Trends
Sci.
2026;
23(10):
13430
Drug Repurposing of Antihistamines as Antibiotic Adjuvants to Inhibit Biofilm Formation in Methicillin-Resistant Staphylococcus aureus: An In Silico and In Vitro Approach
Putri Nurfauziah1, Andi Rofian Sultan2, Helpia Helpia1 and Muhammad Aswad1,*
1Faculty of Pharmacy, Hasanuddin University, Makassar, South Sulawesi 90245, Indonesia
2Departement of Microbiology, Faculty of Medicine, Hasanuddin University, Makassar,
South Sulawesi 90245, Indonesia
(*Corresponding author’s e-mail: [email protected])
Received: 23 January 2026, Revised: 17 March 2026, Accepted: 24 March 2026, Published: 25 May 2026
Abstract
Methicillin-Resistant Staphylococcus aureus is an opportunistic pathogen that remains a serious threat to human health due to its high virulence, antibiotic resistance, and strong ability to form biofilms. Biofilm formation reduces the susceptibility of bacteria to conventional therapies and plays a critical role in persistent chronic infections. Drugs repurposing using antihistamines offers an efficient strategy to identify antibiotic adjuvants that can enhance antibiofilm efficacy. This study evaluates the potential of antihistamines, specifically fexofenadine, as an antibiotic adjuvant against biofilm-forming MRSA using integrated in silico and in vitro approaches. Molecular docking analysis targeting the biofilm regulator proteins AgrA (PDB ID: 4G4K) and SarA (PDB ID: 2FRH) showed that fexofenadine has high binding affinity, forming strong hydrogen and non-covalent interactions, and maintaining a stable complex during molecular dynamics simulations. In vitro checkerboard assay showed a synergistic effect between 0.5 µg/mL fexofenadine and 0.5 µg/mL cefoxitin, reducing planktonic bacterial growth by approximately 70% compared to the control. In the biofilm assay, the combination of fexofenadine 1 µg/mL and cefoxitin 16 µg/mL significantly inhibited biofilm formation by 70%, as indicated by lighter crystal violet staining. The in-silico results suggest that fexofenadine may interact with the biofilm regulatory proteins AgrA and SarA, which are anticipated to be involved in the quorum sensing system of Staphylococus aureus, while in vitro findings confirm its synergistic antibiofilm activity with cefoxitin. Overall, this study highlights the potential repurposing of antihistamines, particularly fexofenadine as an antibiofilm combination strategy to enhance therapeutic efficacy against antibiotic-resistant S.aureus infections.
Keywords: Staphlococcus aureus, Antibiofilm, Drug repurposing, Antihistamines, In silico, In vitro, Fexofenadine
Introduction
Staphylococcus aureus is a Gram-positive bacterium that is opportunistic and known to cause various difficult-to-treat infections in humans. Infections caused by S.aureus range from mild skin infections to more serious conditions such as bacteremia and endocarditis [1], S.aureus is a prevalent opportunistic pathogen in nosocomial infections, including those associated with methicillin-resistant strains (MRSA) [2,3].
S. aureus uses oligopeptides as signaling molecules, which play a crucial role in the secretion of virulence factors and biofilm formation, particularly within host tissues [4,5]. Quorum sensing (QS) regulation in S. aureus is controlled by the Sar (Staphylococcal Accessory Regulator) and the Agr system (Accessory Gene Regulator), which are crucial in this pathway, influencing various aspects of virulence and biofilm synthesis. These systems utilize oligopeptides to modulate the expression of secreted virulence factors [6,7]. The susceptibility of S.aureus to produce multilayered biofilms, which play a crucial role in the synthesis of the extracellular matrix that provides structural support for the biofilm bacteria, is significant. This matrix is resistant to antibiotic treatment and helps the bacteria evade the host’s immune response, thus improving bacterial pathogenicity [8,9].
Of the various mechanisms underlying biofilm formation, the QS system plays a crucial role. The QS system in S. aureus involves 2 main regulators AgrA and SarA, which regulate the expression of virulence genes and the bacteria’s ability to survive in biofilm form [6,10]. Inhibition of the SarA gene observed a significant effect on biofilm formation, with SarA mutants exhibiting a significant decrease in biofilm accumulation compared to the parent strain [7,11]. While AgrA plays a role in regulating toxin and adhesin genes, SarA also functions as a crucial global transcription regulator in the transition from the planktonic to the biofilm phase, interacting with various adhesion and matrix factors [12,13].
The presence of biofilms poses a greater challenge in treating infections caused by S. aureus, as bacteria in biofilm form are more resistant to antibiotic therapy than in their planktonic form. This is due to the protective matrix surrounding the bacterial cells, which acts as a physical barrier against antibiotics, as well as changes in the metabolism of the bacterial cells themselves [14,15]. Therefore, new therapeutic approaches are crucial. Drug repurposing, which involves reusing existing drugs for new purposes, has emerged as a promising strategy for targeting key proteins like AgrA and SarA to disrupt biofilm formation mechanisms and improve the efficacy of therapies.[16]. Overall, a deep understanding of biofilm mechanisms, particularly those involved in quorum sensing is key to developing effective therapeutic approaches against infections caused by S. aureus. Drug repurposing that can target these proteins is expected to provide new solutions to the challenges in treating bacterial infections [17].
The increasing prevalence of antibiotic resistance particularly in S.aureus strains, demands innovative therapeutic strategies to effectively combat infections. One promising approach is drug repurposing, which focuses on using existing medications for new therapeutic purposes, which speeds up the discovery of effective treatments. One of these antihistamines, shows potential as an anti-biofilm adjuvant antibiotic-resistant S.Aureus [18]. First-generation antihistamines, such as diphenhydramine, are known for their sedative properties due to their ability to easily cross the blood-brain barrier, while second-generation agents like cetirizine, desloratadine, and fexofenadine are recognized for fewer side effects and a better safety profile [18].
In this case, loratadine has been shown to inhibit the activity of the regulatory PASTA enzyme in S. aureus, significantly reducing biofilm formation while enhancing the effectiveness of β-lactam and vancomycin antibiotics against resistant strains [19]. Additionally, clemastine has shown the ability to disrupt biofilm integrity by interacting with the GdpP protein, which regulates cyclic AMP levels involved in biofilm regulation, thus improving the effectiveness of conventional antibiotics [20]. Then, there is evidence showing that antihistamines can enhance the antibacterial effectiveness against pathogens like Escherichia coli by disrupting bacterial quorum sensing systems, which are essential for biofilm formation, emphasizing that the structural features of antihistamines, particularly the presence of tertiary amino groups and lipophilic aromatic rings, play a significant role in their bioactive profile because they can provide surfactant characteristics that improve penetration into bacterial membranes and affect cell communication pathways, including QS [21].
However, studies integrating in silico molecular docking of antihistamines targeting quorum sensing regulators such as AgrA and SarA with in vitro validation of their potential remain limited. Therefore, this study to evaluate the potential of selected antihistamines against antibiotic-resistant S.Aureus through computational screening of AgrA and SarA, followed by in vitro assessment using checkerboard and biofilm inhibition assays on the most promising compound. This approach aims to provide new insights into antihistamine repurposing as a combination strategy to enhance the efficacy of conventional antibiotic therapy.
Materials and methods
Ligand preparation
The
3D structures of antihistamine drugs were downloaded from PubChem
(https://pubchem.ncbi.nl
m.nih.gov/).
The UCSF Chimera software was employed to convert these structures
into PDB files, followed by geometry optimization and energy
minimization using the built-in minimization tools to obtain the
most stable ligand conformations.
Protein preparation
The crystal structures of the target proteins (PDB ID: 4G4K, 2FRH) were obtained from the Protein Data Bank (https://www.rcsb.org/). Prior to docking, the protein was preprocessed by removing water molecules, adding hydrogen atoms, and assigning charges using AutoDock Vina® integrated within UCSF Chimera. The processed protein file was then saved in PDBQT format. Docking results and ligand-protein interaction visualizations were analyzed using Discovery Studio.
Molecular docking
Molecular docking was performed using UCSF-Chimera connected to AutoDock Vina, starting with ligand and protein preparation (dock prep), followed by the docking process using a grid box in AutoDock Vina. The highest binding energy and largest cluster population were used as criteria to select the best-docked conformation [22]. The 2D diagrams of the interactions between proteins and each compound. 3D visualization and analysis were performed with Discovery Studio [6].
Molecular dynamic
Molecular dynamics simulations were performed on antihistamine drugs that had greater binding affinity and more bonds formed. Molecular dynamics simulations were performed using YASARA (Yet Another Scientific Artificial Reality Application) software with the AMBER14 force field [23]. The simulation conditions were set to match physiological conditions (310K, pH 7.4, 1 atm, and 0.9% salt concentration) for 100 ns. The macro programs used include MD_run for simulation and MD_analyze for evaluating Root Mean Square Fluctuation (RMSF), Radius of Gyration (Rg), and Root Mean Square Deviation (RMSD).
Materials in vitro
Pure cultures of Methicillin-Resistant Staphylococcus aureus (MRSA) bacteria were obtained from the collection of Hasanuddin University Hospital, 70% alcohol, sterile water, MHB (Mueller-Hinton Broth) media, MSA (Mannitol Salt Agar) media, TSB (Trytic Soy Broth) media, 0.9% NaCl, Dimethylsulfoxide (DMSO), and pure fexofenadine drug from BPOM (Indonesian Agency of Drug nad Food Control).
Preparation of test bacteria
The bacterial strains used in this study consisted of Methicillin-resistant Staphylococcus aureus (MRSA) bacterial strain were stored at 4 °C and maintained through subculturing on agar media prior to the incubation process. MRSA was cultured on Mannitol Salt Agar (MSA), with incubation at 37 °C for 24 h. After incubation, a bacterial suspension was prepared by adding 0.9% NaCl to reach a concentration equivalent to a 0.5 McFarland standard, or approximately 1.5×10⁸ CFU/mL.
Preparation of test solutions
Stock solutions of 6,400 μg/mL fexofenadine and the antibiotic cefoxitin were prepared by dissolving 6.4 mg of the drug in 1 mL of DMSO. DMSO has no impact on bacterial growth or biofilm formation [20].
Preparation of media
The culture media were prepared according to standard microbiological procedures. Specifically, 6 g of Tryptic Soy Broth (TSB), 9.9 g of MacConkey Agar (MCA), and 11.1 g of Mannitol Salt Agar (MSA) were each dissolved in 200 mL of distilled water, while 3.4 g of Mueller-Hinton Agar (MHA) was dissolved in 100 mL of distilled water. All media were heated until completely dissolved and subsequently sterilized by autoclaving at 121 °C for 15 min to ensure aseptic conditions prior to use.
MIC checkerboard assay
A 0.5 McFarland MRSA suspension diluted 1:100 in MHB was used to prepare serial dilutions of fexofenadine (32, 16, 8, 4, 2 and 0 µg/mL) in U-bottom microplates using the 2-fold dilution method, followed by incubation at 37 °C for 15 - 20 min. Similar procedures were applied to cefoxitin without bacteria in separate microplates. The contents of the 2 microplates were combined by adding 100 µL of antibiotic (horizontally) and 100 µL of fexofenadine and bacteria (vertically), so that each well contained a combination of both. The final volume is 200 µL, with a bacterial concentration of 7.5×10⁵ CFU/mL. Incubation was performed at 37 °C for 18 - 20 h, OD was measured at 620 nm, and the results were analyzed using Combenefit to assess synergistic or antagonistic effects [24,25].
Antibiofilm assay
MRSA suspensions with a concentration of 0.5 McFarland diluted 1:1000 in TSB were used for serial dilutions of fexofenadine (32, 16, 8, 4, 2 and 0 µg/mL) in flat-bottom microplates dilutions, followed by incubation at 37 °C for 15 - 20 min. A similar procedure was performed for cefoxitin without bacteria in different microplates. The 2 microplates are combined: 100 µL of antibiotic (horizontal) and 100 µL of fexofenadine and bacteria (vertical), resulting in a concentration gradient. The final volume was 200 µL, with a bacterial concentration of 7.5×104 CFU/mL, incubated at 37 °C for 18 - 20 h with rotation at 150 rpm [26]. After that, the microplate was washed with sterile water, stained with 1.2% crystal violet for 5 minutes, rinsed 5 times with sterile water, dried, then 200 µL of solution (40% ethanol, 10% acetic acid, 50% sterile water) was added, and incubated at 37°C for 15 minutes at 150 rpm in an incubator shaker. OD was measured at a wavelength of 620 nm using an ELISA reader [27].
Statistical analysis
All assays were performed at least 3 times, and the results are presented as mean ± standard deviation (SD). Statistical analysis was performed using GraphPad Prism version 10.5. Differences among treatment groups were evaluated using ordinary one-way analysis of variance (ANOVA) followed by Dunnett’s multiple comparison test to compare each treatment group with the control. All results are expressed as mean ± standard deviation (SD). A p-value < 0.05 was considered statistically significant.
Results and discussion
AgrA and SarA are key regulators in the formation of Staphylococcus aureus biofilms through the control of the quorum sensing (QS) system and the expression of virulence factors. SarA increases the expression of the ica operon, which is essential for the production of extracellular polymeric substance (EPS) matrix, while AgrA regulates biofilm dispersal and maturation from the modulation of adhesion factors and proteolytic enzymes [28,29]. Inhibiting these 2 regulators disrupts QS signaling, reduces EPS and eDNA synthesis, which reduces biofilm structure without inhibiting bacterial growth [30,31]. This approach provides potential therapeutic strategies to combat persistent infections and resistant strains by minimizing the colonization and virulence of S. aureus. Previous studies have shown that natural molecules such as quercetin and 3-hydroxybenzoic acid are able to bind stably to AgrA and SarA, triggering protein conformational changes and reducing the expression of biofilm-related genes, making them ideal targets for antibiofilm strategies. This molecular docking approach has the potential to reduce the bacteria’s ability to form biofilms, thereby increasing the effectiveness of antibiotic therapy [32]. In this study, the target protein SarA with the code (PDB id: 2FRH) and AgrA with the code (PDB id: 4G4K) were used for biofilm formation inhibition.
The molecular docking results from 10 tested first-generation and second-generation antihistamines showed that the best 3 were: Cetirizine, desloratadine, and fexofenadine. A lower affinity value indicates a stronger and more stable interaction. As for the visualization of amino acid residues, the parameters analyzed include hydrogen bonds, as well as non-covalent interaction types such as Van der Waals, Pi-Sigma, Pi-Alkyl, and Pi-Pi stacking with the amino acid residues at the active site. Antihistamine drugs that show the strongest binding affinity values include cetirizine −5.5 kcal/mol, desloratadine −6.2 kcal/mol, and fexofenadine −6.2 kcal/mol. These drugs interact with SarA thru hydrogen bonds at Thr162 for cetirizine, and similarly, desloratadine interacts thru Pi-Alkyl bonds at Leu160 and hydrogen bonds at Asn146. Fexofenadine interacts thru pi-sigma bonds at His159, pi-donor bonds at Tyr162, and hydrogen bonds at Asn146. This is compared to the antibiotic cefoxitin −5.1 kcal/mol, which forms interactions thru hydrogen bonds at Asn161, Thr141, and His159. Meanwhile, the strongest binding affinity values at 4g4k are for cetirizine −6.2 kcal/mol, desloratadine −6.7 kcal/mol, and fexofenadine −6.9 kcal/mol. These interact with AgrA thru hydrogen bonds Tyr183, Gly184 and Pi-Alkyl interactions Leu192 for cetirizine. Similarly, desloratadine interacts thru Pi-Alkyl bonds Leu192, Ile143, while fexofenadine interacts thru hydrogen bonds Glu144, amide-Pi stacking Tyr183, and alkyl bonds Leu192, Leu189, and van der Waals interactions Gly184. Compared to the antibiotic cefoxitin -5.6 kcal/mol, which forms interactions thru hydrogen bonds with Glu144 and Glu188, thru carbon-hydrogen bonds with Tyr183 and Glu184, and thru Pi-alkyl bonds with Lys146. These results indicate the potential of antihistamines as candidate biofilm inhibitors by targeting SarA and AgrA, with lower affinity than cefoxitin. Table 1 presents the molecular docking results, indicating the antibiofilm potential of the tested antihistamine.
Table 1 Ligand docking results with target proteins 2FRH and 4G4K.
No |
Drugs |
2FRH |
4G4K |
|
|||||||
Binding Afinity |
H. Bond |
Interacting residues |
Binding Afinity |
H.Bond |
Interacting residues |
|
|||||
AH1 generasi 1 |
|
||||||||||
1 |
Brompheniramine |
−4.9 |
-
|
Pi-Alkyl: Leu160 |
−6.3 |
-
|
Pi-Alkyl: Leu189
|
|
|||
2 |
Chlorpheniramine |
−4.8 |
- |
- |
−6.4 |
- |
Pi-Alkyl: Leu189
|
|
|||
3 |
Dimethindene |
−5.0 |
- |
Pi-Alkyl: Asn146, Leu160
|
−6.0 |
- |
Pi-Alkyl: Leu189
|
|
|||
4 |
Buclizine |
−5.3 |
- |
Carbon, Pi-hydrogen bone: His159, Tyr162
|
−6.2 |
-
|
Pi-Alkyl: Leu192 Pi-Donor Hydrogen Bond: Glu144 Carbon hydrogen bond: Tyr183 Amide pi stacked: Gly184 |
|
|||
5 |
Diphenhydramine |
−4.7 |
- |
Pi-Alkyl: Tyr 142
|
−5.3 |
-
|
Carbon hydrogen bond: Glu144
|
|
|||
AH1 generasi 2 |
|
||||||||||
6 |
Acrivastine |
−5.5 |
Thr141 |
Pi-Sigma: Tyr162 Hydrogen bond: Thr141 |
−6.2 |
Thr142 |
Pi-Alkyl: Leu192
|
|
|||
7 |
Cetirizine |
−5.5 |
Tyr162 |
Hydrogen bond: Tyr162 |
−6.2 |
Tyr183, Gly184, his169 |
Hydrogen bond: Tyr183, Gly184 Pi-Alkyl: Leu192 |
|
|||
8 |
Desloratadine |
−6.2 |
Asn146 |
Pi-Alkyl: Leu160 Hydrogen bond: Asn 146
|
−6.7 |
- |
Pi-Alkyl: Leu192, Ile143
|
|
|||
9 |
Olopatadine |
−5.4 |
Thr141 |
Hydrogen bond: Tyr 141
|
−5.9
|
Tyr183
|
Hydrogen bond: Tyr183 Pi-Alkyl: Leu192, Ile143 Carbon Hydrogen Bond: Tyr183 |
|
|||
10 |
Fexofenadine |
−6.2 |
Lys123, Asn146 |
Pi-Sigma: His159 Pi-Donor: Tyr162 Hydrogen bond: Asn 146 |
−6.9
|
Glu144 |
Hydrogen bond: Glu144 Amide-Pi Stacked: Tyr183 Alkyl bound: Leu192, Leu189, Lys146 Van der waals: Gly184 |
|
|||
Antibiotic |
|
||||||||||
|
Cefoxitin |
−5.1 |
Asn161, TYR162, Thr141, His159 |
Hydrogen bond: Asn161, Thr14, His159 Pi alkyk: Leu160 |
−5.6 |
Glu144, Asn185, Glu188 |
Hydrogen bond: Glu144, Glu188 Carbon bond: Tyr183, Gly184 Alkyl bound: Leu192 |
||||
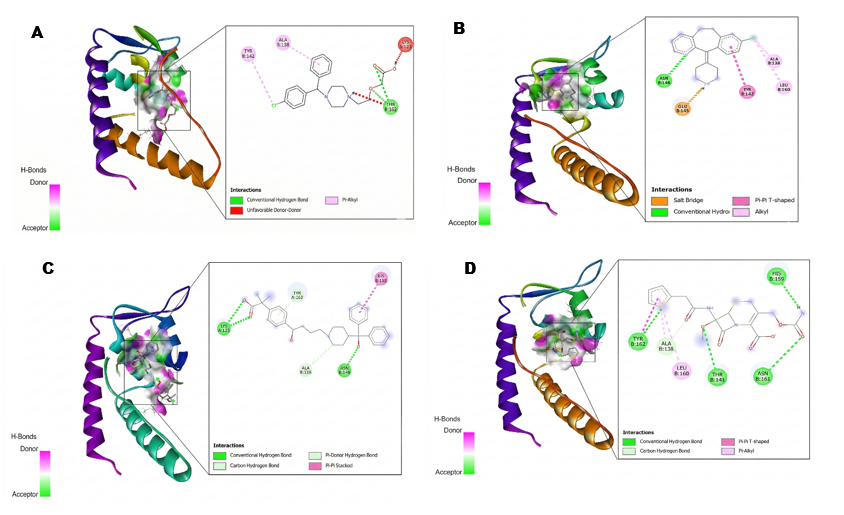
Figure 1 Two-dimensional and 3-dimensional diagrammatic representation of the interaction of the best antihistamine drug docking with cefoxitin against the 2FRH enzyme (A) Cetirizine, (B) Desloratadine, (C)Fexofenadine, and (D) Cefoxitin.
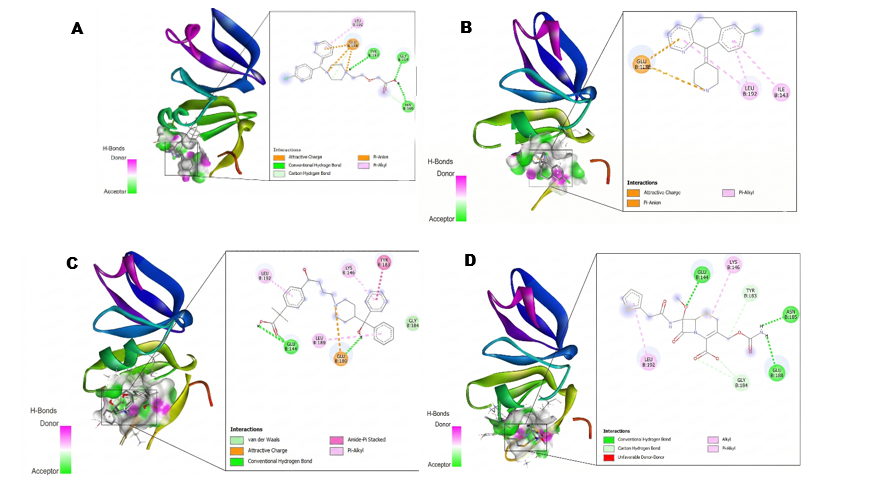
Figure 2 Two-dimensional and 3-dimensional diagrammatic representation of the best antihistamine drug docking interaction with cefoxitin against the 4G4K enzyme (A) Cetirizine, (B) Desloratadine, (C)Fexofenadine, and (D) Cefoxitin.
Molecular dynamic analysis
Molecular dynamics (MD) simulations play a crucial role in validating protein-ligand complexes generated from docking, providing deep insights into the stability and interactions of these complexes thru key parameters such as Radius of Gyration (RoG), Root Mean Square Deviation (RMSD), and Root Mean Square Fluctuation (RMSF) [33,34]. RoG is used to assess the distribution of atoms along the protein’s main axis, reflecting changes in the size and stability of the protein fold [35]. RMSD measures the deviation of complex structures from their initial form, with values below 5 Å indicating structural stability during the simulation [34]. Meanwhile, RMSF evaluates the fluctuation of amino acid residues to assess local flexibility and potential conformational changes around the binding site [36].
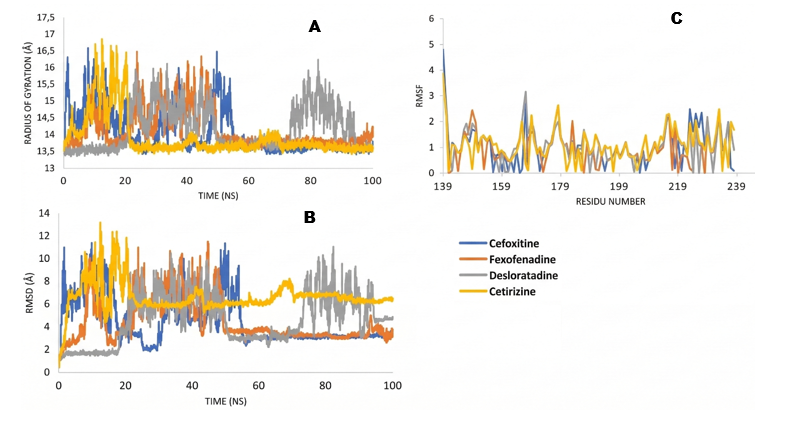
Figure 3 Molecular dynamics diagram in protein SaRA; (A) Radius of Gyration, (B) RMSD, and (C) RMSF.
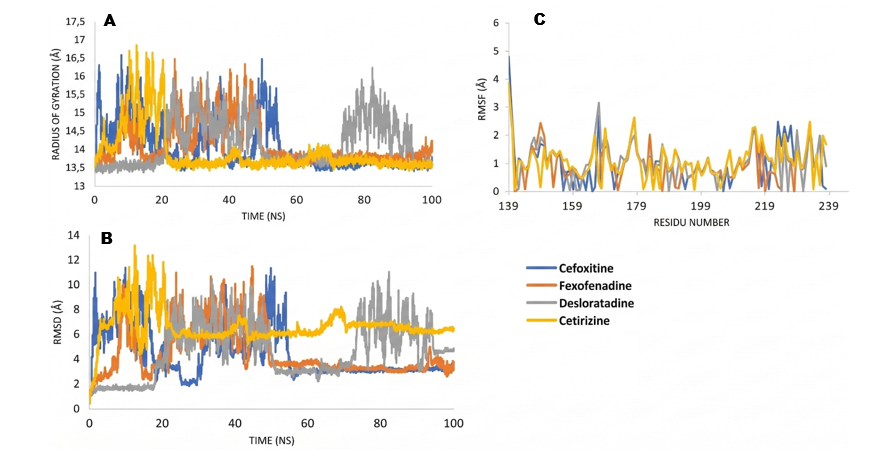
Figure 4 Molecular dynamics diagram in protein AgrA; (A) Radius of Gyration, (B) RMSD, and (C) RMSF.
In Figure 3, the radius of gyration of the SarA protein shows that fexofenadine undergoes 2 size changes during the simulation (peaking at 17.5 Å at 25 ns, dropping to 16 Å at 50 ns), while cefoxitin fluctuates up to 18 Å at 18 ns. RMSD shows that fexofenadine is the most stable (< 5 Å), while cetirizine and desloratadine reach 6 Å and 11 Å, and cefoxitin 7 Å. RMSF shows that fexofenadine exhibits fluctuations at Asn107 and Ile103, which are not active residues. Overall, fexofenadine has better affinity and stability compared to cefoxitin. In 4G4K, in image 4, fexofenadine shows a change in Radius of Gyration with a peak of 16.5 Å at 49 ns, decreasing to 14 Å at 50 ns, while cefoxitin reaches 17 Å at 53 ns. RMSD shows that fexofenadine reaches stability < 5 Å faster at 43 ns compared to cefoxitin at 58 ns. RMSF indicates that cefoxitin is more stable, while fexofenadine shows fluctuations at Arg233 and Val232, which are not active residues. Overall, fexofenadine showed better affinity and stability compared to cefoxitin.
MIC checkerboard assay
The Loewe Combenefit model is primarily used to analyze interactions in checkerboard assay designs involving various dose responses. However, it is important to recognize that this model may not be suitable for assessing drug interactions derived from single-dose experiments [37]. Therefore, a systematic 2-fold serial dilution approach was used to measure the combined effects of various concentrations in a well-controlled manner, ultimately helping to understand the potential for synergistic or antagonistic drug interactions. Data collected from the checkerboard assay uses the visible turbidity of optical density (OD) as a reading [38]. This test was conducted to determine the potential of the antihistamine drug when combined with antibiotics against the bacterium S.Aureus. The results of the fexofenadine checkerboard assay provided synergistic results that were consistent with the in-silico findings. That is, the combination of cefoxitin with fexofenadine was found to increase the antibiotic activity of cefoxitin. The single administration of cefoxitin 0.5 µg/mL, resulted in bacterial growth of about 80% relative to the control (bacterial culture in medium without drug treatment). In contrast, the combination of cefoxitin 0.5 µg/mL and fexofenadine 0.5 µg/mL resulted in bacterial growth of about 10% relative to the control, thus indicating a reduction of about 70%. These results were supported by statistical analysis showing significant differences among treatment groups using 1-way ANOVA (p < 0.0001, as shown in Figure 5.
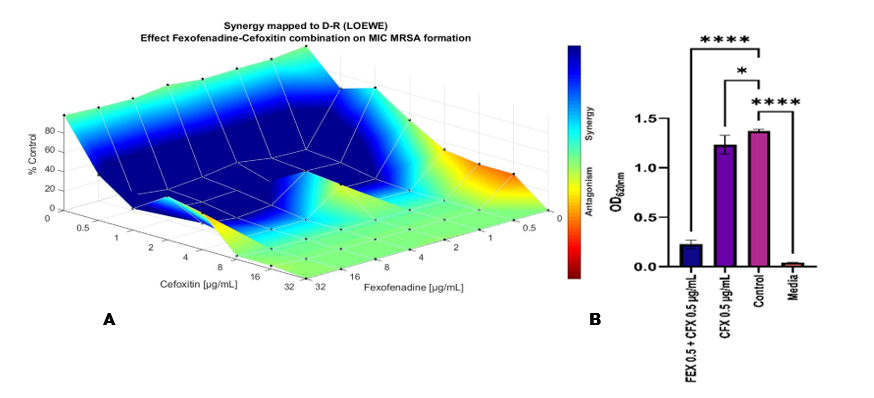
Figure 5 (A) Results of the checkerboard assay analysis combining fexofenadine with cefoxitin using Combenefit software and the LOEWE model, (B) Data are presented as mean ± standard deviation (SD) from 3 independent experiments (n = 3). Statistical significance among groups was determined using 1-way ANOVA. Significant differences are indicated as p < 0.05 (*) and p < 0.0001 (***). Error bars represent standard deviations. FEX = fexofenadine; CFX = cefoxitin.
Biofilms are formations of microbial cells incased in an extracellular polymeric substance (EPS) matrix. This biofilm structure provides protection against the host’s immune system and antibiotics. Biofilms can increase bacterial resistance by up to 1,000 times compared to bacteria in planktonic form, due to limited antibiotic penetration into the matrix [39]. Crystal violet staining is used to measure biofilm formation by bacteria. The varying intensity of purple colour in each well indicates a variation in the amount of biofilm formed. Wells with a darker purple colour indicate more biofilm formation, while wells with a lighter purple or almost colourless appearance indicate less or no biofilm formation [40,41]. However, to clarify further, a reading OD value of 620 nm was taken on an ELISA reader to reflect biofilm density as a function of the amount of dye bound to the biofilm matrix, which is composed of polysaccharides, nucleic acids, and proteins [42]. The percentage of biofilm growth inhibition was calculated using the following equation [43]:
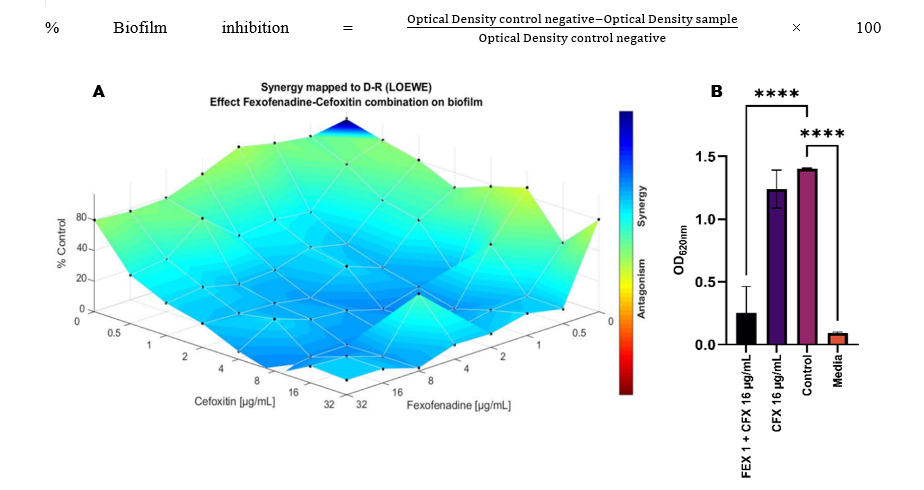
Figure 6 (A) Results of the biofilm analysis combining fexofenadine with cefoxitin using Combenefit software and the LOEWE model, (B) Data are presented as mean ± standard deviation (SD) from 3 independent experiments (n = 3). Statistical significance among groups was determined using 1-way ANOVA. Significant differences are indicated as p < 0.0001 (****). Error bars represent standard deviations. FEX = fexofenadine; CFX = cefoxitin.
After calculating the percentage of biofilm inhibition, the results were then entered into the Combenefit software, as shown in the Figure 6. In this combenefit analysis, it was found that the combination of cefoxitin with fexofenadine enhances the antibiotic activity of cefoxitin. Cefoxitin at a single concentration of 16 µg/mL, when administered alone, residual biofilm biomass relative about 80% relative to the control (bacterial culture in medium without drug treatment). In contrast, the combination of cefoxitin 18 µg/mL and fexofenadine 1 µg/mL generated residual biofilm biomass relative about 10% relative to the control, thus indicating a reduction of about 70%. These findings were further supported by statistical analysis showing significant differences among treatment groups using 1-way ANOVA (p < 0.0001).
Recent studies have increasingly highlighted the potential of repurposing non-antibiotic drugs as antimicrobial adjuvants to enhance the efficacy of conventional antibiotics against resistant pathogens. Phenothiazine antipsychotics, such as chlorpromazine and thioridazine, can inhibit bacterial efflux pumps NorA, disrupt membrane potential, and interfere with cell division, thereby increasing intracellular antibiotic accumulation [44,45]. Similarly, Selective Serotonin Reuptake Inhibitor (SSRI) antidepressants, such as sertraline, have been reported to modulate multidrug resistance-related genes including marA, acrA, and acrB. These mechanisms may also contribute to biofilm inhibition, as disruption of efflux activity and membrane integrity can impair bacterial communication and biofilm formation [45,46]. These findings underscore the growing interest in non-antibiotic drug repurposing as a complementary strategy to combat antimicrobial resistance. In this context, antihistamines have also been reported to exhibit potential as antibiotic adjuvants against S. Aureus. For instance, loratadine has been shown to inhibit regulatory pathways involved in cell wall synthesis, thereby reducing biofilm formation and enhancing the activity of β-lactam antibiotics [19], while terfenadine has also demonstrated notable antibiofilm activity against S. aureus [47]. Similarly, fexofenadine showed synergistic activity when combined with cefoxitin, significantly reducing planktonic bacterial growth by approximately 10% compared to the control. In the checkerboard assay, the combination of cefoxitin 0.5 µg/mL and fexofenadine 0.5 µg/mL having the strongest antibacterial effect, while higher concentrations fexofenadine 1 µg/mL and cefoxitin 16 µg/m were required to inhibit biofilm formation, as indicated by lighter crystal violet staining. These results suggest that the cefoxitin and fexofenadine combination is more effective against planktonic cells, as the biofilm matrix limits drug penetration, the need for optimized dosing or adjunctive agents to enhance antibiofilm efficacy.
Although the present findings were obtained from in vitro experiments, the observed antibiofilm activity of fexofenadine may have potential translational relevance. Fexofenadine is a second-generation antihistamine widely used in clinical practice with an established safety profile, typically administered at doses of 60 - 180 mg per day [48]. Pharmacokinetic studies indicate that peak plasma concentrations (Cmax) of fexofenadine typically range from approximately 0.2 to 1.12 µg/mL following standard therapeutic dosing. The concentration used in the checkerboard assay (0.5 µg/mL) falls within this clinically relevant range, while the concentration applied in the biofilm assay (1 µg/mL) remains close to the upper range of reported plasma levels [48,49]. Given the relatively low oral bioavailability of fexofenadine (approximately 33% - 35%) and its minimal metabolism, local tissue concentrations may vary from systemic plasma levels, which should be considered when interpreting the in vitro findings [49]. Moreover, several studies have demonstrated that non-antibiotic drugs can modulate bacterial virulence and biofilm formation at sub-inhibitory concentrations without exerting strong bactericidal pressure, potentially reducing the risk of resistance development while enhancing antibiotic efficacy. Nevertheless, further pharmacokinetic and pharmacodynamic studies are required to determine the clinical feasibility of antihistamines as antibiotic adjuvants against biofilm-associated infections and gene expression analysis of biofilm regulatory genes are required to better determine the therapeutic feasibility of antihistamines as antibiotic adjuvants.
Conclusions
The combination of fexofenadine with cefoxitin showed an antibiofilm effect against S.Aureus in vitro tests. At a concentration of 0.5 µg/mL fexofenadine and 0.5 µg/mL cefoxitin, the combination synergistically enhances the antibacterial activity of cefoxitin by reducing planktonic bacterial growth by up to 10% compared to the control. At higher concentrations (1 µg/mL fexofenadine and 16 µg/mL cefoxitin), this combination effectively inhibits biofilm formation, as indicated by lighter crystal violet staining. These results are consistent with in silico findings, where fexofenadine exhibits strong binding affinity and stable interactions with the biofilm regulator proteins AgrA and SarA, suggesting a potential interaction that may influence biofilm formation. This study suggests that repurposed antihistamines, particularly fexofenadine, may have potential as an antibiotic adjuvant in combination therapy to enhance antibiofilm activity against methicillin-resistant Staphylococcus aureus. This finding emphasizes the importance of optimizing concentration and formulation strategies to maximize its clinical application in controlling resistant S.Aureus infections.
Acknowledgements
The authors gratefully acknowledge the support of the Clinical Microbiology Laboratory, Hasanuddin University, Indonesia, during the experimental work. The authors also thank to Hasanuddin University, Indonesia, for Thematic Research Group Funding 2025.
Declaration of Generative AI in Scientific Writing
The authors declare that this manuscript was written and revised by the author themselves. AI tools were used only to improve the grammar, clarity, and language quality of the text.
CRediT Author Statement
Putri Nurfauziah: Writing Original draft, Formal analysis, Resources, Visualization, Helpia Helpia: Investigation, Project administration. Andi Rofian Sultan: Conceptualization, Supervision, Methodology and Writing - Review & Editing. Muhammad Aswad: Conceptualization, Methodology, Supervision, and Writing - Review & Editing.
References
[1] SS Alnejadi and AH Saleh. Gene expression of carbapenemase-encoding genes in multi-drug resistant Staphylococcus aureus from clinical sources. European Journal of Medical Genetics and Clinical Biology 2024; 1(5), 179-186.
[2] BA Clent, Y Wang, HC Britton, F Otto, CJ Swain, MH Todd and MH Todd. Molecular docking with open access software: Development of an online laboratory handbook and remote workflow for chemistry and pharmacy master’s students to undertake computer-aided drug design. Journal of Chemical Education 2021; 98(9), 2899-2905.
[3] HR Nasution, AW Septama, T Ernawati, NA Khairunnisa, SE Nugraha and DS Utami. Ethanolic extract of Curcuma domestica Val. and Curcuma xanthorrhiza Roxb.: A comparative study in vitro and in silico antibacterial effect against methicillin-resistant Staphylococcus aureus. Trends in Sciences 2025; 22(5), 9458.
[4] B Kim, HJ Lee, SH Jo, MG Kim, Y Lee, W Lee, W Kim, HS Joo, YG Kim, JS Kim and YH Yang. Study of SarA by DNA affinity capture assay (DACA) employing 3 promoters of key virulence and resistance genes in methicillin-resistant Staphylococcus aureus. Antibiotics 2022; 11(12), 1714.
[5] L Li, A Cheung, AS Bayer, L Chen, W Abdelhady, BN Kreiswirth, MR Yeaman and YQ Xiong. The global regulon sarA regulates beta-lactam antibiotic resistance in methicillin-resistant Staphylococcus aureus in vitro and in endovascular infections. The Journal of Infectious Diseases 2016; 214(9), 1421-1429.
[6] MA Diaz, EG Vega-Hissi, MA Blazquez, MR Alberto and ME Arena. Restraining Staphylococcus aureus virulence factors and quorum sensing through lactic acid bacteria supernatant extracts. Antibiotics 2024; 13(4), 297.
[7] P Liu, X Kang, CH Chen, X Luo, C Li and G Wang. Quercetin targets SarA of methicillin-resistant Staphylococcus aureus to mitigate biofilm formation. Microbiology Spectrum 2024; 12(1), e02722-23.
[8] H Sang, H Jin, P Song, W Xu and F Wang. Gallic acid exerts antibiofilm activity by inhibiting methicillin-resistant Staphylococcus aureus adhesion. Scientific Reports 2024; 14(1), 17220.
[9] W Abdelraheem, S Abdelraheem and S Zaki. Phenotypic and genotypic detection of biofilm formation and methicillin resistance among Staphylococcus aureus isolates. Microbes and Infectious Diseases 2021; 2(3), 485-496.
[10] G Wang, L Li, X Wang, X Li, Y Zhang, J Yu, J Jiang, X You and YQ Xiong. Hypericin enhances beta-lactam antibiotics activity by inhibiting sarA expression in methicillin-resistant Staphylococcus aureus. Acta Pharmaceutica Sinica B 2019; 9(6), 1174-1182.
[11] W Abdelhady, AS Bayer, K Seidl, DE Moormeier, KW Bayles, A Cheung, MR Yeaman and YQ Xiong. Impact of vancomycin on sarA-mediated biofilm formation: Role in persistent endovascular infections due to methicillin-resistant Staphylococcus aureus. The Journal of Infectious Diseases 2014; 209(8), 1231-1240.
[12] Y Li, S Li, K Yang, R Guo, X Zhu, Y Shi and A Huang. Antibiofilm mechanism of a novel milk-derived antimicrobial peptide against Staphylococcus aureus by downregulating agr quorum sensing system. Journal of Applied Microbiology 2022; 133(4), 2198-2209.
[13] J Qin, L Yu, F Peng, X Ye, G Li, C Sun, F Cheng, C Peng and X Xie. Tannin extracted from Penthorum chinense Pursh, a potential drug with antimicrobial and antibiofilm effects against methicillin-sensitive Staphylococcus aureus and methicillin-resistant Staphylococcus aureus. Frontiers in Microbiology 2023; 14, 1134207.
[14] FM Ballah, MS Islam, ML Rana, MA Ullah, FB Ferdous, FH Neloy, S Ievy, MA Sobur, AMMT Rahman, MM Khatun, M Rahman and MT Rahman. Virulence determinants and methicillin resistance in biofilm-forming Staphylococcus aureus from various food sources in Bangladesh. Antibiotics 2022; 11(11), 1666.
[15] K Azmi, W Qrei and Z Abdeen. Screening of genes encoding adhesion factors and biofilm production in methicillin resistant strains of Staphylococcus aureus isolated from Palestinian patients. BMC Genomics 2019; 20(1), 578.
[16] AH Jonker, D O’Connor, MM Cavaller-Bellaubi, C Fetro, M Gogou, PAC T’Hoen, M de Kort, H Stone, N Valentine and AMG Pasmooij. Drug repurposing for rare: Progress and opportunities for the rare disease community. Frontiers in Medicine 2024; 11, 1352803.
[17] H Kawauchi, K Yanai, DY Wang, K Itahashi and K Okubo. Antihistamines for allergic rhinitis treatment from the viewpoint of nonsedative properties. International Journal of Molecular Sciences 2019; 20(1), 213.
[18] A Schafer, H Cheng, R Xiong, V Soloveva, C Retterer, F Mo, S Bavari, G Thatcher and L Rong. Repurposing potential of 1st generation H1-specific antihistamines as anti-filovirus therapeutics. Antiviral Research 2018; 157, 47-56.
[19] N Cutrona, K Gillard, R Ulrich, M Seemann, HB Miller and MS Blackledge. From antihistamine to anti-infective: Loratadine inhibition of regulatory PASTA kinases in staphylococci reduces biofilm formation and potentiates beta-lactam antibiotics and vancomycin in resistant strains of Staphylococcus aureus. ACS Infectious Diseases 2019; 5(8), 1397-1410.
[20] Y Shang, J Guo, Y Zhao, J Chen, Q Meng, D Qu, J Zheng, Z Yu, Y Wu and Q Deng. Clemastine inhibits the biofilm and hemolytic of Staphylococcus aureus through the GdpP protein. Microbiology Spectrum 2022; 10(2), e0054121.
[21] GG Bruer, P Hagedorn, M Kietzmann, AF Tohamy, V Filor, E Schultz, S Mielke-Kuschow and J Meissner. Histamine H1 receptor antagonists enhance the efficacy of antibacterials against Escherichia coli. BMC Veterinary Research 2019; 15(1), 55.
[22] SS Butt, Y Badshah, M Shabbir and M Rafiq. Molecular docking using Chimera and AutoDock Vina software for nonbioinformaticians. JMIR Bioinformatics and Biotechnology 2020; 1(1), e14232.
[23] H Land and MS Humble. Chapter 4 in biocatalytic investigations. Biocatalytic Investigations 2018; 1685, 43-67.
[24] T Chipenzi, G Baloyi, T Mudondo, S Sithole, GF Chi and S Mukanganyama. An evaluation of the antibacterial properties of tormentic acid congener and extracts from Callistemon viminalis on selected ESKAPE pathogens and effects on biofilm formation. Advances in Pharmacological and Pharmaceutical Sciences 2020; 2020, 8848606.
[25] P Aelenei, CM Rimbu, CE Horhogea, A Lobiuc, AN Neagu, SI Dunca, I Motrescu, G Dimitriu, AC Aprotosoaie and A Miron. Prenylated phenolics as promising candidates for combination antibacterial therapy: Morusin and kuwanon G. Saudi Pharmaceutical Journal 2020; 28(10), 1172-1181.
[26] ZX Luo, Y Li, MF Liu and R Zhao. Ciprofloxacin enhances the biofilm formation of Staphylococcus aureus via an agrC-dependent mechanism. Frontiers in Microbiology 2023; 14, 1328947.
[27] F Zhang, W Dong, F Wang, J Yu, F Jiang, J Tang, Y Qian and H Shen. The topical tranexamic acid have potential hazard of promoting biofilm formation of Staphylococcus aureus in microenvironment of the prosthetic joint. BioMed Research International 2021; 2021, 5748069.
[28] L Liu, X Shen, J Yu, X Cao, Q Zhan, Y Guo and F Yu. Subinhibitory concentrations of fusidic acid may reduce the virulence of S. aureus by down-regulating sarA and saeRS to reduce biofilm formation and alpha-toxin expression. Frontiers in Microbiology 2020; 11, 25.
[29] MM Saleh, N Yousef, SM Shafik and HA Abbas. Attenuating the virulence of the resistant superbug Staphylococcus aureus bacteria isolated from neonatal sepsis by ascorbic acid, dexamethasone, and sodium bicarbonate. BMC Microbiology 2022; 22(1), 268.
[30] A Valliammai, A Selvaraj, U Yuvashree, C Aravindraja and SK Pandian. sarA-dependent antibiofilm activity of thymol enhances the antibacterial efficacy of rifampicin against Staphylococcus aureus. Frontiers in Microbiology 2020; 11, 1744.
[31] MJ Campbell, KE Beenken, AM Ramirez and MS Smeltzer. Increased production of aureolysin and staphopain A is a primary determinant of the reduced virulence of Staphylococcus aureus sarA mutants in osteomyelitis. mBio 2024; 15(4), e0338323.
[32] PS Ganesh, K Veena, R Senthil, K Iswamy, EM Ponmalar, V Mariappan, AS Smiline Girija, J Vadivelu, S Nagarajan, D Challabathula and EM Shankar. Biofilm-associated Agr and Sar quorum sensing systems of Staphylococcus aureus are inhibited by 3-hydroxybenzoic acid derived from Illicium verum. ACS Omega 2022; 7(17), 14653-14665.
[33] D Sun, L Xu, M Tong, Z Wei, W Zhang, J Liang, X Liu and Y Wang. De novo design of mIDH1 inhibitors by integrating deep learning and molecular modeling. Frontiers in Pharmacology 2024; 15, 1491699.
[34] CA Sarri, GE Papadopoulos and Z Mamuris. West Nile virus-associated HLA-DRB1 alleles in the Greek population: A structural perspective. Microbiology and Immunology 2023; 67(3), 154-159.
[35] P Garg, VK Vanamamalai, I Jali and S Sharma. In silico prediction of the animal susceptibility and virtual screening of natural compounds against SARS-CoV-2: Molecular dynamics simulation based analysis. Frontiers in Genetics 2022; 13, 906955.
[36] ZB Zhang, YL Xia, GH Dong, YX Fu and SQ Liu. Exploring the cold-adaptation mechanism of serine hydroxymethyltransferase by comparative molecular dynamics simulations. International Journal of Molecular Sciences 2021; 22(4), 1781.
[37] RG Maset, A Hapeshi, J Lapage, N Harrington, J Littler, S Perrier and F Harrison. Combining SNAPs with antibiotics shows enhanced synergistic efficacy against S. aureus and P. aeruginosa biofilms. npj Biofilms and Microbiomes 2023; 9(1), 36.
[38] R Rajendran, C Williams, DF Lappin, O Millington, M Martins and G Ramage. Extracellular DNA release acts as an antifungal resistance mechanism in mature Aspergillus fumigatus biofilms. Eukaryotic Cell 2013; 12(3), 420-429.
[39] T Heinonen, S Hargraves, M Georgieva, C Widmann and N Jacquier. The antimicrobial peptide TAT-RasGAP317-326 inhibits the formation and expansion of bacterial biofilms in vitro. Journal of Global Antimicrobial Resistance 2021; 25, 227-231.
[40] F Shang, L Li, L Yu, J Ni, X Chen and T Xue. Effects of Stigmata maydis on the methicillin resistant Staphylococcus aureus biofilm formation. PeerJ 2019; 7, e6461.
[41] M Setiabudy, DAPS Masyeni, AAG Indraningrat, K Suryawan, IKAI Adhiputra and MABA Rahman. Biofilm formation in Staphylococcus aureus and coagulase-negative Staphylococcus. Folia Medica Indonesiana 2023; 59(3), 222-228.
[42] MMM Raja, N Reehana and AF Begum. Multidrug resistant Staphylococcus aureus isolated from UTI site and its biofilm inhibition activity with nanomaterials in reference with standard antibiotics. International Journal of Zoology Investigations 2022; 8, 152-159.
[43] K Gao, B Su, J Dai, P Li, R Wang and X Yang. Anti-biofilm and anti-hemolysis activities of 10-Hydroxy-2-decenoic acid against Staphylococcus aureus. Molecules 2022; 27(5), 1485.
[44] EM Grimsey and LJ Piddock. Do phenothiazines possess antimicrobial and efflux inhibitory properties? FEMS Microbiology Reviews 2019; 43(6), 577-590.
[45] AKJ Jie, M Hussein, GG Rao, J Li and T Velkov. Drug repurposing approaches towards defeating multidrug-resistant gram-negative pathogens: Novel polymyxin/non-antibiotic combinations. Pathogens 2022; 11(12), 1420.
[46] B Racz and G Spengler. Repurposing antidepressants and phenothiazine antipsychotics as efflux pump inhibitors in cancer and infectious diseases. Antibiotics 2023; 12(1), 137.
[47] JI Perlmutter, LT Forbes, DJ Krysan, K Ebsworth-Mojica, JM Colquhoun, JL Wang, PM Dunman and DP Flaherty. Repurposing the antihistamine terfenadine for antimicrobial activity against Staphylococcus aureus. Journal of Medicinal Chemistry 2014; 57(20), 8540-8562.
[48] C Rauch, L Lucio, BB de Fer and M Lheritier-Barrand. Bioequivalence of 2 pediatric formulations of fexofenadine hydrochloride oral suspension. Clinical Pharmacology in Drug Development 2023; 12(12), 1194-1203.
[49] M Batool, A Zamir, F Alqahtani, T Ahmad, H Saeed and MF Rasool. Clinical pharmacokinetics of fexofenadine: A systematic review. Pharmaceutics 2024; 16(12), 1619.