Trends
Sci.
2026;
23(10):
13267
Neuroprotective Potential of p-MCA in Drosophila
Nur Rahma Rumata1,2, Asbah Asbah3,8, Nadila Pratiwi Latada8, Muhammad Arfandy Gunawan4, Fifi Dismayanti Indriani Nainu5, Irmanida Batubara6, Muhammad Aswad7,
Berry Juliandi1,* and Firzan Nainu3,8
1Department of Biology, Faculty of Mathematics and Natural Sciences, IPB University, Bogor 16680, Indonesia
2Faculty of Health Sciences, Almarisah Madani University, Makassar 90245, Indonesia
3Department of Pharmacy, Faculty of Pharmacy, Hasanuddin University, Makassar 90245, Indonesia
4Postgraduate Program in Faculty of Pharmacy, Hasanuddin University, Makassar 90245, Indonesia
5Postgraduate Program in Biomedical Sciences, Postgraduate Program, Hasanuddin University, Makassar 90245, Indonesia
6Department of Chemistry, Faculty of Mathematics and Natural Sciences, Tropical Biopharmaca Research Center - IPB University, Bogor 16680, Indonesia
7Faculty of Pharmacy, Hasanuddin University, Makassar 90245, Indonesia
8Unhas Fly Research Group, Faculty of Pharmacy, Hasanuddin University, Makassar 90245, Indonesia
(*Corresponding author’s e-mail: [email protected])
Received: 5 January 2026, Revised: 21 February 2026, Accepted: 1 March 2026, Published: 10 May 2026
Abstract
p-methoxycinnamic acid (p-MCA), a bioactive compound from Kaempferia galanga, was evaluated for its neuroprotective effects in an Alzheimer’s disease (AD)-like model of Drosophila melanogaster, supported by in silico docking and molecular dynamics simulations. Toxicity screening identified 10 mM AlCl3 (aluminium chloride) as an effective concentration to induce neurodegenerative phenotypes without excessive mortality. p-MCA (0.01-0.1 mM) significantly improved survival, locomotor performance, and phototactic responses in AlCl3-exposed flies. Gene expression analysis showed upregulation of antioxidant (sod1, hsp22) and anti-aging (srl) genes, along with modulation of the apoptosis-related gene grim, indicating regulation of oxidative stress and cellular homeostasis. Computational analysis demonstrated stable binding of p-MCA to acetylcholinesterase (ΔG = ‒7.2 kcal/mol), comparable to 10-40. Collectively, these findings indicate that p-MCA mitigates AlCl3-induced neurotoxicity through multi-target mechanisms and supports its further evaluation in neurodegenerative disease models.
Keywords: p-MCA, Alzheimer’s disease, Neuroprotection, D. melanogaster, Oxidative stress, Drug discovery, Neurodegenerative disorders
Introduction
Aging is associated with progressive functional decline driven by oxidative stress, genomic instability, and cellular damage, increasing the risk of neurodegenerative diseases such as Alzheimer’s disease (AD) [1,2]. AD is characterized by central nervous system degeneration, amyloid beta (Aβ) plaque deposition, and neurofibrillary tangles, leading to synaptic dysfunction and cognitive decline [3-7]. As the most prevalent age-related neurodegenerative disorder, AD represents a major global health concern, particularly in aging populations, including Indonesia, where cases are projected to rise substantially by 2030 [8].
Current pharmacological treatments, such as N-methyl-D-aspartate receptor antagonists (memantine) and cholinesterase inhibitors, provide only symptomatic relief without preventing disease progression [3,9-12]. These limitations highlight the need for multi-target therapeutic strategies addressing the molecular mechanisms underlying AD pathology [4,11].
Natural compounds have gained attention as potential neuroprotective agents. p-MCA derived from Kaempferia galanga exhibits antimicrobial, anticancer, anti-inflammatory, and anticholinesterase activities [12,13]. Experimental evidence indicates that p-MCA protects neurons from excitotoxicity and attenuates memory impairment in animal models, suggesting neuroprotective potential. However, its underlying molecular and genetic mechanisms remain insufficiently characterized.
Drosophila melanogaster provides a cost-effective and genetically tractable in vivo model for aging and neurodegeneration research [14-16], sharing substantial genetic conservation with humans and supporting transgenic approaches for mechanistic studies and therapeutic screening [16-18]. Therefore, this study aims to evaluate the neuroprotective effects of p-MCA using D. melanogaster as an AD-like model, providing mechanistic insights relevant to the development of novel therapeutic strategies for neurodegenerative diseases.
Materials and methods
This preclinical experimental study used a post-test only control design to evaluate the neuroprotective effects of p-MCA against AlCl3-induced AD-like pathology in D. melanogaster (w1118). Behavioral assays (toxicity, survival, locomotor activity, and phototaxis), gene expression analysis, and in silico molecular studies were conducted.
D. melanogaster stock
Adult male and female D. melanogaster (w1118) were used in this study. The fly line was obtained from the Laboratory of Host Defense and Responses, Kanazawa University, Japan, and maintained for more than 20 generations in the Laboratory of Pharmacology and Toxicology, Faculty of Pharmacy, Universitas Hasanuddin, Indonesia. Flies were reared on a standard cornmeal-based medium at 25 °C under a 12-hour light/dark cycle [19].
Sample preparation
p-MCA, obtained from Tokyo Chemical Industry Co. Ltd®, was dissolved in analytical-grade dimethyl sulfoxide (DMSO).
Toxicity assay
To determine a non-lethal concentration of AlCl3 for AD-like induction, a toxicity assay was performed. w1118 flies (3-5 day-old, 10 flies per vial) were exposed to AlCl3 at concentrations of 10, 20, 30, and 40 mM for 7 days, with three replicates per group. Flies were maintained at 25 °C, and survival was recorded daily. AlCl3 was dissolved in distilled water and administered via the fly medium.
Survival assay
Survival analysis was conducted using 3-5 day-old w1118 flies (5 males and 5 females per vial). Flies were treated with p-MCA at concentrations of 1, 0.1, 0.01, or 0.001 mM in combination with AlCl3, while the control group received standard diet. Food was replaced every two days, and survival was recorded daily for 7 days. Survival data were analyzed using Kaplan-Meier curves with log-rank tests in GraphPad Prism® 8. An initial dose-range screening (1, 0.1, 0.01, and 0.001 mM) was performed in survival and locomotor assays to assess the toxicity and biological responsiveness of p-MCA under AlCl3 exposure. Concentrations showing non-lethal profiles and measurable protective effects were subsequently selected for downstream molecular and behavioral analyses.
Negative geotaxis and phototaxis assays
Locomotor and sensory-cognitive functions were assessed using negative geotaxis and phototaxis assays, respectively. Negative geotaxis was performed with minor modifications of a previously described protocol [26], in which treated flies (3-5 day-old) were transferred into vertically oriented vials marked with a reference line, tapped to the bottom, and the number of flies crossing the line within 15 seconds was recorded. Flies treated with AlCl3 (10 mM) and p-MCA for 7 days were analyzed. Phototactic behavior was evaluated as an indicator of sensory and cognitive function [20,21]. For this assay, 0.01 mM was selected based on preliminary screening demonstrating consistent efficacy without toxicity. Thirty age-matched flies per group (30 males and 30 females analyzed separately) were randomly selected and tested. After 30 min of dark adaptation, flies were placed in a horizontally positioned phototaxis apparatus perpendicular to a light source (15 cm distance). The apparatus was divided into four sections, and fly distribution was recorded at 1-min intervals. Each assay included three consecutive technical trials using the same flies and vial on the same day, and the mean value was used for statistical analysis. Control measurements were performed using an identical apparatus aligned parallel to the light source. Results were expressed as the percentage of flies in the light and dark compartments.
Gene expression assay
Based on the survival screening results, 1, 0.1, and 0.01 mM were selected for gene expression analysis. The highest concentration (1 mM) was included to evaluate potential upper-range molecular responses, while 0.1 and 0.01 mM were selected due to their survival-prolonging effects. The lowest concentration (0.001 mM) was excluded from further mechanistic assays due to the absence of detectable biological effects in the initial screening. Total RNA was isolated from five live flies per experimental group using the Monarch Total RNA Miniprep Kit (New England Biolabs, Inc., MA). RNA concentration was measured using a NanoDrop spectrophotometer (BioDrop, USA). Expression levels of sod1, hsp22, rpr, grim, srl, and AChE were quantified by RT-qPCR using Luna® reagents (New England Biolabs, Inc., MA, USA) in a 10 µL reaction volume. The rp49 gene was used as an internal reference. Amplification was performed using a Rotor-Gene Q thermal cycler (Qiagen, Germany) under standard cycling conditions, followed by melt curve analysis. Relative gene expression was calculated using comparative quantification. Primer sequences are listed in Table 1 [22].
Table 1 Primers used in the RT-qPCR assay.
Genes |
Forward primer |
Reverse primer |
sod1 |
5’– AGG TCA ACA TCA CCG ACT CC – 3’ |
5’– GTT GAC TTG CTC AGC TCG TG – 3’ |
hsp22 |
5’ – TAC AAT GCG TTT CCT TAC CGA – 3’ |
5’ – CAA TCT GCT GCC AGT TCC T – 3’ |
grim |
5’ –TCG GAG TTT GGA TGC TGG GAT CTT– 3’ |
5’ – AGT CAC GTC GTC CTC ATC GTT GTT – 3’ |
srl |
5’ – CTC TTG GAG TCC GAG ATC CG CAA – 3’ |
5’ – GGG ACC GCG AGC TGA TGG TT – 3’ |
AChE |
5’ – GCC GTG GGC AAT GTA ATA GT – 3’ |
5’– CGA CTC TCC GAA CAG TGT CA – 3’ |
rp49 |
5’ – GAC GCT TCA AGG GAC AGT ATC TG – 3’ |
5’ – AAA CGC GGT TCT GCA TGA G – 3’ |
Molecular docking and molecular dynamics
The 3D structures of p-MCA (CID: 699414) and 10-40 (CID: 3152) were retrieved from PubChem and prepared using UCSF Chimera® [23,24]. Docking validation was carried out by re-docking the native ligand and calculating RMSD values using PyMOL, with interaction visualization performed in Discovery Studio® [24,25]. Molecular dynamics simulations were conducted using YASARA with the Amber14 force field. Simulations were run for 100 ns at 310 K and pH 7.4 under periodic boundary conditions. RMSD, RMSF, and radius of gyration were analyzed at 2.5 ps intervals [26].
Data processing
Survival data were analyzed using Kaplan-Meier curves and log-rank tests. Locomotor, phototaxis, and gene expression data were analyzed by one-way ANOVA followed by Tukey’s HSD post hoc test. Data are presented as mean ± SD, and p < 0.05 was considered statistically significant. Statistical analyses were performed using GraphPad Prism® 8.
Results and discussion
Dose-dependent toxicity of AlCl3 in D. melanogaster
This study aimed to determine the lowest effective dose of AlCl3 that induces Alzheimer’s-like symptoms without causing excessive toxicity. Exposure to AlCl3 for 7 days resulted in a concentration-dependent reduction in survival of D. melanogaster. Flies treated with 10-40 mM AlCl3 exhibited progressively decreased survival compared to the untreated control (Figure 1), indicating increasing toxicity at higher concentrations. Among the tested doses, 10 mM AlCl3-induced measurable neurodegenerative stress without causing excessive mortality and was therefore selected for subsequent experiments as an AD-like induction dose.
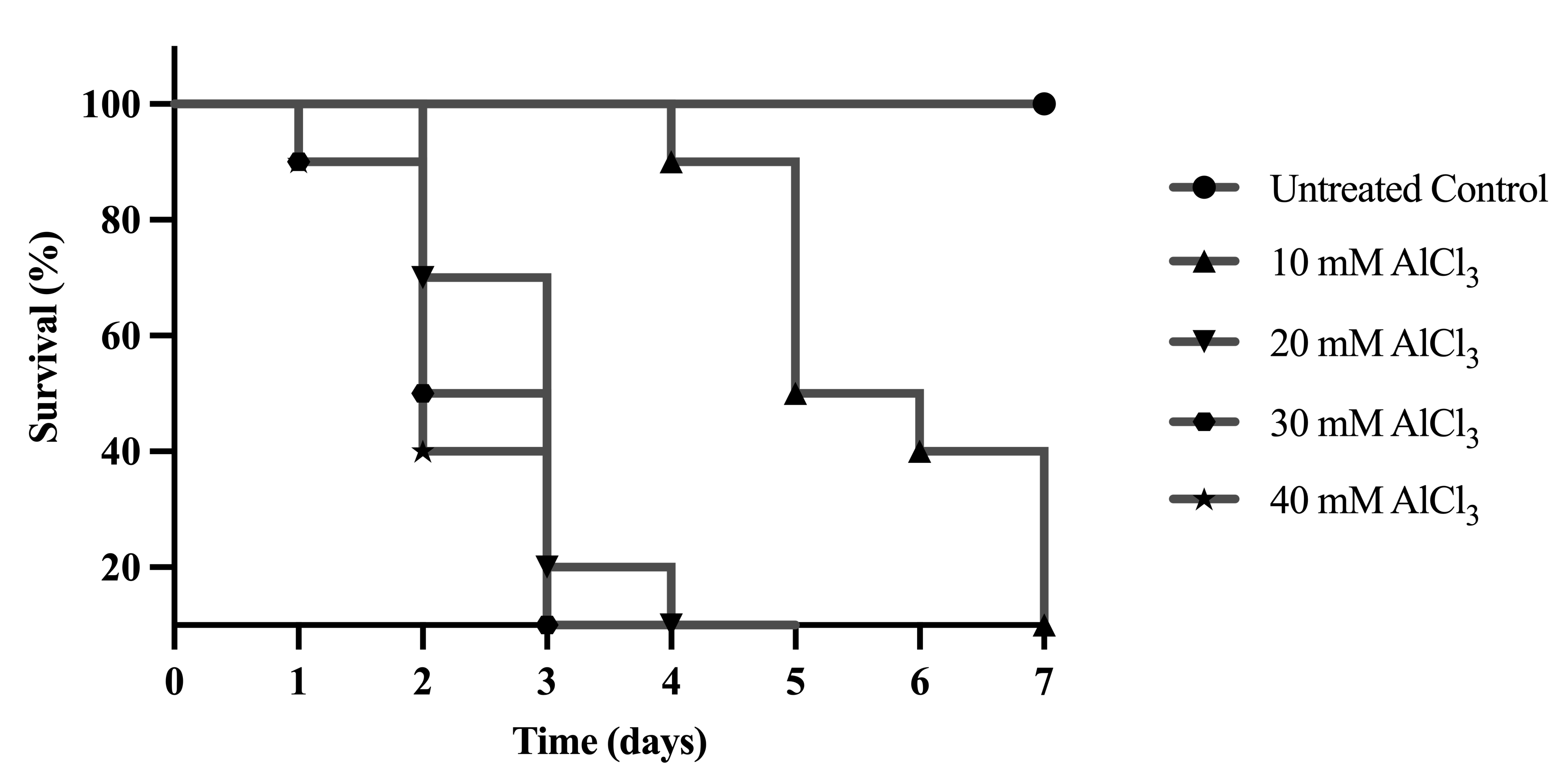
Figure 1 Dose-dependent toxicity of AlCl3 as an inducer for neurodegeneration in D. melanogaster. The survival of D. melanogaster declined upon exposure to different concentrations of AlCl3 (10-40 mM) compared to untreated control group. Since AlCl3 was dissolved in distilled water, the untreated control group served as the appropriate control condition.
p-MCA enhances survival in D. melanogaster exposed to AlCl3
This study investigated the neuroprotective effects of p-MCA by assessing its impact on the lifespan of D. melanogaster treated with AlCl3. Administration of p-MCA significantly improved survival in flies exposed to 10 mM AlCl3 (Figure 2). Notably, p-MCA at 0.1 mM and 0.01 mM produced the most pronounced effects, whereas higher and lower concentrations showed limited benefit. These findings indicate a concentration-dependent protective effect of p-MCA, consistent with its reported antioxidant and cytoprotective properties. Improved survival suggests that p-MCA enhances cellular resilience against AlCl3-induced toxicity, a key feature relevant to neurodegenerative pathology.

Figure 2 Improvement of D. melanogaster survival after administration of 10 mM AlCl3 and p-MCA at various concentrations (1 mM, 0.1 mM, 0.01 mM, and 0.001 mM). Flies were treated with 10 mM AlCl3 prior to treatment with p-MCA.
p-MCA enhances locomotor and cognitive function in D. melanogaster exposed to AlCl₃
Negative geotaxis analysis revealed that exposure to AlCl3 significantly impaired locomotor activity in adult D. melanogaster. In contrast, p-MCA treatment markedly improved climbing performance across the tested concentrations, indicating attenuation of AlCl3-induced motor dysfunction (Figure 3(A)).
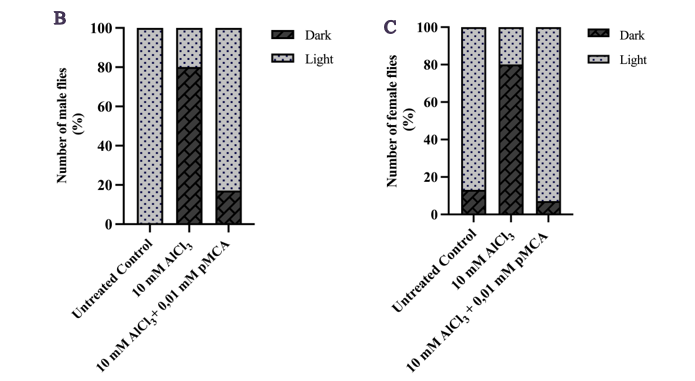
Phototaxis assays further demonstrated that p-MCA preserved light-directed behavior in both male and female flies. Male flies treated with p-MCA showed a significantly higher proportion migrating toward the light source compared to the AlCl3 only group (Figure 3(B)). A similar protective effect was observed in female flies, where p-MCA treatment restored phototactic responses disrupted by AlCl3 exposure (Figure 3(C)). Together, these behavioral outcomes support the neuroprotective role of p-MCA in maintaining motor coordination and sensory-cognitive function under neurotoxic stress.

![]()
Figure 3 Improvement of locomotor activity (A) and phototaxis response in male (B) and female (C) adult D. melanogaster w1118 following exposure to 10 mM AlCl3 and p-MCA at various concentrations (locomotor assay, A) and at 0.01 mM p-MCA (phototaxis assay, B - C). Data are expressed as mean ± SD. ***p < 0.001 and ****p < 0.0001 compared with the AlCl3-treated group.
p-MCA modulates gene expression of D. melanogaster exposed to AlCl3
RT-qPCR analysis demonstrated that p-MCA significantly modulated the expression of genes associated with antioxidant defense and stress response in AlCl3-exposed D. melanogaster. sod1 expression was significantly upregulated at 0.01 mM p-MCA compared with the AlCl3 group (Figure 4(A)). Conversely, hsp22 expression was consistently downregulated across all p-MCA concentrations (1, 0.1, and 0.01 mM) relative to AlCl3 exposure alone (Figure 4(B)).
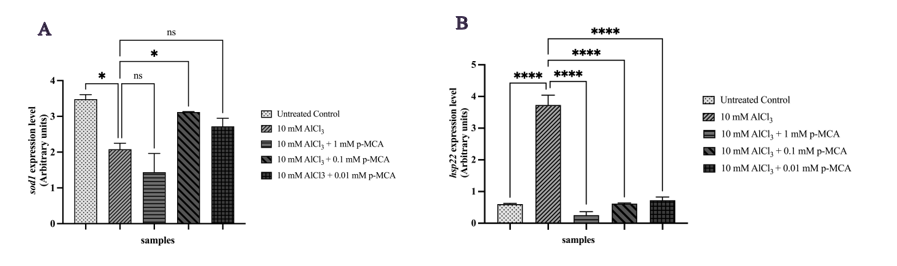
Figure 4 Relative expression levels of sod1 (A) and hsp22 (B) genes in adult D. melanogaster w1118 following exposure to 10 mM AlCl3 and p-MCA at different concentrations, as determined by RT-qPCR and normalized to rp49. Data are presented as mean ± SD. *p < 0.05 and ****p < 0.0001 compared with the AlCl3-treated group; ns, not significant.
Expression of the apoptosis-related gene grim was significantly increased at 0.01 mM p-MCA compared with the AlCl3 group (Figure 5(A)). In parallel, the anti-aging gene srl showed significant upregulation at 0.1 and 0.01 mM p-MCA (Figure 5(B)).
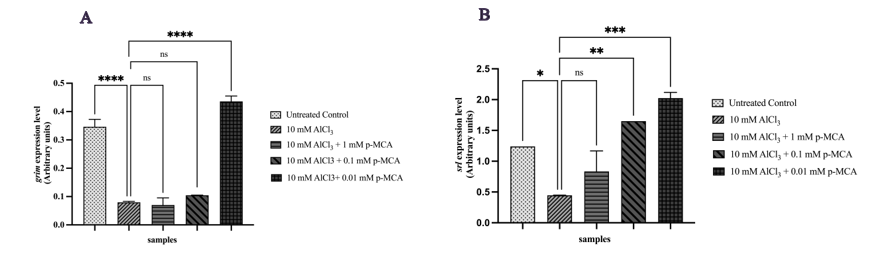
Figure 5 Relative expression levels of grim (A) and srl (B) genes in adult D. melanogaster w1118 following exposure to 10 mM AlCl3 and p-MCA at different concentrations, as determined by RT-qPCR and normalized to rp49. Data are presented as mean ± SD. *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001 compared with the AlCl₃-treated group; ns, not significant.
Expression of AChE was significantly increased in flies treated with p-MCA at 0.01 mM compared with the AlCl3 group (Figure 6). As AChE regulates acetylcholine turnover, this change may reflect compensatory regulation of cholinergic signaling in response to AlCl3-induced neurotoxicity. However, interpretation of AChE expression requires caution due to the use of whole-body RNA extracts, which may include non-neuronal contributions.
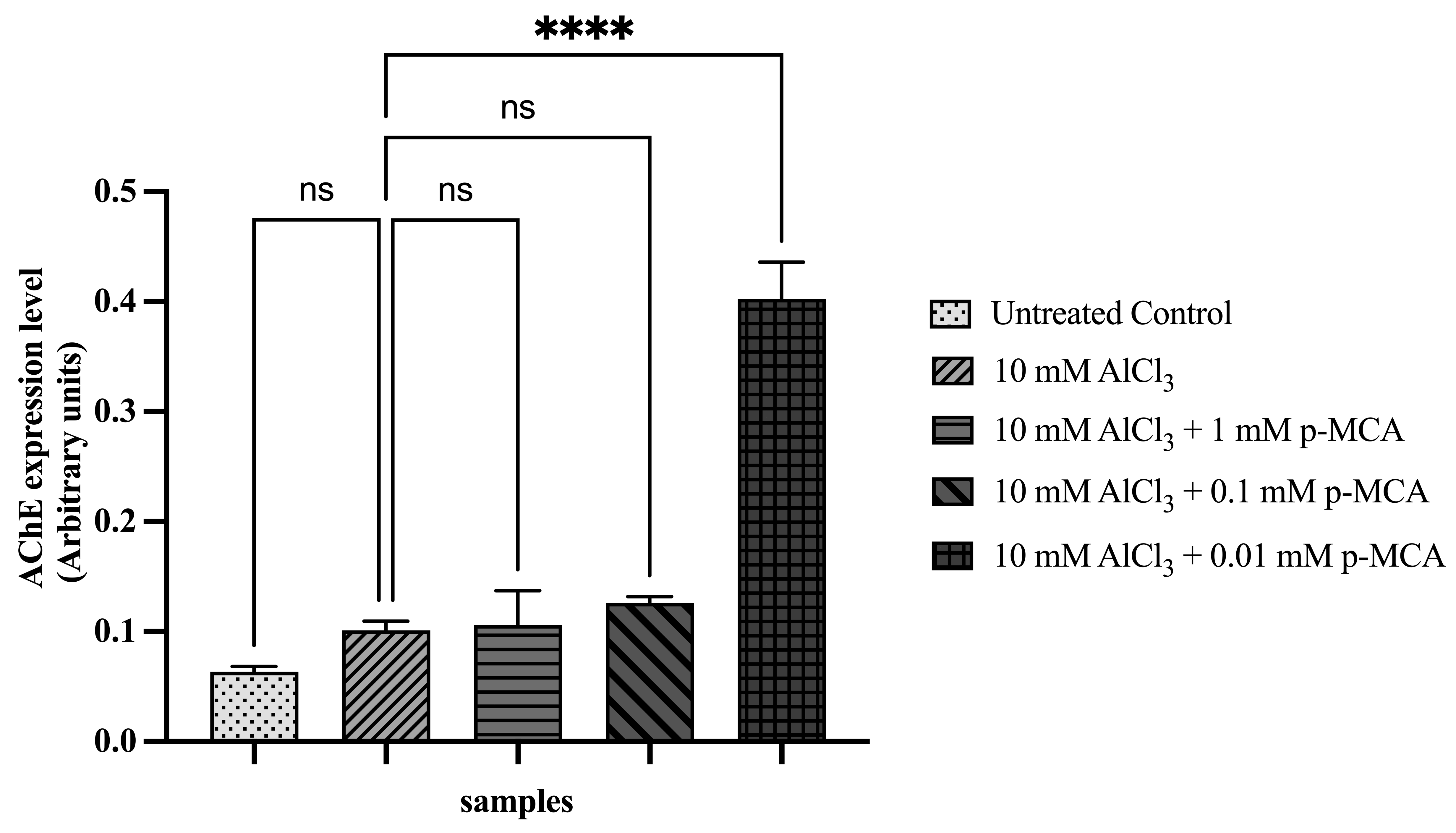
Figure 6 Relative expression level of AChE in adult D. melanogaster w1118 following exposure to 10 mM AlCl3 and p-MCA at different concentrations, as determined by RT-qPCR and normalized to rp49. Data are presented as mean ± SD. ****p < 0.0001 compared with the AlCl3-treated group; ns, not significant.
Molecular docking analysis demonstrated that p-MCA binds to D. melanogaster AChE with a binding free energy of ‒7.8 kcal/mol (Figure 7(A)), indicating moderate affinity compared to 10-40 (‒9.5 kcal/mol). p-MCA interacted with key residues within the catalytic site, supporting its inhibitory potential. Molecular dynamics simulations further revealed stable ligand-protein interactions, with RMSD and radius of gyration profiles comparable to 10-40 (Figure 7(B)).
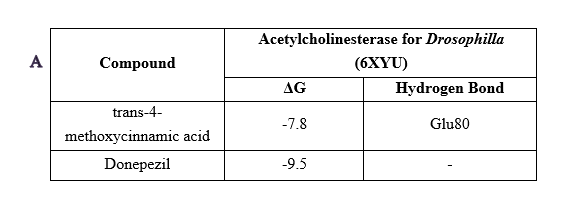
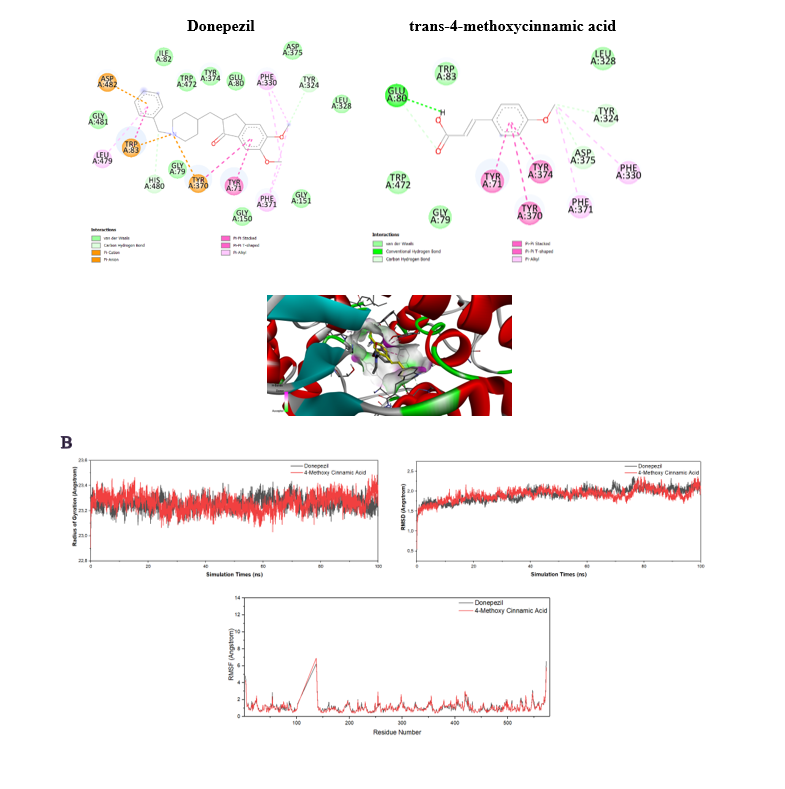
Figure 7 Results of molecular docking (A) and molecular dynamics (B) of acetylcholinesterase inhibitor in D. melanogaster.
Docking and simulation analyses were focused on AChE due to the established relevance of cholinergic dysfunction in AD and the availability of validated computational protocols. This targeted approach enabled direct correlation between molecular interactions and observed behavioral outcomes.
Docking studies using human AChE confirmed that p-MCA interacts with conserved residues, including Phe295, Tyr337, and Trp86, with a binding free energy of −7.2 kcal/mol (Figure 8(A)). Molecular dynamics simulations showed stable interaction profiles comparable to 10-40, as indicated by RMSD, RMSF, and radius of gyration analyses (Figure 8(B)). These findings suggest that p-MCA exhibits conserved inhibitory behavior across species.
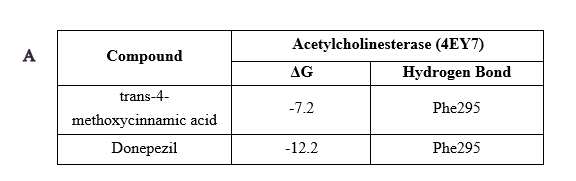
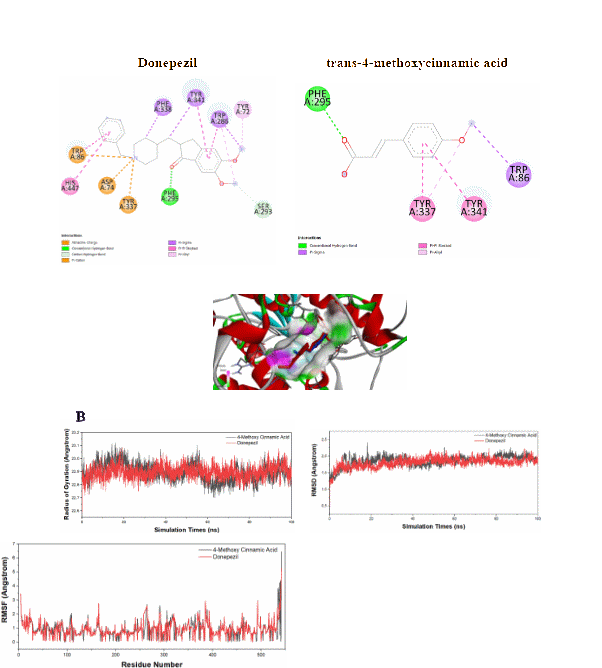
Figure 8 Results of molecular docking (A) and molecular dynamics (B) of acetylcholinesterase inhibitor in Homo sapiens.
p-MCA, a cinnamic acid derivative, has been reported to exert neuroprotective effects through antioxidant, anti-inflammatory, and apoptosis-regulating mechanisms [34,35]. In this study, its neuroprotective potential was evaluated using an integrated in vivo and in silico approach in D. melanogaster as a model of Alzheimer’s disease (AD)-like neurodegeneration. AlCl3 exposure reduced survival and impaired locomotor and cognitive performance, consistent with aluminum-induced neurotoxicity. A concentration of 10 mM produced clear neurodegenerative phenotypes without excessive mortality, providing a stable model. p-MCA, particularly at 0.01 and 0.1 mM, significantly improved survival and behavioral performance, indicating protection against oxidative and cellular damage associated with AD pathology [12,27].
Behavioral improvements in negative geotaxis and phototaxis further confirmed preservation of motor and sensory-cognitive functions, both established indicators of neurodegeneration in Drosophila. Dose selection for mechanistic assays was guided by preliminary survival screening to ensure biologically active yet non-toxic concentrations. At the molecular level, p-MCA modulated genes involved in oxidative stress, mitochondrial function, apoptosis, and aging. Upregulation of sod1 suggests enhanced antioxidant capacity and improved redox homeostasis, which is critical given neuronal vulnerability to reactive oxygen species (ROS) and mitochondrial dysfunction in AD [28,29].
Increased expression of srl, the Drosophila homolog of PGC-1α, supports improved mitochondrial biogenesis and bioenergetic resilience under neurotoxic stress [30]. In contrast, reduced hsp22 expression likely reflects attenuation of mitochondrial stress rather than diminished protection [31], consistent with concurrent sod1 and srl upregulation. Upregulation of the pro-apoptotic gene grim indicates modulation of apoptosis-related pathways. In Drosophila, apoptotic signaling can occur in a regulated, context-dependent manner to eliminate damaged cells without widespread degeneration [32]. Thus, grim induction may represent adaptive cellular regulation rather than pathological neurodegeneration.
Although acetylcholinesterase (AChE) expression increased in some groups, molecular docking and dynamics simulations demonstrated stable binding of p-MCA to AChE with affinities comparable to 10-40. The apparent discrepancy between gene expression and predicted inhibition may reflect analysis of whole-body homogenates, which include non-neuronal tissues [33]. Brain-specific studies are needed to clarify cholinergic effects. The stronger protective effects observed at 0.01-0.1 mM compared to higher doses suggest a hormetic or biphasic response, commonly reported for polyphenols, where low-to-moderate concentrations activate adaptive defense pathways without additional benefit at higher levels [34,35].
Overall, p-MCA exerts multi-target neuroprotective effects in D. melanogaster by enhancing antioxidant defenses, preserving mitochondrial function, modulating apoptosis, and interacting with cholinergic targets. The integration of in vivo and in silico analyses strengthens the mechanistic interpretation and supports the suitability of Drosophila for early-stage neuroprotective screening. However, translational limitations must be acknowledged. As an invertebrate model, Drosophila may not fully reflect vertebrate systems [36]. Furthermore, analyses were limited to gene expression in whole-body homogenates without protein-level validation, functional cellular assays, or histopathological evaluation [33,37,38]. Future studies should validate these findings in vertebrate models, including zebrafish or mammals, and incorporate protein-level, histological, and safety assessments to better establish the translational potential of p-MCA in AD.
Conclusions
This study demonstrates the neuroprotective potential of p-MCA against Alzheimer’s disease (AD)-like pathology through multi-target mechanisms. In D. melanogaster, p-MCA improved survival, locomotor activity, and cognitive-related behavior following AlCl3-induced neurotoxicity. Gene expression analyses indicated modulation of antioxidant, apoptosis-related, and aging-associated pathways, supporting enhanced cellular resilience under neurodegenerative stress. In silico docking showed strong binding affinity of p-MCA to acetylcholinesterase, comparable to 10-40, suggesting potential cholinergic modulation. The divergence between AChE expression and predicted inhibition underscores the need for tissue-specific validation. Collectively, these integrated phenotypic, molecular, and computational findings support p-MCA as a potential multi-target neuroprotective candidate that warrants further preclinical and translational investigation.
Acknowledgements
The authors would like to express their sincere gratitude to Prof. Takayuki Kuraishi from Kanazawa University, Ishikawa 920-1192, Japan, for his invaluable support in providing the necessary materials and special consumables, particularly the wild-type strain of D. melanogaster used in this study. Furthermore, the authors extend their acknowledgments to Prof. Elly Wahyuddin (Biofarmaka Laboratory, Faculty of Pharmacy, Universitas Hasanuddin, Indonesia), for generously providing all the required instruments to carry out this research. This research was funded by the Directorate General of Higher Education, Ministry of Education, Culture, Research, and Technology, Indonesia, through the PDD 2024 research grant (Contract Number 027/E5/PG.02.00.PL/2024), awarded to B.J.L and colleagues.
Declaration of Generative AI in Scientific Writing
This study used artificial intelligence (AI) tools and methodologies in the following capacity: Manuscript Writing Support: AI-based language model ChatGPT was employed in the language refinement (improving grammar, sentence structure, and readability of the manuscript). We confirm that all AI-assisted processes were critically reviewed by the authors to ensure the integrity and reliability of the results. The final decisions and interpretations presented in this article were solely made by the authors.
CRediT Author Statement
Nur Rahma Rumata: Conceptualization; Methodology; Investigation; Formal analysis; Writing – Original draft preparation, Data curation. Asbah Asbah: Investigation; Formal analysis; Data curation; Validation. Nadila Pratiwi Latada: Investigation; Visualization; Data curation. Muhammad Arfandy Gunawan: Software; Formal analysis; Visualization. Fifi Dismayanti Indriani Nainu: Resources; Validation; Writing – Review & Editing. Irmanida Batubara: Methodology; Resources; Supervision. Muhammad Aswad: Project administration; Resources; Supervision. Berry Juliandi: Conceptualization; Supervision; Writing – Review & Editing; Funding acquisition. Firzan Nainu: Conceptualization; Methodology; Writing – Review & Editing; Supervision.
References
[1] J Guo, X Huang, L Dou, M Yan, T Shen, W Tang and J Li. Aging and aging-related diseases: From molecular mechanisms to interventions and treatments. Signal Transduction and Targeted Therapy 2022; 7, 391.
[2] Y Li, X Tian, J Luo, T Bao, S Wang and X Wu. Molecular mechanisms of aging and anti-aging strategies. Cell Communication and Signaling 2024; 22, 285.
[3] E Passeri, K Elkhoury, M Morsink, K Broersen, M Linder, A Tamayol, C Malaplate, FT Yen and E Arab-Tehrany. Alzheimer’s disease: Treatment strategies and their limitations. International Journal of Molecular Sciences 2022; 23(22), 13954.
[4] DC Miculas, PA Negru, SG Bungau, T Behl, SSU Hassan and DM Tit. Pharmacotherapy evolution in Alzheimer’s disease: Current framework and relevant directions. Cells 2023; 12(1), 131.
[5] JW Kinney, SM Bemiller, AS Murtishaw, AM Leisgang AM Salazar and BT Lamb. Inflammation as a central mechanism in Alzheimerʼs disease. Alzheimer’s & dementia 2018; 4, 575-590.
[6] Y Zhang, H Chen, R Li, K Sterling and W Song. Amyloid β-based therapy for Alzheimer’s disease: Challenges, successes and future. Signal Transduction and Targeted Therapy 2023; 8, 248.
[7] PT Kamatham, R Shukla, DK Khatri and LK Vora. Pathogenesis, diagnostics, and therapeutics for Alzheimerʼs disease: Breaking the memory barrier. Ageing Research Reviews 2024; 101, 102481.
[8] E Hogervorst, E Schröder-Butterfill, YS Handajani, P Kreager and TB W Rahardjo. Dementia and dependency vs. proxy indicators of the active ageing index in Indonesia. International Journal of Environmental Research and Public Health 2021; 18(16), 8235.
[9] J Zhang, Y Zhang, J Wang, Y Xia, J Zhang and L Chen. Recent advances in Alzheimer’s disease: Mechanisms, clinical trials and new drug development strategies. Signal Transduction and Targeted Therapy 2024; 9, 211.
[10] B Miziak, B Błaszczyk and SJ Czuczwar. Some candidate drugs for pharmacotherapy of Alzheimer’s disease. Pharmaceuticals 2021; 14(5), 458.
[11] LK Huang, SP Chao and CJ Hu. Clinical trials of new drugs for Alzheimer disease. Journal of Biomedical Science 2020; 27, 18.
[12] A Płowuszyńska and A Gliszczyńska. Recent developments in therapeutic and nutraceutical applications of p-MCA from plant origin. Molecules 2021; 26(13), 3827.
[13] T Begum, R Gogoi, N Sarma, SK Pandey and M Lal. Novel ethyl p-methoxy cinnamate rich Kaempferia galanga (L.) essential oil and its pharmacological applications: Special emphasis on anticholinesterase, anti-tyrosinase, α-amylase inhibitory, and genotoxic efficiencies. PeerJ 2023; 11, 14606.
[14] A Finelli, A Kelkar, HJ Song, H Yang and M Konsolaki. A model for studying Alzheimerʼs Aβ42-induced toxicity in D. melanogaster. Molecular and Cellular Neuroscience 2004; 26(3), 365-375.
[15] UB Pandey and CD Nichols. Human disease models in D. melanogaster and the role of the fly in therapeutic drug discovery. Pharmacological Reviews 2011; 63(2), 411-436.
[16] H Bolus, K Crocker, G Boekhoff-Falk and S Chtarbanova. Modeling neurodegenerative disorders in D. melanogaster. International Journal of Molecular Sciences 2020; 21(9), 3055.
[17] LM Luheshi, GG Tartaglia, AC Brorsson, AP Pawar, IE Watson, F Chiti, M Vendruscolo, DA Lomas, CM Dobson and DC Crowther. Systematic in vivo analysis of the intrinsic determinants of amyloid β pathogenicity. PLoS Biology 2007; 5(11), 290.
[18] F Nainu, E Salim, RM Asri, A Hori and T Kuraishi. Neurodegenerative disorders and sterile inflammation: Lessons from a Drosophila model. The Journal of Biochemistry 2019; 166(3), 213-221.
[19] NR Rumata, D Purwaningsih, A Asbah, MF As’Ad, D Chadran, TB Emran and F Nainu. Phenotypical and molecular assessments on the pharmacological effects of curcumin in D. melanogaster. Narra J 2023; 3(2), 117.
[20] C Welch, E Johnson, A Tupikova, J Anderson, B Tinsley, J Newman, E Widman, A Alfareh, A Davis, L Rodriguez, C Visger, JP Miller-Schulze, W Lee and K Mulligan. Bisphenol a affects neurodevelopmental gene expression, cognitive function, and neuromuscular synaptic morphology in D. melanogaster. Neurotoxicology 2022; 89, 67-78.
[21] LL Vang, AV Medvedev and J Adler. Simple ways to measure behavioral responses of Drosophila to stimuli and use of these methods to characterize a novel mutant. PLoS One 2012; 7(5), 37495.
[22] C Solana-Manrique, MD Moltó, P Calap-Quintana, FJ Sanz, JV Llorens and N Paricio. Drosophila as a model system for the identification of pharmacological therapies in neurodegenerative diseases. Springer, Singapore, 2019, p. 433-467.
[23] M Aswada, R Nugrahaa, R Yuliantya, Ismaila, YM Evarya, Kasmiatib and ZP Tachrimc. Potency of bisindoles from caulerpa racemosa in handling diabetes-related complications: In silico ADMET properties and molecular docking simulations. Turkish Computational and Theoretical Chemistry 2024; 8(3), 99-107.
[24] CDK Amengor, P Biniyam, AA Brobbey, FK Kekessie, FK Zoiku, S Hamidu, P Gyan and BM Abudey. N‐substituted phenylhydrazones kill the ring stage of Plasmodium falciparum. BioMed Research International 2024; 2024, 6697728.
[25] N Mert-Ozupek, G Calibasi-Kocal, N Olgun, Y Basbinar, L Cavas and H Ellidokuz. In-silico molecular interactions among the secondary metabolites of Caulerpa spp. and colorectal cancer targets. Frontiers in Chemistry 2022; 10, 1046313.
[26] H Rasyid, NH Soekamto, S Firdausiah, R Mardiyanti, B Bahrun, Siswanto, M Aswad, WD Saputri, AAT Suma, NH Syahrir and RV Listyarini. Revealing the potency of 1, 3, 5-Trisubstituted pyrazoline as antimalaria through combination of in silico studies. Sains Malaysiana 2023; 52(10), 2855-2867.
[27] A Drakontaeidi and E Pontiki. Multi-target-directed cinnamic acid hybrids targeting Alzheimer’s disease. International Journal of Molecular Sciences 2024; 25(1), 582.
[28] J Xu, X Su, SK Burley and XFS Zheng. Nuclear SOD1 in growth control, oxidative stress response, amyotrophic lateral sclerosis, and cancer. Antioxidants 2022; 11(2), 427.
[29] A Houldsworth. Role of oxidative stress in neurodegenerative disorders: A review of reactive oxygen species and prevention by antioxidants. Brain Communications 2024; 6(1), 356.
[30] T Alqahtani, SL Deore, AA Kide, BA Shende, R Sharma, RD Chakole, LS Nemade, NK Kale, S Borah, SS Deokar, A Behera, DD Bhandari, N Gaikwad, AK Azad and A Ghosh. Mitochondrial dysfunction and oxidative stress in Alzheimer’s disease, and Parkinson’s disease, Huntington’s disease and amyotrophic lateral sclerosis-an updated review. Mitochondrion 2023; 71, 83-92.
[31] S Tripathi and S Sinha. Small heat shock protein and D. melanogaster development. Advances in Zoology and Botany 2023; 11(4), 246-256.
[32] CJ Sproston, JE Rak, EC Marin, S Kondo and DW Williams. Hemilineage-specific deployment of the pro-apoptotic RHG genes reaper and grim during neurogenesis sculpts segment and sex-specific neural network composition in Drosophila. Development 2025; 152(21), dev204902.
[33] OB Ogunsuyi, OC Olagoke, BA Afolabi, G Oboh, OM Ijomone, NV Barbosa and JBT da Rocha. Dietary inclusions of Solanum vegetables mitigate aluminum-induced redox and inflammation-related neurotoxicity in D. melanogaster model. Nutritional Neuroscience 2022; 25(10), 2077-2091.
[34] EJ Calabrese, P Pressman, AW Hayes, G Dhawan, R Kapoor, E Agathokleous, LA Baldwin and V Calabrese. The chemoprotective hormetic effects of rosmarinic acid. Open Medicine 2024; 19(1), 20241065.
[35] P Perrone and S D’Angelo. Hormesis and health: Molecular mechanisms and the key role of polyphenols. Food Chemistry Advances 2025; 7, 101030.
[36] Y Liu. Zebrafish as a model organism for studying pathologic mechanisms of neurodegenerative diseases and other neural disorders. Cellular and Molecular Neurobiology 2023; 43, 2603-2620.
[37] A Samih, MADM Ferreira and Z Nikoloski. Gene expression and protein abundance: Just how associated are these molecular traits? Biotechnology Advances 2026; 86, 108720.
[38] N Dhiman, S Deshwal, R Vikas, NK Singhal and S Rajat. Zebrafish as a model organism to study sporadic Alzheimerʼs disease: Behavioural, biochemical and histological validation. Experimental Neurology 2025; 383, 115034.