Trends
Sci.
2026;
23(10):
13296
Modulation of Potassium Ion Transport in Mitochondria by Kaempferol and Its Glycosides
Boburbek Yuldoshev1, Nurali Ergashev1,2,*, Esokhon Komilov1,3,
Ulugbek Gayibov4, Izzatullo Abdullaev4, Khasan Kayumov3,
Оysara Tojikulova2, Doniyor Siddikov5, Dildora Isamukhamedova6,
Lubov Kuchkarova3, Shakhriyor Sharipov7 and Muzaffar Asrarov1
1Institute of Biophysics and Biochemistry at the National University of Uzbekistan, Tashkent, Uzbekistan
2Department of Human and Animal Physiology and Biochemistry, Samarkand State University, Samarkand, Uzbekistan
3Department of Human and animals Physiology, National University of Uzbekistan, Tashkent, Uzbekistan
4Institute of Bioorganic Chemistry, Academy of Science of Uzbekistan, Tashkent, Uzbekistan
5Institute of the Chemistry of Plant Substances, Academy of Sciences of Uzbekistan, Tashkent, Uzbekistan
6Department of Clinical Foundations of Deaf Pedagogy and Special Pedagogy, National Pedagogical University of Uzbekistan Named after Nizami, Tashkent, Uzbekistan
7Department of Clinical Biochemistry, Central Asian University and AKFA Medline University Hospital,
Tashkent, Uzbekistan
(*Corresponding author’s e-mail: [email protected])
Received: 8 January 2026, Revised: 26 March 2026, Accepted: 10 April 2026, Published: 15 May 2026
Abstract
Several types of potassium channels are embedded in the inner mitochondrial membrane and play a crucial role in cellular function. Among them, the ATP-sensitive potassium channel located in the mitochondrial inner membrane (mitoKATP) is known to protect organs and tissues from ischemia-reperfusion injury. In recent years, increasing attention has been devoted to investigating the effects of flavonoids-widely distributed plant-derived secondary metabolites-on various physiopathological processes occurring in the organism. Accordingly, the present study examined the effects of kaempferol and its glycosides isolated from Geranium rotundifolium on mitochondrial ATP-sensitive potassium channel activity in an ATP- and Mg2+-dependent manner, as well as their influence on K+ ion transport. At a constant ATP concentration of 200 μM, kaempferol and its glycosides activated mitoKATP activity in a dose-dependent manner, with their effectiveness decreasing in the following order: kaempferol > kaempferitrin > kaempferol-7-O-rhamnoside > afzelin. At lower ATP concentrations, kaempferol induced weaker channel activation; however, increasing ATP levels enhanced the degree of mitoKATP activation by kaempferol. Simultaneously, increasing the concentration of Mg2+ ions resulted in a reduction of kaempferol-induced mitoKATP activation. In the presence of the K+ ionophore valinomycin, kaempferol more effectively counteracted the succinate-driven reverse K+ influx into the mitochondrial matrix compared to other flavonoids. To gain mechanistic insight at the molecular level, in silico docking analysis was performed using the human mitochondrial ATP-binding cassette transporter ABCB8. The docking results revealed stable binding of kaempferol and its glycosides within the ligand-recognition cavity of ABCB8, with binding energies ranging from –6.9 to –8.8 kcal/mol, and indicated a higher affinity of glycosylated flavonoids compared to the aglycone. These interactions were stabilized by hydrogen bonding and hydrophobic contacts with key transmembrane residues, suggesting a potential modulatory role of flavonoids on mitochondrial transport processes. In conclusion, kaempferol and its glycosides may provide protective effects against ischemia-reperfusion injury in the liver and other organs under various stress conditions by modulating the activity of mitoKATP and regulating K+ ion transport.
Keywords: Liver mitochondria, mitoKATP, ATP, Mg2+, valinomycin, kaempferol and its glycosides
Introduction
Several potassium channels have been identified in the inner mitochondrial membrane, and their crucial roles in regulating cell survival and death have been demonstrated. One of these is the ATP-sensitive potassium channel (mitoKATP) located in the inner mitochondrial membrane [1]. This channel was first described in the 1990s by Inoue and colleagues [2] as a K+-conducting channel in cardiac mitochondria that is inhibited by ATP. Therefore, this channel has been designated as the mitoKATP in mitochondria. The mitoKATP has been identified in the mitochondria of various tissues in both vertebrate and invertebrate animals, as well as in plant mitochondria [3-5]. Through this channel, the influx of K+ ions into the mitochondrial matrix contributes to the protection of tissues against ischemia–reperfusion injury [6].
ATP and Mg2+ ions have been shown to inhibit the activity of the mitoKATP, whereas diazoxide [6], GTP [7], as well as uridine and its nucleotides [8,9], have been demonstrated to activate the channel. Using molecular docking analysis, the binding of GTP and ATP to the human nucleotide-binding domain of ABCB8 (ATP-Binding Cassette Subfamily B Member 8)/mitoSUR (sulfonylurea receptor) has been identified. In this context, ATP exhibited a half-maximal inhibitory concentration for the mitoKATP channel of IC50 = 21.24 ± 1.4 μM, whereas GTP demonstrated a half-maximal activating concentration of EC50 = 13.19 ± 1.33 μM [7]. Additionally, it has been shown that inhibitory Mg2+ ions of the mitoKATP channel exert their effect from the mitochondrial matrix side, thereby influencing the channel’s conductivity and its open–closed state [6]. Furthermore, uridine and its nucleotides, through their action on mitoKATP channels, were found to enhance tolerance in susceptible animals compared to resistant ones [8].
It has been demonstrated that, following partial hepatectomy, diazoxide, an activator of the mitoKATP channel, enhances liver regeneration and is associated with increased ATP accumulation in hepatic tissue. This effect indicates that activation of the mitoKATP channel supports mitochondrial energy processes [10]. Moreover, diazoxide has been shown to stabilize liver mitochondria, increase tolerance to ischemia-reperfusion injury, and reduce the systemic inflammatory response [11]. Furthermore, it has been shown that remote ischemic preconditioning in combination with the mitoKATP channel activator diazoxide effectively protects the liver from ischemic injury while reducing pro-inflammatory cytokines. This protective effect is associated with the inhibition of the HMGB1 (high-mobility group protein B1)-induced TLR4 (Toll-like receptor 4)/MyD88 (myeloid differentiation primary response 88)/NF-κB (nuclear factor kappa-light-chain-enhancer of activated B cells) signaling pathway by mitoKATP channel activation [12]. These findings highlight that the mitoKATP channel and its activators are key mediators of hepatic protection against ischemia-reperfusion injury.
Moreover, plant-derived bioactive compounds, including flavonoids and flavonoid-containing plant extracts [13-16], as well as other types of bioactive molecules [17,18], have been identified as potential modulators of the mitoKATP channel. Flavonoids are widely distributed secondary metabolites in the plant kingdom and can be found in various plant tissues. Historically, plant materials rich in flavonoids have been extensively used in traditional medicine. Flavonoids, particularly kaempferol and its glycosides, are potent antioxidant bioactive compounds that exhibit a broad range of biological activities, including anti-inflammatory, anti-reprotoxic, neuroprotective, anti-apoptotic, pro-apoptotic, cardioprotective, hepatoprotective, anti-diabetic, anti-cancer, anti-allergic, nephroprotective, and immunomodulatory effects, among others [19-31].
Flavonoids have been identified as modulators of mitochondrial K+ channels, including mitoKATP channels, Ca2+-activated K+ channels (mitoBKCa), voltage-dependent K+ channels (mitoKv), 2-pore domain acid-sensitive K+ channels (mitoTASK), and Na+-activated K+ channels (mitoSLO) [32]. They selectively affect K+ channels located in biological membranes, thereby regulating cellular signaling and membrane potential [16]. Additionally, flavonoids influence intracellular and tissue signaling pathways, contributing to the regulation of ATP synthesis, mitochondrial biogenesis, autophagy, cell division, and fusion processes [33].
Considering the aforementioned findings, in vitro studies were conducted to investigate the effects of kaempferol and its glycosides, isolated from Geranium rotundifolium, on mitoKATP channel activity in the presence of ATP and Mg2+ ions, as well as their influence on K+ ion transport.
Materials and methods
Materials
Kaempferol and its glycosides were employed in the experiments, and their structural formula are illustrated in Figure 1. The kaempferol and its glycosides kaempferol-7-O-rhamnoside, kaempferitrin, and afzelin were extracted from Geranium rotundifolium, each with a purity greater than 97 %. In the experiments, solutions of flavonoids dissolved in DMSO were used. Diazoxide, KCl, glibenclamide, valinomycin, and EDTA which are used in the experiments purchased by Sigma-Aldrich company. K2HPO4, KH2PO4, sodium succinate, MgSO4, and Tris-HCl were purchased from Reachim (Russia). Sucrose and other reagents were obtained from Macklin (China).
Сare and handing of animals
The study was carried out on adult male albino rats weighing 180 - 200 g. Additionally, more than 50 3-month-old rats were utilized in the study. The animals were maintained under standard vivarium conditions with strict control of diet and housing. All experimental procedures adhered to the International Helsinki Declaration, the International Guiding Principles for Biomedical Research Involving Animals [CIOMS, 1985], and the bioethical guidelines of the Institute of Biophysics and Biochemistry, National University of Uzbekistan (Approval No. BEC/IBB-NUU 2024/14-1).
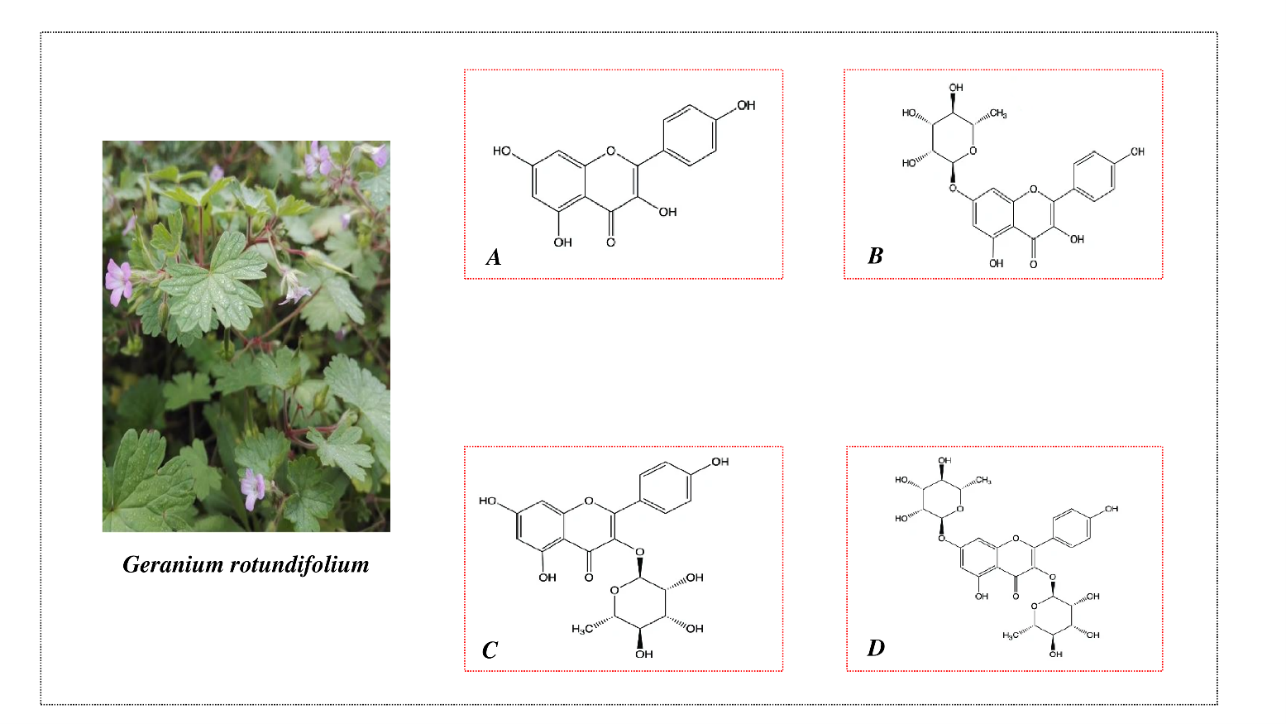
Figure 1 Geranium rotundifolium plant and the structural composition of flavonoids isolated from the plant: (A) – kaempferol, (B) – kaempferol-7-O-rhamnoside, (C) – afzelin and (D) – kaempferitrin.
Mitochondria isolation
Experimental animals were euthanized by decapitation. After decapitation, the abdominal cavity was opened, the liver was excised, and cold isolation medium was added at a ratio of 1:6. The liver was then homogenized using a Teflon homogenizer to prepare a liver homogenate. Rat liver mitochondria were isolated using the differential centrifugation method [34]. The isolation medium contained 250 mM sucrose, 10 mM Tris-HCl, and 1 mM EDTA, with a pH of 7.4. Centrifugation was carried out at ±2 °C using an RC-6CM centrifuge. First, the homogenate was spun at 1,500 rpm for 5 min to sediment nuclei and cellular debris. The resulting supernatant was then centrifuged at 6,000 rpm for 15 min to obtain the mitochondrial pellet. The mitochondria were resuspended in EDTA-free isolation medium at a 10:1 ratio and kept on ice. Protein concentration in liver mitochondria was measured by the biuret method [35] and ranged from 60 to 80 mg/mL.
Mitochondrial ATP-sensitive K+-dependent swelling
The activity of ATP-sensitive potassium channels in mitochondria was determined spectrophotometrically in a 3 mL cuvette using a V-5000 spectrophotometer at a wavelength of 540 nm, based on the dynamics of optical density changes. The incubation medium consisted of 125 mM KCl, 10 mM HEPES, 5 mM succinate, 1 mM MgSO4, 2.5 mM K2HPO4, 2.5 mM KH2PO4, and 0.005 mM rotenone, with a pH of 7.4 [36]. The protein concentration in the incubation medium was 0.3 - 0.4 mg/mL. The activity of mitoKATP channels was analyzed under conditions with ATP at concentrations of 50, 100, 150, and 200 µM, as well as Mg2+ ions at 0.5, 1.0, 1.5, and 2.0 mM. In addition, the effects of various concentrations of kaempferol and its glycosides on mitoKATP activity were investigated.
Measurement of K+ ion influx (50 mM) through the mitoKATP under hypotonic solution
In studies of mitoKATP activity in mitochondria, Mironova and her colleagues [37] characterized channel activity by examining K+ ion influx (50 mM) under hypotonic conditions. Accordingly, in the present study, the effects of different ATP concentrations were assessed in a hypotonic medium, and the incubation medium was composed as follows: 50 mM KCl, 10 mM HEPES, 5 mM succinate, 1 mM MgSO4, 2.5 mM K2HPO4, 2.5 mM KH2PO4, 0.005 mM rotenone, pH 7.4.
Swelling and shrinkage of mitochondria mediated by K+
K+ ion influx into mitochondria (swelling) and K+ ion efflux (shrinkage) in the presence of KCl were investigated using the following incubation medium: 30 mM KCl, 10 mM Tris-HCl, 1 mM EDTA, and 1 µM rotenone, pH 7.5. K+ entry into mitochondria was induced by the K+ ionophore valinomycin (0.1 µg), whereas K+ efflux from mitochondria was triggered by 2 mM sodium succinate [38]. The mitochondrial protein concentration in the medium was 0.3 - 0.4 mg/mL. The effects of kaempferol and its glycosides on these processes were examined. The swelling and shrinkage of rat liver mitochondria mediated by K+ were analyzed by measuring potassium ion concentrations in the incubation medium using an ion-selective method with a Roche/Hitachi Cobas c501 Chemistry Analyzer (Cobas 6000, USA).
Molecular docking
Molecular docking was performed to investigate the interaction of kaempferol and its glycosides with the human mitochondrial ABC transporter ABCB8 (PDB ID: 5OCH). The protein structure was prepared by removing water molecules and non-protein residues, followed by the addition of polar hydrogens and assignment of Gasteiger charges. The prepared protein was saved in PDBQT format. The structures of kaempferol, kaempferitrin, kaempferol-7-O-rhamnoside, and afzelin were energy-minimized, and rotatable bonds were defined. Polar hydrogens and Gasteiger charges were added prior to docking. Docking simulations were carried out using AutoDock Vina. The grid box was centered at center_x = 207.86, center_y = 8.40, center_z = 484.48, covering the putative ligand-binding cavity of ABCB8. The best binding poses were selected based on the lowest binding free energy values. Protein-ligand interactions were analyzed in terms of hydrogen bonding, hydrophobic interactions, and π-type interactions using molecular visualization tools [39].
Statistical analysis
Data are expressed as the mean ± standard error of the mean (S.E.M.). Statistical significance was tested by Student’s t test or paired t test. A p value less than 0.05 was considered significant.
Results and discussion
Initially, the studies were conducted to investigate the inhibitory effect of ATP at a concentration of 200 µM on mitoKATP channel activity [40]. In this case, the flavonoid kaempferol exhibited a specific effect on mitoKATP activity: At a concentration of 1 µM, it activated the mitoKATP by 14.9 ± 1.4% compared to the ATP-inhibited state, whereas at a concentration of 50 µM, it activated the mitoKATP by 123.0 ± 2.3% compared to the ATP-inhibited state (p < 0.001) (Figure 2(A)). The kaempferol glycoside - kaempferol-7-O-rhamnoside - activated the mitoKATP channel by 20.5 ± 2.7% (p < 0.05) at a concentration of 100 µM compared to the ATP-inhibited state, by 64.0 ± 2.1% (p < 0.001) at 150 µM, and by 181.1 ± 3.15% (p < 0.001) at 200 µM (Figure 2(B)).
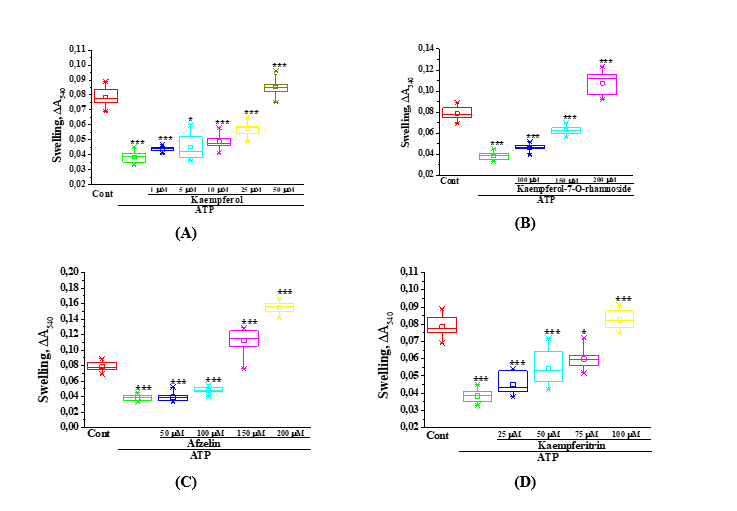
Figure 2 The effects of kaempferol and its glycosides at different concentrations on mitoKATP activity in liver mitochondria were evaluated. A - effects of kaempferol at concentrations of 1, 5, 10, 25, and 50 µM; B - effects of kaempferol-7-O-rhamnoside at 100, 150, and 200 µM; C - effects of kaempferitrin at 25, 50, 75, and 100 µM; D - effects of afzelin at 50, 100, 150, and 200 µM (* –p < 0,05, ** –p < 0.01, *** –p < 0.001; n = 5).
Another glycoside of kaempferol, namely the flavonoid kaempferitrin, activated the mitoKATP channel by 17.8 ± 4.4% at a concentration of 25 µM compared to the ATP-inhibited state, and by 115.2 ± 2.2% (p < 0.001) at a concentration of 100 µM (Figure 2(C)). In subsequent experiments, the effect of another kaempferol glycoside, the flavonoid afzelin, on mitoKATP activity was investigated (Figure 2(D)). At a concentration of 50 µM, afzelin did not exhibit an activating effect on the mitoKATP compared to the ATP-inhibited state. At 100 µM, the flavonoid increased mitoKATP activity by 25.8 ± 2.3% relative to the ATP-inhibited state, while at concentrations of 150 and 200 µM, it activated the mitoKATP by an average of 3- and 4-fold, respectively, compared to the ATP-inhibited state. The obtained results indicate that in an incubation medium containing ATP at a concentration of 200 µM, kaempferol and its glycosides activated mitoKATP activity in a dose-dependent manner. Specifically, kaempferol at 50 µM was able to activate the mitoKATP to a level comparable to that in the absence of ATP, while its glycoside kaempferitrin activated the channel at 100 µM. In addition, the other 2 glycosides of kaempferol – kaempferol-7-O-rhamnoside and afzelin - activated the mitoKATP at 150 µM to a level similar to that observed under ATP-free conditions. In the studies, kaempferol and its glycosides exhibited different biological activities. Kaempferol was active at concentrations up to 50 µM, whereas its glycosides demonstrated their activity at higher concentrations. This difference may be attributed to the presence of glycosidic moieties in their structures.
MitoKATP activity was shown to be dose-dependently inhibited in the presence of ATP [7]. Based on this, the effect of kaempferol on mitoKATP activity was investigated in the presence of varying ATP concentrations. Initially, the K+ ion permeability of intact rat liver mitochondria was assessed and found to be on average 36.4 ± 0.27 (ΔA540×100).
Subsequently, the effect of the flavonoid kaempferol on mitoKATP activity was investigated under conditions in which the channel was inhibited by ATP at concentrations of 50, 100, 150, and 200 µM. At an ATP concentration of 50 µM, mitoKATP activity was inhibited by 29.4%, corresponding to a value of 25.7 ± 3.03. At the same time, kaempferol at a concentration of 1 µM activated mitoKATP channel activity by 10.9% (28.5 ± 0.23) compared to the ATP-inhibited state,
while at concentrations of 5 and 10 µM, it increased activity by 26.1% (32.4 ± 0.33) and 45.5% (37.4 ± 0.46), respectively (Figure 3(A)). In subsequent experiments, mitoKATP activity in liver mitochondria was examined under conditions of inhibition by 100 µM ATP. At this ATP concentration, mitoKATP activity was inhibited by 53.6%, corresponding to a value of 16.9 ± 0.33. At the same time, kaempferol at a concentration of 1 µM increased mitoKATP activity by 47.9% (25.0 ± 0.47) compared to the ATP-inhibited state, while at concentrations of 5 and 10 µM, it increased activity by 53.8% (26.0 ± 0 .85) and 89.3% (32.0 ± 0.79), respectively (Figure 3(B)). In subsequent experiments, mitoKATP activity in liver mitochondria was examined under conditions of inhibition by 150 µM ATP. At this ATP concentration, mitoKATP activity was inhibited by up to 57.7%, corresponding to a value of 15.4 ± 0.33. At the same time, kaempferol at a concentration of 1 µM increased mitoKATP channel activity by 31.2% (20.2 ± 0.35) compared to the ATP-inhibited state, while at concentrations of 5 and 10 µM, it increased activity by 79.2% (27.6 ± 0.12) and 111.7% (32.6 ± 0.29), respectively (Figure 3(C)). Similarly, in subsequent experiments, mitoKATP activity was examined under conditions of inhibition by 200 µM ATP. At this ATP concentration, mitoKATP activity was inhibited by 61.0%, corresponding to a value of 14.2 ± 0.19. Thereafter, kaempferol at a concentration of 1 µM increased mitoKATP activity by 16.9% (16.6 ± 0.24) compared to the ATP-inhibited state, while at concentrations of 5 and 10 µM, it increased activity by 81.7% (25.8 ± 0.18) and 112.0% (30.1 ± 0.26), respectively (Figure 3(D)). The obtained results indicate that mitoKATP activity was increasingly inhibited as the ATP concentration in the incubation medium increased. However, the activation of the mitoKATP by the flavonoid kaempferol was dependent on the ATP concentration; at low ATP concentrations, the channel was weakly activated, whereas increasing ATP levels enhanced the effect of kaempferol.
|
|
|
|
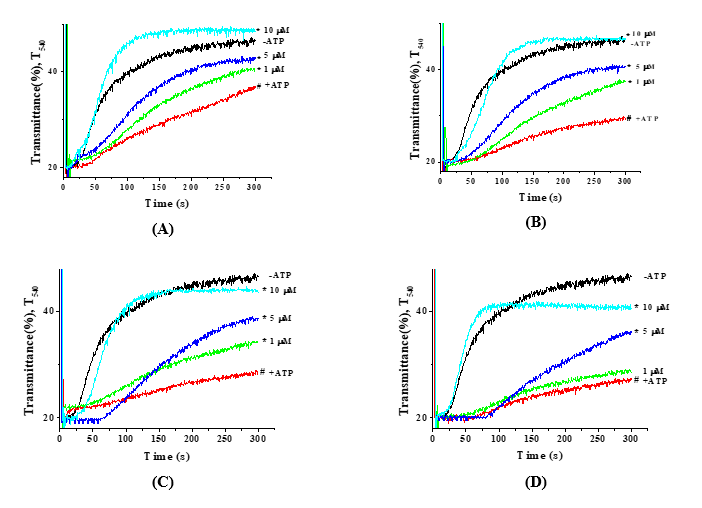
Figure 3 The effect of the flavonoid kaempferol on mitoKATP activity in rat liver mitochondria under hypotonic solution was evaluated under inhibition by ATP at concentrations of 50 µM (A), 100 µM (B), 150 µM (C), and 200 µM (D) (* –p < 0,05; n = 5; # –p value of +ATP relative to −ATP; * –p value of kaempferol concentrations relative to +ATP).
In subsequent experiments, in the absence of ATP and at varying Mg2+ concentrations, specifically 0.5, 1.0, 1.5, and 2.0 mM, it was observed that increasing Mg2+ concentration led to a gradual decrease in mitoKATP channel activation (Figures 4(A) - (D)). However, under these conditions, ATP-mediated inhibition of mitoKATP activity was relatively reduced.
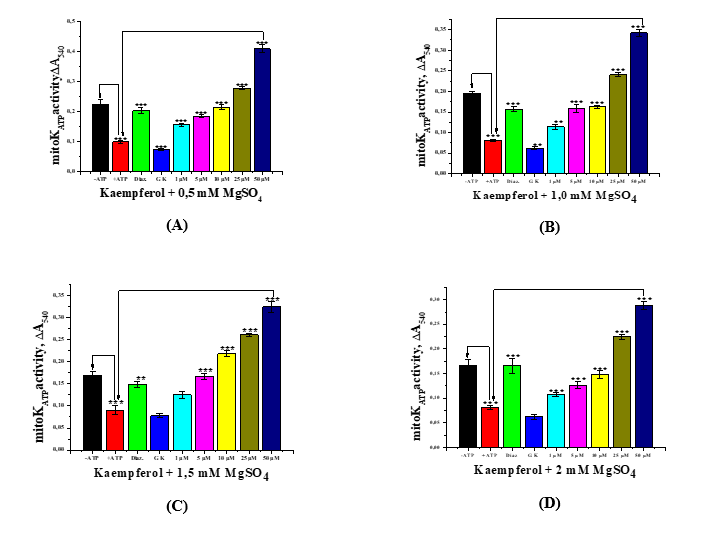
Figure 4 The effect of various concentrations of the flavonoid kaempferol on mitoKATP channel activity in liver mitochondria in media containing Mg2+ (MgSO4) at concentrations of 0.5 mM (A), 1.0 mM (B), 1.5 mM (C), and 2.0 mM (D) (* −p < 0,05, ** −p < 0.01, *** −p < 0.001; n = 5).
Similarly, the activation of the mitoKATP by diazoxide (50 µM) was found to depend on the Mg2+ concentration in the incubation medium, with increasing Mg2+ levels negatively affecting the activating effect of diazoxide. Increasing the concentration of Mg2+ ions in the incubation medium consistently manifested the inhibitory effect of glibenclamide (100 µM) on mitoKATP activity. At the same time, a specific effect of the studied flavonoid was also observed. Kaempferol at concentrations of 1, 5, 10, 25, and 50 µM acted similarly to the mitoKATP activator diazoxide, such that increasing Mg2+ levels in the incubation medium reduced the activation of the mitoKATP by kaempferol. This indicates that Mg2+ ions also exert a modulatory effect on mitoKATP activity.
It has been shown that during ischemic preconditioning, changes in K+ ion levels in mitochondria occur primarily as a result of the activation of mitoKATP and associated regulatory proteins [40]. Accordingly, K+ ions, in the presence of the ionophore valinomycin, induce swelling of energized mitochondria and promote the oxidation of extramitochondrial NADH/cytochrome c, which is associated with dissipation of the mitochondrial membrane potential (ΔΨm) [41] and the mitochondrial matrix becomes acidified, resulting in the influx of K⁺ ions into the mitochondria [40,42].
Under the same conditions, the effect of kaempferol and its glucosides on the swelling and shrinkage of mitochondria in the presence of valinomycin and succinate was investigated. Kaempferol at concentrations of 10, 20, 30, 40, and 50 µM had little effect on mitochondrial swelling induced by K+ influx in the presence of valinomycin, but it was found to reduce the shrinkage of mitochondria triggered by succinate in a concentration-dependent manner (Figure 5(A)).
Kaempferol-7-O-rhamnoside, in contrast to kaempferol, was found to increase mitochondrial swelling induced by K+ influx in the presence of valinomycin in a concentration-dependent manner. At the same time, it was shown to reduce succinate-induced mitochondrial shrinkage in a concentration-dependent manner (Figure 5(B)). Afzelin, at the studied concentrations, had little effect on mitochondrial swelling induced by K+ influx in the presence of valinomycin. Although it reduced succinate-induced mitochondrial shrinkage, no concentration-dependent changes were observed (Figure 5(C)). The next studied flavonoid, kaempferitrin, had little effect on mitochondrial swelling induced by K+ influx in the presence of valinomycin. However, it was shown to slightly reduce succinate-induced mitochondrial shrinkage, associated with K+ efflux, in a concentration-dependent manner (Figure 5(D)).
Thus, the results indicate that, at the studied concentrations, kaempferol significantly counteracts succinate-induced mitochondrial shrinkage associated with K+ efflux, to a greater extent than the other flavonoids.
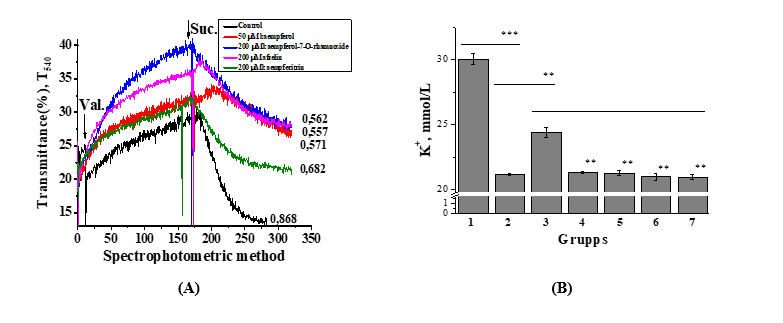
Figure 5 Effect of various concentrations of kaempferol and its glucosides on K+ influx- and efflux-related changes in liver mitochondria – A (Spectrophotometric (n = 5)) and ion-selective determination of K+ ions in solution – B (** −p < 0.01, *** −p < 0.001; n = 3 - 5). Mitochondrial swelling associated with K+ influx was induced by 0.1 µg valinomycin, while shrinkage associated with K+ efflux was triggered by 2 mmol/L Na succinate. Ion-selective determination of K+ ions in solution: 1-control - incubation medium; 2-valinomycin (0.1 μg); 3-Na succinate (2.0 mM/L); 4-kaempferol (50 μM/L); 5-kaempferol-7-ramnoside (200 μM/L); 6-kaempferitrin (200 μM/L); 7-afzelin (200 μM/L).
At present, the concentration of K+ ions in the solution were investigated using an ion-selective method (Figure 5(B)). The results demonstrated that the K+ ion concentration in the incubation medium was 30.05 ± 0.51 mM/L. However, the addition of liver mitochondria together with valinomycin (0.1 µg) to the incubation medium led to a significant decrease in K+ ion concentration by 29.6%, reaching a value of 21.15 ± 0.08 mM/L. The addition of 2.0 mM succinate to the valinomycin-containing medium resulted in a relative restoration of the K+ ion concentration in the incubation medium by 14.2%, reaching 24.15 ± 0.99 mM/L. Kaempferol and its glycosides-kaempferol-7-O-rhamnoside, kaempferitrin, and afzelin–reduced the K⁺ ion concentration in the incubation medium, thereby promoting a certain degree of K+ ion retention within mitochondria. Following kaempferol treatment, the K+ ion concentration in the medium was 21.31 ± 0.06 mM/L, whereas exposure to kaempferol-7-O-rhamnoside, kaempferitrin, and afzelin resulted in values of 21.27 ± 0.17, 20.99 ± 0.27, and 20.97 ± 0.17 mM/L, respectively. Notably, compared with the effect of succinate, treatment with flavonoids led to a further decrease in the K⁺ ion concentration in the incubation medium. These findings are generally consistent with the results presented in Figure 5(A).
Thus, the obtained results indicate that the flavonoid kaempferol, at the studied concentrations, significantly attenuates the contraction process associated with the release of K+ ions from mitochondria induced by succinate, compared with the other flavonoids.
Molecular docking studies were performed to evaluate the binding affinity and interaction patterns of selected flavonoids with the human ABCB8 protein (PDB ID: 5OCH) in its outward-facing conformation. Docking simulations were carried out within a defined binding region using the grid box centered at center x = 207.86, center_y = 8.40, center_z = 484.48, which corresponds to the putative ligand-recognition cavity of the transporter. The obtained docking poses revealed stable ligand accommodation within the binding pocket, supported by multiple noncovalent interactions with key amino acid residues (Figure 6).
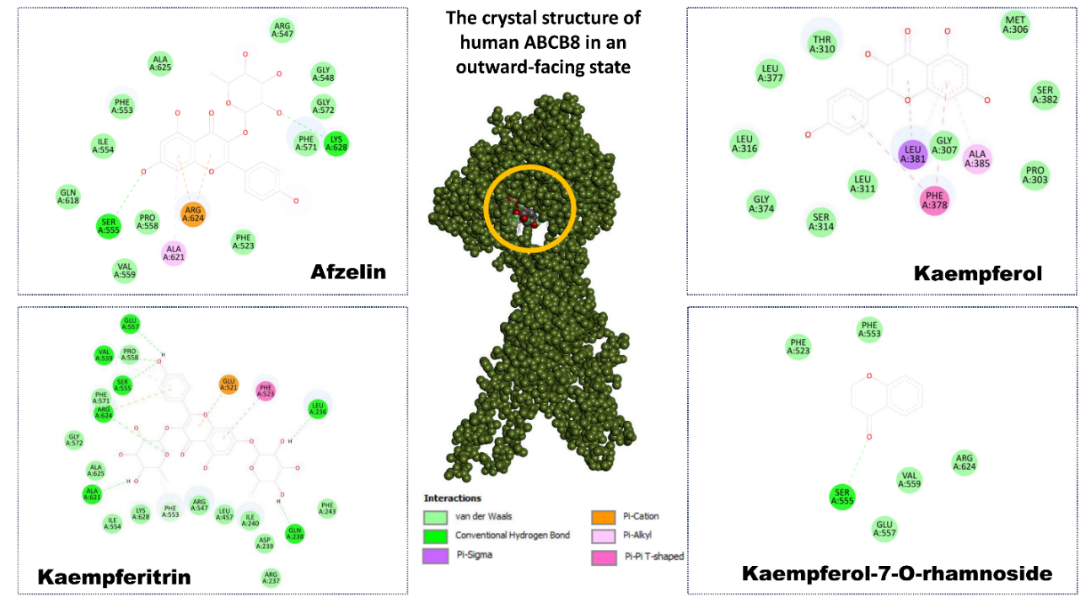
Figure 6 Molecular docking analysis of flavonoids isolated from Geranium rotundifolium with the human mitochondrial ABC transporter ABCB8 (PDB ID: 5OCH). The central panel shows the crystal structure of ABCB8 in the outward-facing conformation, with the ligand-binding cavity highlighted. Surrounding panels illustrate the 2-dimensional interaction maps of afzelin, kaempferol, kaempferitrin, and kaempferol-7-O-rhamnoside within the binding pocket. Key amino acid residues involved in ligand stabilization are indicated, along with noncovalent interactions, including hydrogen bonds, π–cation, π–alkyl, π–π T-shaped, and van der Waals contacts.
Among the investigated compounds, kaempferitrin exhibited the strongest binding affinity toward ABCB8, with a docking score of –8.8 kcal/mol. The ligand formed an extensive interaction network involving conventional hydrogen bonds with residues such as Ser555, Glu557, and Gln238, as well as π–alkyl and π–cation interactions with aromatic and charged residues including Phe523, Phe553, Arg624, and Lys628. Additional van der Waals contacts with Ala621, Ala625, Val559, and Ile554 further stabilized the ligand within the binding pocket, indicating a highly favorable binding mode.
Kaempferol-7-O-rhamnoside also demonstrated strong binding affinity (–8.2 kcal/mol) and occupied a similar region of the binding cavity. The interaction profile was dominated by hydrogen bonding with Ser555 and Glu557, complemented by hydrophobic interactions involving Val559, Arg624, and Phe523. The presence of the sugar moiety contributed to additional polar contacts, which likely enhanced ligand anchoring and overall binding stability.
In comparison, afzelin displayed a moderate yet stable binding affinity (–7.7 kcal/mol). Its docking pose was characterized by π–cation interaction with Arg624, hydrogen bonding with Ser555, and hydrophobic contacts involving Phe553, Ile554, Val559, and Ala625. These interactions suggest that afzelin is well accommodated within the ABCB8 binding pocket, although with a slightly reduced interaction density relative to kaempferitrin. Kaempferol, the aglycone form, showed the weakest binding among the tested compounds (–6.9 kcal/mol). Its interaction pattern mainly consisted of π–π T-shaped and π–alkyl interactions with aromatic residues such as Phe378 and Leu381, along with limited hydrogen bonding. The absence of glycosidic substituents reduced the number of polar contacts, resulting in lower binding affinity compared to its glycosylated derivatives.
Importantly, the present molecular docking analysis provides additional mechanistic insight into the mitochondrial actions of flavonoids by demonstrating their ability to directly interact with ABCB8. Docking simulations revealed stable binding of afzelin, kaempferol, kaempferitrin, and kaempferol-7-O-rhamnoside within the ligand-recognition cavity of ABCB8, with binding energies ranging from –6.9 to –8.8 kcal/mol. Among these compounds, glycosylated flavonoids exhibited stronger binding affinities than the aglycone kaempferol, suggesting that sugar moieties substantially enhance ligand-protein interactions through additional hydrogen bonding and polar contacts.
At the molecular level, the interaction patterns were characterized by a combination of conventional hydrogen bonds, π–cation, π–alkyl, and van der Waals interactions involving functionally relevant amino acid residues such as Ser555, Glu557, Arg624, Lys628, Phe523, and Phe553, which are located within the transmembrane region of ABCB8 responsible for substrate recognition and transport. The formation of these interaction networks indicates that flavonoids can be stably accommodated within the ABCB8 binding cavity and may influence transporter conformational dynamics and ATP-dependent transport activity.
Overall, the docking results demonstrate that all investigated flavonoids are capable of binding within the ABCB8 ligand-recognition cavity. Notably, glycosylated flavonoids exhibited stronger binding affinities and more extensive interaction networks than the aglycone kaempferol, highlighting the important role of sugar moieties in stabilizing ligand-protein interactions. These findings suggest that afzelin, kaempferitrin, and kaempferol-7-O-rhamnoside may act as potential modulators of ABCB8 function, providing a structural basis for their possible involvement in mitochondrial transport regulation.
Based on the results of these studies, it can be concluded that the entry of K+ ions into mitochondria via the mitoKATP channel or in the presence of valinomycin, under steady-state conditions, leads to an increase in mitochondrial matrix volume, stimulation (uncoupling) of respiration, and alkalization of the matrix [43], while simultaneously causing a decrease in reactive oxygen species (ROS) generation [7,40].
However, it has also been shown that activation of the mitoKATP channel may temporarily lead to a certain increase in ROS levels [44]. It has been determined that during ischemia/reperfusion processes, stimulation of the mPTP leads to a weakening of the K+/H+ antiporter function and acidification of the mitochondrial matrix, thereby facilitating the influx of phosphate ions. Conversely, the entry of K+ ions into mitochondria has been shown to protect cells from ischemia/reperfusion injury [45]. Activation of the mitoKATP channel by diazoxide during hepatic ischemia/reperfusion injury has been shown to confer cytoprotective effects by protecting the organ from ischemia/reperfusion-induced damage [11]. Specifically, based on conducted experiments, it can be inferred that kaempferol and its glycosides may exert cytoprotective effects in rat liver during ischemia/reperfusion by activating the mitoKATP channel in hepatic mitochondria.
Furthermore, biologically active compounds with cytoprotective properties have been reported to improve mitochondrial function by activating the SIRT1 (sirtuin 1)/AMPK (AMP-activated protein kinase) signaling pathway, reduce TLR4 (Toll-like receptor 4)/NF-κB (nuclear factor kappa-light-chain-enhancer of activated B cells) activity, thereby inhibiting the production of proinflammatory cytokines, and modulate the TGF-β (transforming growth factor-β)/Smad (Small mothers against decapentaplegic) signaling pathway to attenuate the activation of hepatic stellate cells [46]. This process may also involve partial contributions from mitoKATP channel activation, cyclooxygenase-1 activity, and nitric oxide modulation [47].
It has been established that the pore-forming subunit of the mitoKATP channel is MITOK, encoded by the CCDC51 (Coiled-Coil Domain Containing 51) gene, while the sulfonylurea receptor subunit MITOSUR, which confers sensitivity of the channel to glibenclamide, is encoded by the ABCB8 gene. MITOK and MITOSUR form the mitoKATP channel responsible for K+ ion transport into mitochondria. The MITOSUR subunit has been shown to interact with glibenclamide, 5-hydroxydecanoate, ATP, and Mg2+ ions to inhibit the channel, whereas activators such as diazoxide bind to MITOSUR to promote channel activation [3,48]. The pore-forming MITOK subunit of the mitoKATP channel is expressed in all tissues of both humans and mice, and its expression exhibits a strong correlation with MITOSUR expression across these tissues [3]. Thus, ABCB8, located in the inner mitochondrial membrane, represents a key regulatory factor that contributes to cellular energy metabolism and responses to oxidative stress [49-52].
Diazoxide has been shown to interact with mitoSUR (ABCB8), binding to specific amino acid residues within the molecule. In particular, hydrogen bonds are formed with Gln205 and Arg275, van der Waals interactions with Gln422, Asn383, Asn162, Thr209, Ala158, Thr278, and Gln279, and alkyl/π-alkyl interactions with Ala159, Tyr212, Ala155, and Arg423. In contrast, the competitive inhibitor glibenclamide interacts with mitoSUR through hydrogen bonds with Gln422, Asn383, and Ala159; carbon-hydrogen bonds with Gln279 and Gly282; van der Waals interactions with Val163, Leu156, Thr278, Asn162, Gln205, Ser382, Ser429, Gln379, Thr209, and Arg275; and alkyl/π-alkyl interactions with Leu160, Arg423, Ala426, Met425, and Phe386 [39]. Notably, the binding sites of kaempferol and its glycosides on ABCB8 have been found to differ from the binding sites of diazoxide, suggesting distinct interaction patterns.
Taken together, these findings suggest that, in addition to their well-established antioxidant and mitoKATP-modulating properties, flavonoids may exert part of their mitochondrial effects through direct interaction with ABCB8. Modulation of this transporter may contribute to the regulation of mitochondrial ion balance, redox status, and energy homeostasis, thereby reinforcing the cytoprotective effects of flavonoids observed in various pathological conditions. Thus, the combined experimental evidence and in silico results support the concept that flavonoids act as multifunctional mitochondrial modulators targeting multiple components of mitochondrial ion transport and regulatory systems.
It is evident that the high antioxidant capacity of flavonoids and their ability to modulate mitoKATP channel activity may provide effective outcomes in the correction of various pathological conditions. Notably, the mitoKATP channel plays a crucial role in both physiological and pathological processes by regulating mitochondrial volume and function while meeting the energetic demands of the cell [3]. At the same time, kaempferol and its glycosides, by promoting mitoKATP channel activation in an ATP- and Mg2+-dependent manner, facilitate the influx of K+ ions into mitochondria, protect cells from ischemia/reperfusion injury, and contribute to the preservation and improvement of mitochondrial and cellular functional status.
Conclusions
It was demonstrated that, at a constant ATP concentration of 200 μM, the activity of the mitoKATP was dose-dependently activated by kaempferol and its glycosylated derivatives. The efficacy of the tested flavonoids in activating the mitoKATP followed the order: kaempferol > kaempferitrin > kaempferol-7-O-rhamnoside > afzelin. Modulation of ATP concentration exerted a distinct effect on mitoKATP activity. Kaempferol produced only weak channel activation at low ATP concentrations; however, increasing ATP levels significantly enhanced its stimulatory effect, indicating an ATP-dependent modulation of channel sensitivity. An increase in Mg2+ ion concentration resulted in a marked reduction of kaempferol-induced mitoKATP activation, suggesting the involvement of Mg2+-dependent regulatory mechanisms in flavonoid–channel interactions. Furthermore, kaempferol was found to more effectively counteract the succinate-induced reverse flux of K+ ions into the mitochondrial matrix mediated by the K+ ionophore valinomycin, compared with the other investigated flavonoids. Also, molecular docking simulations revealed stable binding of afzelin, kaempferol, kaempferitrin, and kaempferol-7-O-rhamnoside within the ligand-recognition cavity of ABCB8, with binding energies ranging from –6.9 to –8.8 kcal/mol. This indicates that kaempferol and its glycosides, by interacting with the mitochondrial KATP channel, protect against ischemia-reperfusion and restore mitochondrial and cellular function. The observed effects of flavonoids highlight the importance of structural features in determining their effects on mitochondria and support their potential role in the regulation of mitochondrial ion homeostasis and bioenergetic stability.
Acknowledgements
We would like to express our appreciation to the scientific team of the flavonoids laboratory of the Institute of Plant Substances Uzbek Academy of Sciences for kindly provided flavonoids for the research.
Declaration of generative AI in scientific writing
During the preparation of this manuscript, generative artificial intelligence (AI) tools were used only to improve the readability and language of the text. The authors reviewed and edited the content as needed and take full responsibility for the content of this publication.
CRediT author statement
N. Ergashev: Conceptualization and methodology, Writing-Original Draft, Writing-Review, English Editing, Supervision and Formal Analysis. B. Yuldoshev: Conceptualization and Methodology, Investigation, Writing-Original Draft, Writing-Review, English Editing, Supervision. U. Gayibov and I. Abdullaev: Molecular Docking Analysis. E. Komilov: Conceptualization and Methodology, Investigation, Funding Acquisition, Supervision. K. Kayumov: Funding Acquisition. О. Tojikulova and D. Isamukhamedova:: Investigation, Funding Acquisition. D. Siddikov and S. Sharipov: Investigation. L. Kuchkarova: Writing-Review, English Editing. M. Asrarov: Conceptualization and Methodology.
References
M Krajewska, A Szewczyk, B Kulawiak and P Koprowski. Pharmacological characterization of a recombinant mitochondrial ROMK2 potassium channel expressed in bacteria and reconstituted in planar lipid bilayers. Membranes 2023; 13(3), 360.
I Inoue, H Nagase, K Kishi and T Higuti. ATP-sensitive K+ channel in the mitochondrial inner membrane. Nature 1991; 352(6332), 244-247.
A Paggio, V Checchetto, A Campo, R Menabo, G Di Marco, F Di Lisa, I Szabo, R Rizzuto and D De Stefani. Identification of an ATP-sensitive potassium channel in mitochondria. Nature 2019; 572(7771), 609-613.
M Slocinska, J Lubawy, W Jarmuszkiewicz and G Rosinski. Evidences for an ATP-sensitive potassium channel (KATP) in muscle and fat body mitochondria of insect. Journal of Insect Physiology 2013; 59(11), 1125-1132.
D Pastore, M Soccio, M Laus and D Trono. The uniqueness of the plant mitochondrial potassium channel. BMB Reports 2013; 46(8), 391-397.
M Laskowski, B Augustynek, P Bednarczyk, M Zochowska, J Kalisz, B O’Rourke, A Szewczyk and B Kulawiak. Single-channel properties of the ROMK-pore-forming subunit of the mitochondrial ATP-sensitive potassium channel. International Journal of Molecular Sciences 2019; 20(21), 5323.
PB Palacio, GC de Freitas Soares, GMB Lima, PLO Cunha, ALN Varela and HT Facundo. Competitive interaction between ATP and GTP regulates mitochondrial ATP-sensitive potassium channels. Chemico-Biological Interactions 2023; 381, 110560.
IN Man’kovskaia, VI Nosar, OS Gorbacheva, OA Gonchar, BL Gavenauskas, LV Bratus and GD Mironova. The effect of uridine on the endurance of animals with different resistance to physical stress: The role of mitochondrial ATP-dependent potassium channel. Biofizika 2014; 59(5), 941-945.
IB Krylova, EN Selina, VV Bulion, OM Rodionova, NR Evdokimova, NV Belosludtseva, MI Shigaeva and GD Mironova. Uridine treatment prevents myocardial injury in rat models of acute ischemia and ischemia/reperfusion by activating the mitochondrial ATP-dependent potassium channel. Scientific Reports 2021; 11(1), 16999.
Y Nakagawa, M Yoshioka, Y Abe, H Uchinami, T Ohba, K Ono and Y Yamamoto. Enhancement of liver regeneration by adenosine triphosphate-sensitive K+ channel opener (diazoxide) after partial hepatectomy. Transplantation 2012; 93(11), 1094-1100.
MA Nogueira, AM Coelho, SN Sampietre, RA Patzina, F Pinheiro da Silva, LA D'Albuquerque and MC Machado. Beneficial effects of adenosine triphosphate-sensitive K+ channel opener on liver ischemia/reperfusion injury. World Journal of Gastroenterology 2014; 20(41), 15319-15326.
WU Koh, J Kim, J Lee, GW Song, GS Hwang, E Tak and JG Song. Remote ischemic preconditioning and diazoxide protect from hepatic ischemic reperfusion injury by inhibiting HMGB1-induced TLR4/MyD88/NF-κB signaling. International Journal of Molecular Sciences 2019; 20(23), 5899.
SV Murzaeva, SP Belova, EI Lezhnev, LD Luk’yanova and GD Mironova. Effects of flavonoid-containing preparation Extralife on hydrogen peroxide production and functioning of mitochondrial ATP-dependent potassium channel. Bulletin of Experimental Biology and Medicine 2013; 155(6), 767-770.
LM Meng, HJ Ma, H Guo, QQ Kong and Y Zhang. The cardioprotective effect of naringenin against ischemia-reperfusion injury through activation of ATP-sensitive potassium channel in rat. Canadian Journal of Physiology and Pharmacology 2016; 94(9), 973-978.
M Rameshrad, SF Omidkhoda, BM Razavi and H Hosseinzadeh. Evaluating the possible role of mitochondrial ATP-sensitive potassium channels in the cardioprotective effects of morin in the isolated rat heart. Life Sciences 2021; 264, 118659.
M Richter-Laskowska, P Trybek, DV Delfino and A Wawrzkiewicz-Jalowiecka. Flavonoids as modulators of potassium channels. International Journal of Molecular Sciences 2023; 24(2), 1311.
UG Gayibov, EJ Komilov, RN Rakhimov, NA Ergashev, NG Abdullajanova, MI Asrorov and TF Aripov. Influence of new polyphenol compound from Euphorbia plant on mitochondrial function. Journal of Microbiology, Biotechnology and Food Sciences 2019; 8(4), 1021-1025.
DK Muratova, NA Ergashev, AV Shkinev, MI Asrarov and UK Kurbanov. Effect of songorine on the activity of ATP-dependent K+ channels and the state of megapore in rat liver mitochondria. Eksperimental’naya i Klinicheskaya Farmakologiya 2023; 84(4), 12-15.
M Akter, MS Parvin, MM Hasan, MAA Rahman and ME Islam. Anti-tumor and antioxidant activity of kaempferol-3-O-alpha-L-rhamnoside (Afzelin) isolated from Pithecellobium dulce leaves. BMC Complementary Medicine and Therapies 2022; 22(1), 169.
KM Alharthy, S Rashid, HS Yusufoglu, SI Alqasoumi, MA Ganaie and A Alam. Neuroprotective potential of Afzelin: A novel approach for alleviating catalepsy and modulating Bcl-2 expression in Parkinson’s disease therapy. Saudi Pharmaceutical Journal 2024; 32(2), 101928.
F Alrumaihi, SA Almatroodi, HOA Alharbi, WM Alwanian, FA Alharbi, A Almatroudi and AH Rahmani. Pharmacological potential of kaempferol, a flavonoid in the management of pathogenesis via modulation of inflammation and other biological activities. Molecules 2024; 29(9), 2007.
W Alam, H Khan, MA Shah, O Cauli and L Saso. Kaempferol as a dietary anti-inflammatory agent: Current therapeutic standing. Molecules 2020; 25(18), 4073.
J Cao, Y Wang, S Hu, Y Ding, Q Jia, J Zhu and H An. Kaempferol ameliorates secretagogue-induced pseudo-allergic reactions via inhibiting intracellular calcium fluctuation. Journal of Pharmacy and Pharmacology 2020; 72(9), 1221-1231.
F Hua, P Zhou, PP Liu and GH Bao. Rat plasma protein binding of kaempferol-3-O-rutinoside from Lu’an GuaPian tea and its anti-inflammatory mechanism for cardiovascular protection. Journal of Food Biochemistry 2021; 45(7), e13749.
F Hua, JY Li, M Zhang, P Zhou, L Wang, TJ Ling and GH Bao. Kaempferol-3-O-rutinoside exerts cardioprotective effects through NF-κB/NLRP3/Caspase-1 pathway in ventricular remodeling after acute myocardial infarction. Journal of Food Biochemistry 2022; 46(10), e14305.
F Kabeer, S Anjum, AA Behan, M Usman, NY Khan, M Azeem, HMU Din, MA Arshad, S Nazir, S Cheng and JA Buzdar. Kaempferol alleviates cypermethrin induced reproductive toxicity in rats via Nrf2-mediated antioxidant and apoptotic regulation with histopathological evidence: In vivo and in silico study. Journal of Molecular Histology 2025; 56(5), 278.
K Kayumov, L Kuchkarova, T Dura-Trave, S Petrenko, N Ergashev, J Abdurakhmonov, R Achilov, S Khan, V Pulatova and M Ernazarov. Immunomodulatory and histoprotective effects of luteolin and related flavonoids in autoimmune thyroiditis: Evidence from a rat model. Trends in Sciences 2025; 22(12), 10981.
S Kaur, P Mendonca and KFA Soliman. The anticancer effects and therapeutic potential of kaempferol in triple-negative breast cancer. Nutrients 2024; 16(15), 2392.
C Luo, J Yan, Y Shen, Z Sun and X Xiao. Kaempferol attenuates oxidative stress-induced injury in gastric mucosal cells by activating Nrf2/GPX4 axis to inhibit ferroptosis. Immunity, Inflammation and Disease 2026; 14(2), e70352.
DK Patel. Pharmacological activities and therapeutic potential of kaempferitrin in medicine for the treatment of human disorders: A review of medicinal importance and health benefits. Cardiovascular & Hematological Disorders-Drug Targets 2021; 21(2), 104-114.
S Sugita, Y Matsuo, M Ishida, Y Uehara, Y Eguchi, Y Denda, K Nonoyama, H Murase, T Kato, K Saito, T Sato, Y Yamakawa, H Sagawa, R Ogawa, H Takahashi, A Mitsui and S Takiguchi. Kaempferol inhibits MMP-1-mediated migration and invasion in gemcitabine-resistant pancreatic cancer cells. Nutrients 2026; 18(3), 380.
RRP Kampa, A Sek, P Bednarczyk, A Szewczyk, V Calderone and L Testai. Flavonoids as new regulators of mitochondrial potassium channels: Contribution to cardioprotection. Journal of Pharmacy and Pharmacology 2023; 75(4), 466-481.
A Kicinska and W Jarmuszkiewicz. Flavonoids and mitochondria: Activation of cytoprotective pathways? Molecules 2020; 25(13), 3060.
N Ergashev, Kh Sayfieva, R Makhmudov and M Asrarov. Effect of polyphenols isolated from Plantago major L. and Plantago lanceolata L. on mitochondrial permeability transition pore in rat liver. Trends in Sciences 2024; 21(7), 7661.
C Watters. A one-step biuret assay for protein in the presence of detergent. Analytical Biochemistry 1978; 88(2), 695-698.
OB Vadzuk and SO Kosterin. Diazoxide-induced mitochondrial swelling in the rat myometrium as a consequence of the activation of the mitochondrial ATP-sensitive K+-channel. Ukrainian Biochemical Journal 2008; 80(5), 45-51.
NV Khmil, AA Mosencov, MI Shigaeva and GD Mironova. A comparison of methods for estimating the activity of the ATP-sensitive potassium channel in mitochondria based on the effect of ATP. Biophysics 2019; 64(5), 751-754.
GF Azzone, S Massair and T Pozzan. Mechanism of active shrinkage in mitochondria. II. Coupling between strong electrolyte fluxes. Biochimica et Biophysica Acta 1976; 423(1), 27-41.
P Bezerra Palacio, AM Brito Lucas, J Varlla de Lacerda Alexandre, PL Oliveira Cunha, YI Ponte Viana, AC Albuquerque, AL Nunes Varela and HT Facundo. Pharmacological and molecular docking studies reveal that glibenclamide competitively inhibits diazoxide-induced mitochondrial ATP-sensitive potassium channel activation and pharmacological preconditioning. European Journal of Pharmacology 2021; 908, 174379.
S Morota, S Piel and MJ Hansson. Respiratory uncoupling by increased H+ or K+ flux is beneficial for heart mitochondrial turnover of reactive oxygen species but not for permeability transition. BMC Cell Biology 2013; 14, 40.
DD Lofrumento, G La Piana, V Palmitessa, DI Abbrescia and NE Lofrumento. Stimulation by pro-apoptotic valinomycin of cytosolic NADH/cytochrome c electron transport pathway-Effect of SH reagents. The International Journal of Biochemistry & Cell Biology 2016; 76, 12-18.
M Aldakkak, DF Stowe, Q Cheng, WM Kwok and AK Camara. Mitochondrial matrix K+ flux independent of large-conductance Ca2+-activated K+ channel opening. American Journal of Physiology-Cell Physiology 2010; 298(3), C530-C541.
AD Costa, CL Quinlan, A Andrukhiv, IC West, M Jaburek and KD Garlid. The direct physiological effects of mitoK(ATP) opening on heart mitochondria. American Journal of Physiology-Heart and Circulatory Physiology 2006; 290(1), H406-H415.
J Matejikova, J Kucharska, M Pinterova, D Pancza and T Ravingerova. Protection against ischemia-induced ventricular arrhythmias and myocardial dysfunction conferred by preconditioning in the rat heart: Involvement of mitochondrial K(ATP) channels and reactive oxygen species. Physiological Research 2009; 58(1), 9-19.
ES Kharechkina, AB Nikiforova and AG Kruglov. Regulation of mitochondrial permeability transition pore opening by monovalent cations in liver mitochondria. International Journal of Molecular Sciences 2023; 24(11), 9237.
P Nizinski, A Krajewska, T Oniszczuk, B Polak and A Oniszczuk. Hepatoprotective effect of kaempferol: A review. Molecules 2025; 30(9), 1913.
MA Ibrahim, SA Abdel-Gaber, EF Amin, SA Ibrahim, RK Mohammed and AM Abdelrahman. Molecular mechanisms contributing to the protective effect of levosimendan in liver ischemia-reperfusion injury. European Journal of Pharmacology 2014; 741, 64-73.
O Rodrigues Pereira, JDC Serna, CC Caldeira da Silva, H Camara, SD Kodani, WT Festuccia, YH Tseng and AJ Kowaltowski. Mitochondrial ATP-sensitive K+ channels (MitoKATP) regulate brown adipocyte differentiation and metabolism. American Journal of Physiology-Cell Physiology 2025; 329(2), C574-C584.
HC Chang, JS Shapiro, X Jiang, G Senyei, T Sato, J Geier, KT Sawicki and H Ardehali. Augmenter of liver regeneration regulates cellular iron homeostasis by modulating mitochondrial transport of ATP-binding cassette B8. eLife 2021; 10, e65158.
Y Ichikawa, M Bayeva, M Ghanefar, V Potini, L Sun, RK Mutharasan, R Wu, A Khechaduri, T Jairaj Naik and H Ardehali. Disruption of ATP-binding cassette B8 in mice leads to cardiomyopathy through a decrease in mitochondrial iron export. Proceedings of the National Academy of Sciences 2012; 109(11), 4152-4157.
S Li, Y Ren, X Lu, Y Shen and X Yang. Cryo-EM structure of human ABCB8 transporter in nucleotide binding state. Biochemical and Biophysical Research Communications 2021; 557, 187-191.
Y Ichikawa, M Ghanefar, M Bayeva, R Wu, A Khechaduri, SV Naga Prasad, RK Mutharasan, TJ Naik and H Ardehali. Cardiotoxicity of doxorubicin is mediated through mitochondrial iron accumulation. The Journal of Clinical Investigation 2014; 124(2), 617-630.