1 mM H₂O₂ in PBS for 30 min at 37 °C. After washing with cold phosphate buffer, cells were harvested, collected, and centrifuged. The supernatant was discarded, and the cell lysates were stored at –80 °C until further analysis. All enzyme assays were performed in four independent biological replicates (n = 4) according to the manufacturer’s instructions. α-tocopherol (vitamin E) was used as the positive control for the cell-based assays (intracellular ROS and antioxidant enzyme studies) due to its biological relevance as a lipid-soluble antioxidant that protects cellular membranes from oxidative stress. Absorbance readings were obtained using a microplate reader, and data analysis was performed using GraphPad Prism 9 software.
Statistical analysis
All experiments were performed in 4 independent biological replicates (n = 4). Data are expressed as mean ± standard error of the mean. Statistical analysis was performed using a one-way analysis of variance followed by Dunnett’s post-hoc test for multiple comparisons against the control group using with GraphPad Prism 9 software. A p - value less than 0.05 was considered statistically significant.
Result and discussion
Result
Baeckea frutescens branches ethanolic extract (BFBE)
Table 1 presents the yield of ethanolic crude extract obtained from B. frutescens L. branches using the maceration technique. The average yield from two extractions was 8.84 ± 0.28%, with the first extraction yielding 9.04% and the second 8.64%.
Table 1 Yield of the plant extraction.
Extraction (n = 2) |
Dry branches used (g) |
Extract obtained (g) |
Yield (%) |
Average yield (%) |
1 |
500.0 |
45.201 |
9.04% |
8.84 ± 0.28 |
2 |
500.0 |
43.231 |
8.64% |
Phytochemical screening of BFBE
Table 2 shows the phytochemical components of BFBE. The phytochemical analysis of the ethanolic extract from the branches of B. frutescens revealed a high content of saponins and hydrolysable tannins. Additionally, the extract exhibited moderate levels of flavonoids and a lower concentration of steroids. However, no detectable amounts of alkaloids and triterpenoids were found in this phytochemical screening.
Table 2 Phytochemical components of Baeckea frutescens branches ethanolic extract (BFBE).
Phytochemicals classes |
Ethanolic extract of Baeckea frutescens branches extract |
Alkaloid |
- |
Saponin |
+++ |
Tannin |
+++ (H) |
Flavonoids |
++ |
Triterpenoid |
- |
Steroid |
+ |
H = Hydrolysable tannin, +++ High concentration, ++ Moderate concentration, + low concentration, - not detected.
Antioxidant capacities of BFBE
The antioxidant capacities of BFBE were evaluated through radical scavenging activity, assessed by the DPPH assay, and ferric ion reduction, measured by the FRAP assay. Figure 1(A) illustrates the antioxidant activities of BFBE at various concentrations. The radical scavenging activity of the extract was tested at concentrations ranging from 1,000 to 31.25 µg/mL, using ascorbic acid as the positive control. The results indicate that the radical scavenging properties of BFBE can scavenge free radicals in a dose-dependent manner, with higher extract concentrations exhibiting greater radical scavenging activity. The highest concentration of BFBE, 1,000 µg/mL, demonstrated a 91.24% radical scavenging activity, while the lowest concentration, 31.25 µg/mL, exhibited a 33.36% radical scavenging activity. The median inhibitory concentration for BFBE was 86.16 µg/mL.
The FRAP assay was used to measure the total antioxidant activity by evaluating the reduction of ferric (Fe3+) ions to the ferrous (Fe2+) form. Figure 1(B) displays the gallic acid equivalent of the Baeckea frutescens L. branches ethanolic extract at various concentrations. The reducing antioxidant capacity was expressed in terms of gallic acid equivalence, with 125 µM ascorbic acid serving as the positive control. The BFBE was tested at concentrations ranging from 1,000 to 125 µg/mL. The results indicated that the extract could reduce ferric ions to ferrous ions in a dose-dependent manner. The gallic acid equivalent of the BFBE at 1,000 µg/mL was 31.91 µM, while the lowest extract concentration of 125 µg/mL had a gallic acid equivalent of 2.69 µM.
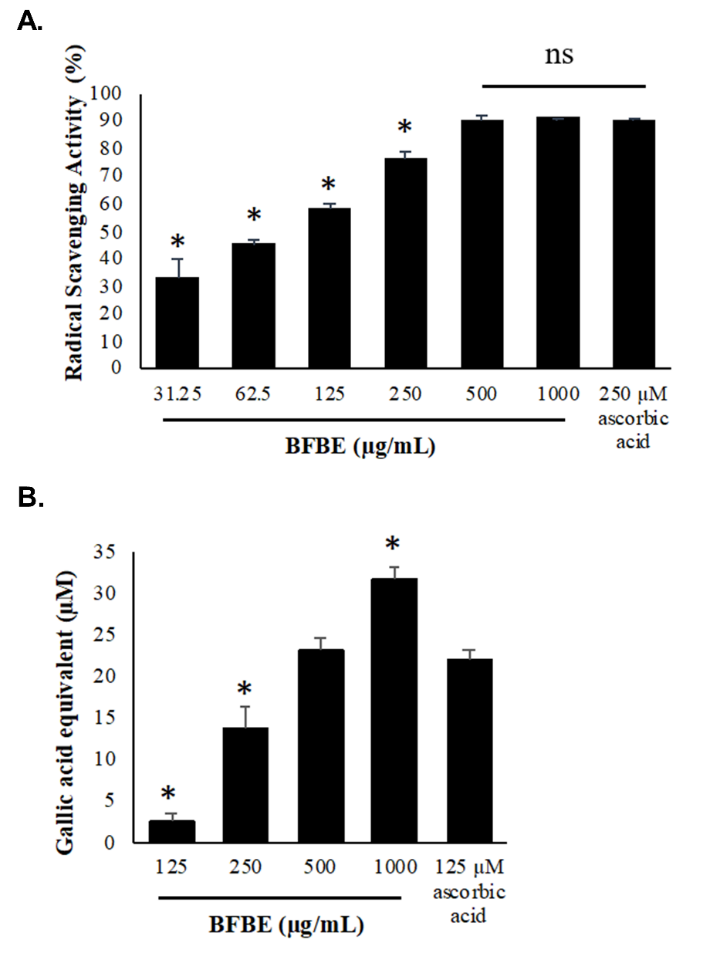
Figure 1 Antioxidant activities of Baeckea frutescens L. branches ethanolic extract (BFBE). (A) DPPH radical scavenging assay with 250 µM ascorbic acid positive control. (B) FRAP reducing power assay expressed as gallic acid equivalent with 125 µM ascorbic acid positive control. Data are presented as mean ± standard error of the mean from 4 independent biological replicates experiments (n = 4). Symbols denote statistical significance relative to ascorbic acid positive control: (*) indicates p < 0.05; NS indicates non-significant visual labels.
The high antioxidant capacity of the BFBE, as demonstrated by its DPPH radical scavenging and FRAP assay results, suggests that the extract could be a promising natural source of antioxidants [14,19]. This antioxidant potential is likely attributed to the presence of bioactive phytochemicals, such as phenolic compounds and flavonoids, which are known to significantly contribute to the antioxidant properties of plant extracts [20-31].
The antioxidant properties observed in the BFBE are consistent with findings from studies on other plant extracts, including those from Bouea macrophylla Griff leaves, Hieracium pilosella L., and Plumbago zeylanica roots [14,19,24]. These studies have reported the presence of various antioxidant compounds, such as polyphenols, flavonoids, and phenolic acids, which play a crucial role in scavenging free radicals and reducing ROS generation and oxidative stress [3,4,24,25].
Cell viability (MTT assay)
The cytotoxic effects of the BFBE and H₂O₂ on the viability of RAW 264.7 cells were assessed using the MTT assay [14,19,26]
.
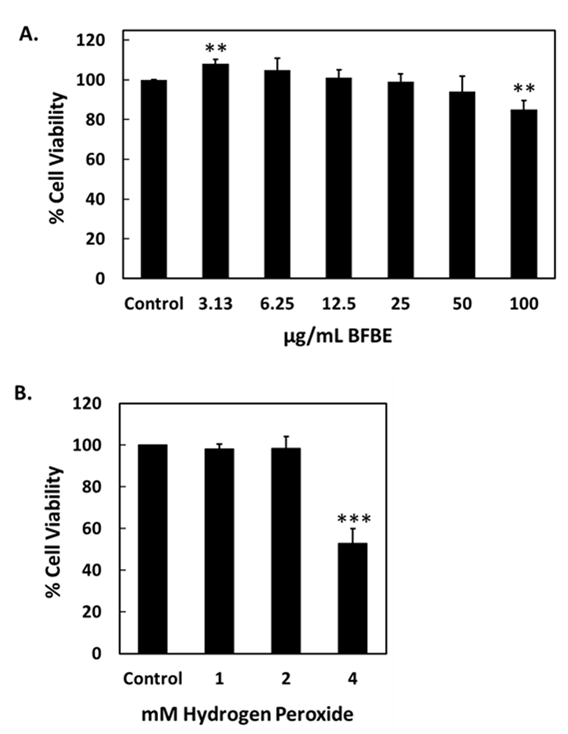
Figure 2 Impact of Baeckea frutescens branch ethanolic extract (BFBE) and H₂O₂ on RAW 264.7 cell viability. RAW 264.7 macrophages were treated with (A) BFBE (0 - 100 µg/mL) for 24 h and (B) H₂O₂ (0 - 4 mM) for 0.5 h. Viability was determined by MTT assay. Data are presented as mean ± S.D. from four independent experiments (n = 4). Statistical significance compared to the control group was determined using One-way ANOVA followed by Dunnett’s post-hoc test: ****p < 0.0001, ***p < 0.001, **p < 0.01, *p < 0.1, ns = not significant.
The results demonstrated that treatment with BFBE for 24 h produced concentration-dependent effects on RAW 264.7 cell viability (Figure 2(A)). At the lowest concentration (3.13 µg/mL), a significant increase in cell viability was observed (**p < 0.01), suggesting a possible proliferative effect. In contrast, the highest concentration tested (100 µg/mL) significantly reduced cell viability (**p < 0.01), indicating cytotoxicity at elevated doses. Intermediate concentrations (6.25 - 50 µg/mL) did not significantly differ from the control group, suggesting that BFBE is well tolerated within this range.
In contrast, treatment with H₂O₂ for 0.5 h showed a concentration-dependent reduction in RAW 264.7 cell viability (Figure 2(B)). A marked and significant decrease was only observed at 4 mM (***p < 0.001), while lower concentrations (1 - 2 mM) did not significantly impact cell viability compared to the control. This may indicate that although H₂O₂ induces ROS generation, a longer exposure duration might be required to observe cytotoxic effects at lower concentrations, as short-term treatment may not be sufficient to overwhelm cellular antioxidant defenses.
These findings suggest that BFBE has biphasic effects on macrophage viability by enhancing it at low doses and decreasing it at high doses, while H₂O₂-induced cytotoxicity appears to be both dose- and time-dependent. Further investigation into the mechanisms of action, particularly the involvement of oxidative stress and inflammation pathways, is warranted to better understand these cellular responses [26,27].
Antioxidant capacity
Intracellular reactive oxygen species
The antioxidant and free radical scavenging capabilities of plant-derived compounds have been extensively explored for their potential applications in various health conditions [1,2,28]. In the present study, RAW 264.7 cells were treated with BFBE at different concentrations and the intracellular ROS levels were assessed using the DCFH-DA assay.
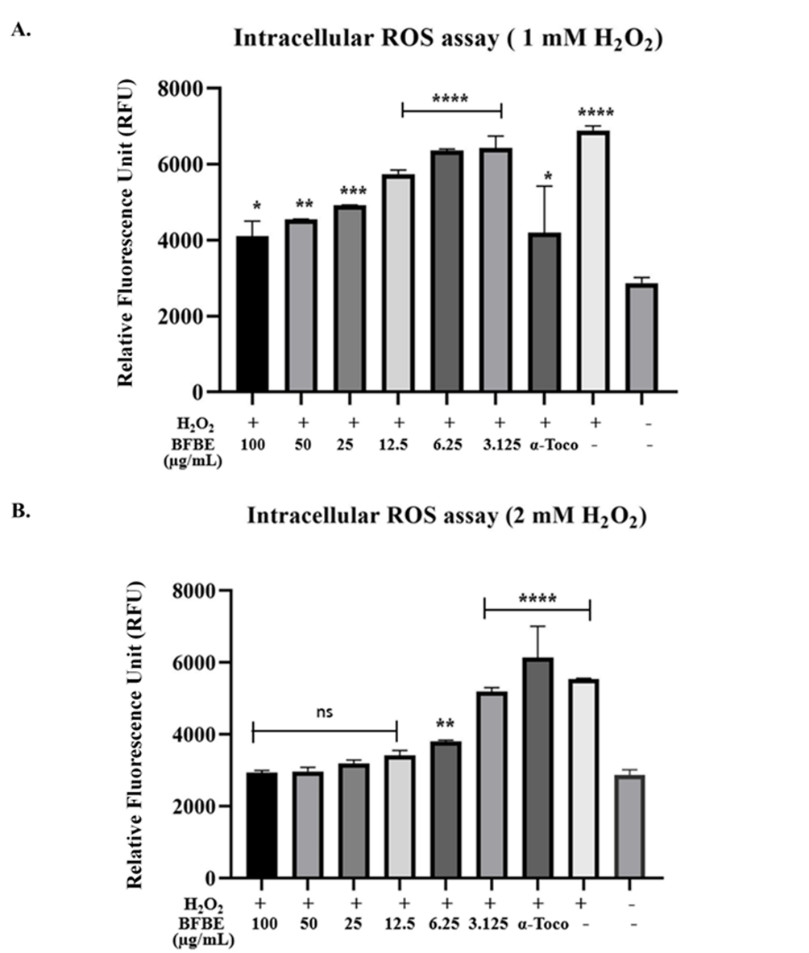
Figure 3 Effects of Baeckea frutescens branches ethanolic extract (BFBE) on intracellular Reactive Oxygen Species (ROS) levels in H₂O₂-exposed RAW 264.7 cells. RAW 264.7 cells were pretreated for 24 h with varying concentrations of BFBE (100, 50, 25, 12.5, 6.25, and 3.125 µg/mL) or α-Tocopherol (α-Toco, positive control), followed by exposure to either (A) 1 mM or (B) 2 mM H₂O₂. Intracellular ROS levels were measured using DCFH-DA fluorescence intensity, expressed in Relative Fluorescence Units (RFU). Data are presented as mean ± SEM (n = 4). Statistical significance compared to the untreated negative control group (the far-right bar) was determined using One-way ANOVA followed by Dunnett’s post-hoc test: ***p < 0.0001, ***p < 0.001, **p < 0.01, *p < 0.1, ns = not significant.
The study investigated the potential of BFBE to mitigate H₂O₂ induced ROS generation in RAW 264.7 cells. The results demonstrate that the extract exhibited a significant, dose-dependent reduction in H₂O₂-induced reactive oxygen species production compared to the control group. The antioxidant and free radical scavenging capabilities of plant-derived compounds have been extensively explored for their potential applications in various health conditions [1,2,28]. In the present study, RAW 264.7 cells were treated with BFBE at different concentrations and the intracellular ROS levels were assessed using the DCFH-DA assay.
Figure 3 presents evidence of BFBEʼs ability to protect RAW 264.7 cells from H₂O₂-induced ROS generation. When cells were pretreated with BFBE and then exposed to 1 mM H₂O₂ (Figure 3(A)), a clear dose-dependent reduction in ROS production was observed. Concentrations of 50 µg/mL (p < 0.01), 25 µg/mL (p < 0.001), and 12.5 µg/mL (p < 0.0001) BFBE all significantly lowered ROS levels compared to untreated cells. While 100 µg/mL BFBE and α-tocopherol also showed a protective trend, their effects were less pronounced (p < 0.1). However, under higher ROS generation (2 mM H₂O₂, Figure 3B), only 6.25 µg/mL BFBE significantly reduced ROS (p < 0.01), while the higher concentrations did not differ significantly from untreated cells. Both α-tocopherol and the positive control (H₂O₂ alone) consistently showed significantly elevated ROS levels (p < 0.0001) compared to untreated cells in both experimental conditions.
The results suggest that BFBE may possess potent antioxidant properties and can effectively mitigate ROS generation induced by H₂O₂. This indicates the potential of the extract to be further investigated for its therapeutic applications in oxidative stress-related disorders [1,29].
Antioxidant enzyme assay
The effects of BFBE on antioxidant enzyme activity were evaluated using commercially available kits from Cayman Chemical Company. Interestingly, while the positive controls for each assay confirmed their validity, BFBE exhibited a mixed effect on the enzymes tested.
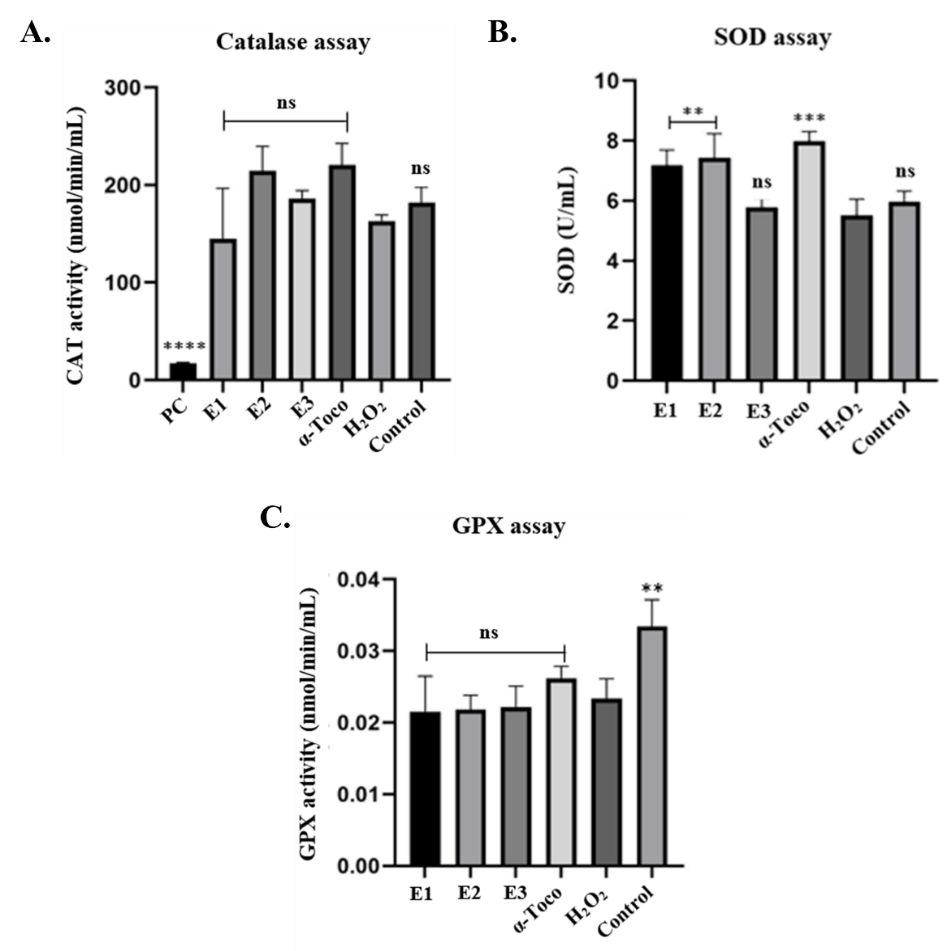
Figure 4 Effect of Baeckea frutescens branches ethanolic extract BFBE on antioxidant enzyme activities in H₂O₂-challenged RAW 264.7 cells. Cells were pretreated with BFBE (E1: 50 µg/mL, E2: 25 µg/mL, E3: 12.5 µg/mL) or α-tocopherol (α-Toco) for 24 h before exposure to 1 mM H₂O₂ for 30 min. (A) Catalase (CAT) activity, (B) Superoxide dismutase (SOD) activity, and (C) Glutathione peroxidase (GPX) activity. Data are presented as mean ± SEM (n = 4). Significant differences compared to the H₂O₂-treated group were determined using one-way ANOVA followed by Dunnettʼs post-hoc test (p < 0.01, ***p < 0.001, **p < 0.0001, ns = non-significant).
Specifically, BFBE (at concentrations of 50, 25, and 12.5 µg/mL) and α-tocopherol did not significantly alter catalase activity compared to the H₂O₂ control group. However, pre-treatment with the two higher concentrations of BFBE (50 and 25 µg/mL) significantly mitigated the H₂O₂-induced decrease in superoxide dismutase activity. α-Tocopherol also showed a significant protective effect on SOD. In contrast, none of the treatments, including BFBE or α-tocopherol, significantly impacted glutathione peroxidase activity, compared to the H₂O₂ control. These results suggest a potential selective action of BFBE on SOD activity, warranting further investigation [1,29].
Discussion
The maceration method effectively yielded 8.84% crude ethanolic extract from the branches of Baeckea frutescens. The extract was found to be exceptionally rich in bioactive compounds, notably saponins and hydrolysable tannins. These phytochemicals are widely recognized for their robust anti-inflammatory, antioxidant, antimicrobial, and antiviral properties [7,26,27]. The extract also contained moderate levels of flavonoids, a class of polyphenolic compounds recognized for their antioxidant and anti-inflammatory activities [2,30]. While steroids were less abundant, the absence of alkaloids and triterpenoids provides valuable insight into the extractʼs overall chemical makeup, guiding future research and potential applications [22,31].
Saponins are secondary plant compounds that are structurally diverse, comprising triterpene and steroid glycosides. Beyond their role in defending plants against pathogens and herbivores, saponins have been found to provide numerous health benefits, including anti-coagulant, anti-inflammatory, anti-carcinogenic, antioxidant, and neuroprotective properties [32].
Tannins represent another significant group of plant secondary metabolites that have demonstrated promising antimicrobial, antioxidant, and anti-inflammatory properties [4]. These polyphenolic compounds are naturally present in plants, particularly in the bark and wood. Tannins function as a defense mechanism against environmental stressors such as light, insects, fungi, and bacteria [33]. Hydrolysable tannins are composed of simple phenols that can be broken down by weak acids or bases into phenolic acids and carbohydrates [3]. The tanning industry has utilized tannins that are reported to possess medicinal attributes, including antiviral, antimicrobial, anti-inflammatory, and anti-tumor effects [3,33].
Flavonoids are low-molecular-weight polyphenolic secondary metabolites widely distributed throughout the plant kingdom [34]. Research has demonstrated that these compounds play a vital role in plant survival, functioning in signaling processes, flower pigmentation, auxin transport, and allelopathy [30]. Furthermore, flavonoids have been shown to exhibit health-promoting effects in various pharmaceutical, medicinal, nutraceutical, and cosmetic applications [4,28]. Numerous studies have proven that flavonoids possess anti-inflammatory, antioxidant, anti-mutagenic, and anti-carcinogenic properties [3].
Steroids are commonly used as anti-inflammatory drugs to treat various medical conditions [35]. They are also essential for plant growth and development, and may help regulate animal cellular processes [36]. Steroids found in plants possess a range of agrochemical, medicinal and pharmaceutical properties, including immunosuppressive, antibacterial, anthelminthic and anti-inflammatory effects [37].
The phytochemical compounds present in plants, such as glycosides, saponins, steroids, tannins, and flavonoids, contribute significantly to the medicinal and pharmaceutical industries [37]. In light of the growing concern over the emergence of antibiotic-resistant pathogens, there is an increasing emphasis on exploring alternative therapeutic approaches, including the use of plant-derived bioactive compounds [4,25].
The antioxidant properties of Baeckea frutescens L. branches ethanolic extract (BFBE) were comprehensively evaluated through DPPH assays and FRAP assay [3,26]. The DPPH radical scavenging assay demonstrated a clear concentration-dependent relationship, where higher concentrations of the extract exhibited greater free radical scavenging ability [24,38]. The highest tested concentration of 1,000 μg/mL showed a remarkably potent 91.24% radical scavenging activity, corroborated by the extract’s median inhibitory concentration of 86.16 μg/mL, indicating its highly efficient free radical quenching properties. This suggests that the extract possesses a strong capacity to neutralize free radicals, which are known contributors to oxidative stress and cellular damage.
The ferric reducing antioxidant power assay further revealed the extract’s robust reducing antioxidant capacity, as it could reduce ferric ions (Fe3+) to the ferrous form (Fe2+) in a concentration-dependent manner [18,32]. The highest tested concentration of 1,000 μg/mL exhibited a gallic acid equivalent of 31.91 μM, suggesting the extractʼs overall potent antioxidant potential. The ability to reduce ferric ions is an important indicator of a compound’s capacity to prevent oxidative reactions and protect against oxidative stress.
These findings indicate that the BFBE possesses strong antioxidant capacities, which may be attributed to its phytochemical composition, consistent with reports on the antioxidant properties of other plant extracts [14,19]. The extractʼs concentration-dependent radical scavenging and ferric ion reducing abilities suggest it as a promising natural source of antioxidants, likely due to the presence of bioactive compounds such as phenolics and flavonoids. Further research to identify and quantify the specific antioxidant compounds in the extract would provide deeper insights into its potential applications and mechanisms of action.
This study utilized the MTT assay to assess the cytotoxic effects of BFBE and H₂O₂ on RAW 264.7 murine macrophages [18,39]. The MTT assay, a colorimetric method, measures cell viability by assessing metabolic activity. Viable cells with active mitochondria reduce the yellow tetrazolium salt MTT to purple formazan crystals, which are then solubilized and quantified spectrophotometrically. The absorbance readings directly correlate with the number of viable cells [18,31,40]. Therefore, a reduction in absorbance indicates a decrease in cell viability, suggesting a cytotoxic effect. The results demonstrated that the BFBE exhibited a concentration-dependent effect on RAW 264.7 cell viability [41,42].
The results align with the objectives of using the MTT assay by demonstrating concentration-dependent cytotoxicity for all three compounds. The BFBE showed significant reductions in cell viability at higher concentrations, indicating a cytotoxic effect at these levels. The lack of significant effect at lower concentrations suggests a potential threshold for cytotoxicity [18,31,43,44].
Similarly, the concentration-dependent effect of H₂O₂, show significant effects at 4 mM, further validates the assayʼs ability to detect cytotoxic effects across a range of concentrations. The use of lower, non-cytotoxic concentrations of H₂O₂ in subsequent antioxidant experiments, respectively, further demonstrates the utility of the MTT assay in determining appropriate concentrations for follow-up studies. [18,19,42,45].
Next, the antioxidant potential of BFBE in RAW 264.7 cells exposed to H₂O₂-induced ROS generation was evaluated using 2 complementary approaches. Firstly, a DCFH-DA assay demonstrated a significant, dose-dependent reduction in intracellular ROS levels upon BFBE pre-treatment, particularly at concentrations between 12.5 and 50 µg/mL [29,41]. This finding aligns with previous research highlighting the potent free radical scavenging capabilities of plant-derived compounds, including those from Baeckea frutescens [14,19,26]. This direct ROS neutralization capacity is likely a significant contributor to BFBEʼs protective effects against oxidative damage. Secondly, antioxidant enzyme activity assays were performed to evaluate whether BFBE modulates endogenous cellular antioxidant defenses. The findings revealed a nuanced effect of BFBE on antioxidant enzyme activity in RAW 264.7 cells under H₂O₂-induced ROS generation. BFBE exhibited a selective modulation of antioxidant enzymes, primarily affecting superoxide dismutase while leaving catalase and glutathione peroxidase activity largely unchanged.
Specifically, BFBE, at concentrations of 50 and 25 µg/mL, significantly protected SOD activity against H₂O₂-induced suppression. This suggests that BFBE may contain compounds that either directly protect SOD from oxidative damage or enhance its activity under stress conditions [42]. The preservation of SOD activity compared with CAT and GPx observed in this study may be related to the phytochemical composition of BFBE, which likely contains flavonoids or polyphenolic compounds with antioxidant properties and potential superoxide radical scavenging activity. SOD plays an important role as the first line of defense in the cellular antioxidant system by catalyzing the conversion of superoxide radicals, a primary reactive oxygen species involved in oxidative damage, into hydrogen peroxide (H₂O₂) [46]. Because superoxide radicals can also inactivate the SOD enzyme through oxidative modification, it is possible that BFBE may reduce superoxide accumulation and thereby help preserve SOD activity under oxidative stress conditions. While H₂O₂ can contribute to oxidative damage, it also functions as a signaling molecule involved in various cellular processes [47-49]. Therefore, maintaining SOD activity may contribute to regulating the balance between ROS detoxification and redox signaling.
The lack of significant changes in catalase and GPx activity suggests that BFBE did not noticeably influence these antioxidant enzymes under the experimental conditions tested. Catalase is responsible for decomposing H₂O₂ into water and oxygen, whereas GPx reduces lipid hydroperoxides and other organic peroxides. The absence of significant modulation of these enzymes may indicate that BFBE exerted its antioxidant effects primarily through ROS scavenging rather than through broad modulation of multiple antioxidant enzyme systems. However, since the present study only assessed enzyme activity, further investigations examining gene and protein expression levels are required to determine whether BFBE directly regulates antioxidant enzymes at the transcriptional or translational level.
It is also noteworthy that α-tocopherol, a well-known antioxidant, showed a similar protective trend for SOD activity but did not significantly alter catalase or GPx activity [1,40]. This observation suggests that certain antioxidants, including BFBE, may preferentially support the superoxide dismutation pathway under oxidative stress conditions. Nevertheless, additional studies are required to clarify the molecular mechanisms underlying these effects. Investigating whether this response involves pathways such as Nrf2-mediated antioxidant regulation would be valuable [50]. Future studies using RT-qPCR or protein expression analysis could further determine whether these effects are associated with transcriptional or post-translational regulation of antioxidant enzymes.
Overall, the results of this study provide compelling evidence for the antioxidant activity of BFBE in RAW 264.7 cells, highlighting a dual-action mechanism involving both direct ROS scavenging and selective antioxidant enzyme modulation [18,29]. On the other hand, further research is warranted to fully elucidate the specific bioactive compounds responsible for these effects, explore potential modulation of antioxidant enzyme gene expression, and investigate the involvement of BFBE in cellular signaling pathways related to oxidative stress responses [4,25].
Conclusions
This study highlights the phytochemical richness of the ethanolic extract of Baeckea frutescens branches, which is abundant in bioactive compounds such as saponins, tannins, and flavonoids. The extract demonstrated potent antioxidant activity and significantly reduced intracellular ROS levels. These findings suggest that Baeckea frutescens extract holds promise as a natural therapeutic agent for managing elevated ROS generation and oxidative stress, warranting further in vivo studies and clinical investigations.
Acknowledgements
This research was financially supported by Universiti Teknologi MARA (UiTM), Malaysia under Internal Research Grant Universiti Teknologi MARA Selangor (DUCS-A) RMI File No: 600-UiTMSEL (PI. 5/4/8) (014/2025) and Institute of Postgraduate Studies UiTM.
Declaration of Generative AI in Scientific Writing
During the preparation of this work, we utilized Jenni AI during the preparation of this manuscript to generate ideas and paraphrase text. Following the use of this tool, the author(s) carefully reviewed and edited the content and took full responsibility for the final publication.
CRediT Author Statement
Nurhanani Ayub: Writing - Original Draft, Methodology, Validation, Formal analysis, Investigation, Data Curation. Salfarina Ramli: Writing - Review and Editing, Conceptualization, Methodology, Validation. Ihsan Safwan Kamarazaman: Writing - Review and Editing, Conceptualization, Methodology, Validation. Evana Kamarudin: Writing - Review and Editing, Conceptualization, Methodology, Validation. Richard Johari James: Writing - Review and Editing, Conceptualization, Methodology, Validation. Nurul Badriah: Writing - Review and Editing, Conceptualization, Methodology, Validation. Hasseri Halim: Project administration, Project acquisition, Formal analysis, Writing - Review and Editing, Conceptualization, Methodology, Validation, Resources, Data Curation, Supervision.
References
[1] AM Pisoschi and A Pop. The role of antioxidants in the chemistry of oxidative stress: A review. European Journal of Medicinal Chemistry 2015; 97, 55-74.
[2] P Arulselvan, MT Fard, WS Tan, S Gothai, S Fakurazi, ME Norhaizan and SS Kumar. Role of antioxidants and natural products in inflammation. Oxidative Medicine and Cellular Longevity 2016; 2016, 5276130.
[3] A Altemimi, N Lakhssassi, A Baharlouei, DG Watson and DA Lightfoot. Phytochemicals: Extraction, isolation, and identification of bioactive compounds from plant extracts. Plants 2017; 6, 42.
[4] M Samtiya, RE Aluko, T Dhewa and JM Moreno-Rojas. Potential health benefits of plant food-derived bioactive components: An overview. Food 2021;10(4), 839.
[5] FFM Ariff, SK Muniandi, N Saffie, SNS Bahari, Z Abdullah, LS Kiong, MM Taini, SNAM Hashim, F Jamaludin and MZ Abdullah. Genotype screening and propagation techniques of three selected medicinal plant species for production of high-quality planting material. Horticulturae 2021; 7(12), 570.
[6] DTL Huong, DX Duc and NT Son. Baeckea frutescens L.: A review on phytochemistry, biosynthesis, synthesis, and pharmacology. Natural Product Communications 2023; 18(7), 1-22.
[7] M Herb and M Schramm. Functions of ROS in macrophages and antimicrobial immunity. Antioxidants 2021; 10(2), 313.
[8] M Mittal, MR Siddiqui, K Tran, SP Reddy and AB Malik. Reactive oxygen species in inflammation and tissue injury. Antioxidants & Redox Signaling 2014; 20(7), 1126-1167.
[9] S Pérez and S Rius-Pérez. Macrophage polarization and reprogramming in acute inflammation: A redox perspective. Antioxidants 2022; 11(7), 1394.
[10] SH Baek, T Park, MG Kang and D Park. Anti-inflammatory activity and ROS regulation effect of sinapaldehyde in LPS-stimulated RAW 264.7 macrophages. Molecules 2020; 25, 4089.
[11] L Stanca, OI Geicu, AI Serban and A Dinischiotu. Interplay of oxidative stress, inflammation, and autophagy in RAW 264.7 murine macrophage cell line challenged with Si/SiO2 quantum dots. Materials 2023; 16(14), 5083.
[12] O Mohanta, A Ray, S Jena, A Sahoo, SS Panda, PK Das, S Nayak and PC Panda. Mesosphaerum suaveolens essential oil attenuates inflammatory response and oxidative stress in LPS-stimulated RAW 264.7 macrophages by regulating NF-κB signaling pathway. Molecules 2023; 28(15), 5817.
[13] S Yoojam, A Ontawong, N Lailerd, K Mengamphan and D Amornlerdpison. The enhancing immune response and anti-inflammatory effects of Caulerpa lentillifera extract in RAW 264.7 cells. Molecules 2021; 26(19), 5734.
[14] K Nisa, S Nurhayati, W Apriyana and AW Indrianingsih. Investigation of total phenolic and flavonoid contents, and evaluation of antimicrobial and antioxidant activities from Baeckea frutescens extracts. IOP Conference Series: Earth and Environmental Science 2017; 101, 012002.
[15] Q Zeng, H Xie, H Song, F Nie, J Wang, D Chen and F Wang. In vivo wound healing activity of Abrus cantoniensis extract. Evidence-Based Complementary and Alternative Medicine 2016; 2016, 6568528.
[16] E Iqbal, K Abu and LBL Lim. Phytochemical screening, total phenolics and antioxidant activities of bark and leaf extracts of Goniothalamus velutinus (Airy Shaw) from Brunei Darussalam. Journal of King Saud University - Science 2015; 27(3), 224-232.
[17] MJ Kim, SM Jeong, BK Kang, KBWR Kim and DH Ahn. Anti-inflammatory effects of grasshopper ketone from Sargassum fulvellum ethanol extract on lipopolysaccharide-induced inflammatory responses in RAW 264.7 cells. Journal of Microbiology and Biotechnology 2019; 29(5), 820-826.
[18] KA Hwang, YJ Hwang and J Song. Antioxidant activities and oxidative stress inhibitory effects of ethanol extracts from Cornus officinalis on RAW 264.7 cells. BMC Complementary and Alternative Medicine 2016; 16, 196.
[19] S Navanesan, NA Wahab, S Manickam and KS Sim. Evaluation of selected biological capacities of Baeckea frutescens. BMC Complementary and Alternative Medicine 2015; 15, 186.
[20] PTH Ha, TK Cuong, NTT Nha, NPH Lan, NHT Anh, BL Giang, PM Quan, TQ Toan, PQ Long and NT Luan. A study on biological activities of Bouea macrophylla Griff leaf extract. Vietnam Journal of Biotechnology 2022; 20(2), 339-349.
[21] MP Junqueira-Gonçalves, L Yáñez, C Morales, M Navarro, RA Contreras and GE Zúñiga. Isolation and characterization of phenolic compounds and anthocyanins from murta (Ugni molinae Turcz.) fruits. Assessment of antioxidant and antibacterial activity. Molecules 2015; 20(4), 5698-5713.
[22] L Stanojević, M Stanković, V Nikolić, L Nikolić, D Ristić, J Čanadanovic-Brunet and V Tumbas. Antioxidant activity and total phenolic and flavonoid contents of Hieracium pilosella L. extracts. Sensors 2009; 9(7), 5702-5714.
[23] MM Raimi and OO Oyedapo. Bioactivity-guided evaluation of the root extract of Plumbago zeylanica. International Journal of Biological and Chemical Sciences 2009; 3(4), 672-680.
[24] J Li, Y Qin, X Yu, Z Xiong, L Zheng, Y Sun, J Shen, N Guo, L Tao, Z Deng and X Liu. Trends in phytochemical research. Journal of Food Biochemistry 2019; 43(6), 12947.
[25] DM Kasote, SS Katyare, MV Hegde and H Bae. Significance of antioxidant potential of plants and its relevance to therapeutic applications. International Journal of Biological Sciences 2015; 11(8), 982-991.
[26] S Razmavar, MA Abdulla, SB Ismail and P Hassandarvish. Antibacterial activity of leaf extracts of Baeckea frutescens against methicillin-resistant Staphylococcus aureus. BioMed Research International 2014; 2014, 521287.
[27] SH Shahruzaman, MF Mustafa, S Ramli, S Maniam, S Fakurazi and S Maniam. The cytotoxic effect and glucose uptake modulation of Baeckea frutescens on breast cancer cells. BMC Complementary and Alternative Medicine 2019; 19, 220.
[28] K Neha, MR Haider, A Pathak and MS Yar. Medicinal prospects of antioxidants: A review. European Journal of Medicinal Chemistry 2019; 178, 687-704.
[29] M Gasparrini, TY Forbes-Hernandez, F Giampieri, S Afrin, JM Alvarez-Suarez, L Mazzoni, B Mezzetti, JL Quiles and M Battino. Anti-inflammatory effect of strawberry extract against LPS-induced stress in RAW 264.7 macrophages. Food and Chemical Toxicology 2017; 102, 1-10.
[30] M Khalid, Saeed-Ur-Rahman, M Bilal and D Huang. Role of flavonoids in plant interactions with the environment and against human pathogens - a review. Journal of Integrative Agriculture 2019; 18(1), 211-230.
[31] SA Adebayo, JP Dzoyem, LJ Shai and JN Eloff. The anti-inflammatory and antioxidant activity of 25 plant species used traditionally to treat pain in southern African. BMC Complementary and Alternative Medicine 2015; 15, 159.
[32] S Addisu and A Assefa. Role of plant containing saponin on livestock production; A review. Advances in Biological Research 2016; 10(5), 309-314.
[33] Y Shirmohammadli, D Efhamisisi and A Pizzi. Tannins as a sustainable raw material for green chemistry: A review. Industrial Crops and Products 2018; 126, 316-332.
[34] AN Panche, AD Diwan and SR Chandra. Flavonoids: An overview. Journal of Nutritional Science 2016; 5(47), 1-15.
[35] AB Shinde and YR Mulay. Phytochemical analysis and antibacterial properties of some selected Indian medicinal plants. International Journal of Current Microbiology and Applied Sciences 2015; 4(3), 228-235.
[36] HE Tamez-Pérez, DL Quintanilla-Flores, R Rodríguez-Gutiérrez, JG González-González and AL Tamez-Peña. Steroid hyperglycemia: Prevalence, early detection and therapeutic recommendations: A narrative review. World Journal of Diabetes 2015; 6(8), 1073-1081.
[37] SS Patel and JK Savjani. Systematic review of plant steroids as potential antiinflammatory agents: Current status and future perspectives. The Journal of Phytopharmacology 2015; 4(2), 121-125.
[38] EO Sousa, CMBA Miranda, CB Nobre, AA Boligon, ML Athayde and JGM Costa. Phytochemical analysis and antioxidant activities of Lantana camara and Lantana montevidensis extracts. Industrial Crops and Products 2015; 70, 7-15.
[39] T Poongodi, TH Nazeema and B Ranjini. Enhanced anti-inflammatory effect of polyherbal formulation (MKA) comprising of three selected plants in lipopolysaccharide (LPS)-induced RAW 264.7 macrophage cell line. Indian Journal of Pharmaceutical Sciences 2020; 82(4), 692-697.
[40] L Jamalzadeh, H Ghafoori, R Sariri, H Rabuti, J Nasirzade, H Hasani and MR Aghamaali. Cytotoxic effects of some common organic solvents on MCF-7, RAW-264.7 and human umbilical vein endothelial cells. Avicenna Journal of Medical Biochemistry 2016; 4(1), 33458.
[41] Z Lin, S Ku, T Lim, SY Park, MS Park, GE Ji, K O’Brien and KT Hwang. Antioxidant and anti-inflammatory properties of recombinant Bifidobacterium bifidum BGN4 expressing antioxidant enzymes. Microbiology 2021; 93(3), 595.
[42] S Chandra, S Khan, B Avula, H Lata, MH Yang, MA ElSohly and IA Khan. Assessment of total phenolic and flavonoid content, antioxidant properties, and yield of aeroponically and conventionally grown leafy vegetables and fruit crops: A comparative study. Evidence-Based Complementary and Alternative Medicine 2014; 2014, 253875.
[43] APA de Carvalho and CA Conte-Junior. Health benefits of phytochemicals from Brazilian native foods and plants: Antioxidant, antimicrobial, anti-cancer, and risk factors of metabolic/endocrine disorders control. Trends in Food Science & Technology 2021; 111, 534-548.
[44] N Jaradat, M Hawash and G Dass. Phytochemical analysis, in-vitro anti-proliferative, anti-oxidant, anti-diabetic, and anti-obesity activities of Rumex rothschildianus Aarons. extracts. BMC Complementary Medicine and Therapies 2021; 21, 107.
[45] D Figueroa, M Asaduzzaman and F Young. Real time monitoring and quantification of reactive oxygen species in breast cancer cell line MCF-7 by 2′,7′–dichlorofluorescin diacetate (DCFDA) assay. Journal of Pharmacological and Toxicological Methods 2018; 94(1), 26-33.
[46] OM Ighodaro and OA Akinloye. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alexandria Journal of Medicine 2018; 54(4), 287-293.
[47] M Schieber and NS Chandel. ROS function in redox signaling and oxidative stress. Current Biology 2014; 24(10), 453-462.
[48] P Sharma, AB Jha, RS Dubey and M Pessarakli. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. Journal of Botany 2012; 2012, 217037.
[49] TAF Aguilar, BCH Navarro and JAM Pérez. Endogenous antioxidants: A review of their role in oxidative stress. IntechOpen Limited, London, 2016, p. 3-20.
[50] Q Ma. Role of nrf2 in oxidative stress and toxicity. Annual Review of Pharmacology and Toxicology 2013; 53, 401-426.