Trends Sci. 2026; 23(10): 13065
Assessment of the Cytotoxic, Antioxidant, and Antimelanogenic Properties of the Extracts of the Orchid Dendrobium
Pornwipa Phuangbubpha, Patthicha Deesubin, Puretat Saetan,
Sarunyaporn Maksup and Kullanart Obsuwan*
Department of Biology, Faculty of Science, Silpakorn University, Nakhon Pathom 73000, Thailand
(*Corresponding author’s e-mail: Obsuwan_k @su.ac.th)
Received: 17 December 2025, Revised: 31 January 2026, Accepted: 7 February 2026, Published: 10 April 2026
Abstract
Dendrobium extracts are rich sources of bioactive phytochemicals with potential applications in cosmetics and healthcare. This study investigated the phytochemical composition and bioactivities of four hybrid cut-flower Dendrobium cultivars: Sonia ‘Jo Daeng’, Suree Peach, Black Pearl, and White Pearl. The crude flower extracts were evaluated for their antioxidant and antimelanogenic properties, and cytotoxic effects against human fibroblasts, keratinocytes, melanocytes, and macrophages. Among the tested cultivars, Black Pearl flower extracts exhibited the highest contents of phenolics, flavonoids, tannins, sugars, and vitamin C, which correlated with strong DPPH and Ferric Reducing Antioxidant Power (FRAP) antioxidant activities. The extracts were noncytotoxic at concentrations ≤ 800 µg/mL, maintaining > 90% cell viability. The extracts of D. Black Pearl and D. Sonia ‘Jo Daeng’ flowers significantly reduced ROS generation in THP-1-derived macrophages. However, Black Pearl extracts decreased melanin production in α-MSH-stimulated B16F10 melanocytes by ~41.0%, comparable to control. These findings suggest Black Pearl as a promising source of natural antioxidants and antimelanogenic compounds suitable for cosmetic and dermatological applications.
Keywords: Dendrobium hybrids, Antioxidant, Melanogenesis inhibition, Phenolic compounds, Reactive oxygen species reduction, Macrophages, Melanocytes
Introduction
Orchid species have long been recognized as valuable sources of naturally occurring bioactive compounds, and multiple studies have reported the presence of diverse phytochemicals in the genus Dendrobium. Notably, significant amounts of phenolic compounds and flavonoids have been identified in Dendrobium species such as stem and leaf of D. nobile [1], as well as across numerous Dendrobium species, which contain bibenzyl derivatives such as moscatilin and gigantol and flavonoid-related compounds including homoeriodictyol [2]. In addition, polysaccharides and bibenzyl have been widely detected in various Dendrobium spp [3-5]. Collectively, these findings highlight Dendrobium orchids as a rich
botanical resource with strong potential for functional and bioactive applications.
In recent years, growing attention has been directed toward the development of plant-derived ingredients for dermatological and cosmetic products, particularly those targeting oxidative stress, skin inflammation, and hyperpigmentation. Oxidative stress plays a central role in skin aging and dysfunction by promoting cellular damage, impairing barrier integrity, and accelerating inflammatory responses. Reactive oxygen species (ROS) can further exacerbate skin problems by amplifying oxidative injury and contributing to uneven pigmentation and premature aging. Therefore, antioxidant-rich botanical extracts that can reduce ROS accumulation are increasingly investigated as promising candidates for skin-protective and anti-aging cosmeceutical development [6-8].
Regulation of melanogenesis is another key target in cosmetic science, especially for managing hyperpigmentation and achieving uniform skin tone. Melanocytes primarily control melanin biosynthesis and are strongly influenced by environmental stressors, including ultraviolet radiation and oxidative imbalance. Tyrosinase is the rate-limiting enzyme in melanin production, and inhibiting its activity remains a widely used screening strategy for identifying potential skin-brightening agents. Importantly, cell-based melanogenesis models provide biologically relevant confirmation beyond enzyme-based assays, allowing evaluation of both efficacy and cellular responses within melanocytes and thereby strengthening the translational relevance of cosmetic research [9-11]. Consequently, combining antioxidant profiling with tyrosinase inhibition and cell-based melanogenesis assays offers a robust approach for evaluating the cosmeceutical potential of botanical materials.
Several investigations have already shown that Dendrobium species contain phytochemicals relevant to skin health, including compounds associated with antioxidant defense, pigmentation modulation, and cellular protection [12]. Phenolics, flavonoids, and anthocyanins, particularly in flower tissues, have been strongly linked to antioxidant capacity and may contribute to the regulation of oxidative and pigmentary pathways [7,8,13]. Compared with vegetative organs, floral tissues, especially pigmented petals, are metabolically specialized for the accumulation of secondary metabolites involved in UV protection and stress adaptation, making them promising sources of antioxidant and pigmentation-modulating compounds [7,12,14]. In addition, dendropachol isolated from D. pachyglossum has been reported to exhibit no cytotoxic effects on HaCaT keratinocytes at 50 µg/mL after 24 h exposure, supporting the preliminary dermatological safety potential of certain Dendrobium-derived compounds [15].
From a cosmetic science perspective, demonstration of biological efficacy alone is insufficient; botanical extracts must also exhibit cellular safety across multiple skin-relevant cell types. Cytotoxicity screening in fibroblasts, keratinocytes, and melanocytes therefore represents a critical prerequisite for cosmetic applicability, enabling differentiation between functional bioactivity and non-specific cellular stress responses.
Despite these promising pharmacological findings, the practical use of wild Dendrobium species is limited by their slow growth and seasonal flowering, which typically occurs only once per year. In contrast, commercial hybrid Dendrobium orchids cultivated for the cut-flower industry exhibit faster growth and can flower continuously under managed production systems. In Thailand, orchids are among the most important floricultural crops, with large-scale production of Dendrobium flowers concentrated in Kanchanaburi, Ratchaburi, Samut Sakhon, and Nakhon Pathom provinces [16]. Thailand is also a leading exporter of tropical orchids, contributing substantially to the global market and generating significant export revenue. However, oversupply frequently occurs during peak production periods, resulting in severe price declines and causing farmers to discard surplus flowers or sell them at markedly reduced prices. Therefore, the valorization of surplus orchid flowers as functional bio-ingredients may provide both economic and sustainability benefits by increasing product value and supporting more efficient utilization of floral biomass.
Flower pigmentation may further contribute to variability in bioactivity among Dendrobium cultivars. Dark-colored orchid flowers are frequently associated with higher anthocyanin and phenolic contents, which are strongly correlated with enhanced antioxidant potential when compared with light-colored varieties [7,12,15]. However, systematic comparisons linking floral pigmentation, phytochemical composition, and skin-relevant biological activities in commercial hybrid Dendrobium flowers remain limited. Despite accumulating evidence of bioactive compounds in Dendrobium species, integrated studies combining phytochemical profiling with multi-cellular safety assessment and melanogenesis-related bioactivity of commercially available hybrid Dendrobium flower extracts are still scarce.
In this study, crude flower extracts were systematically evaluated for antioxidant activity, tyrosinase inhibitory effects, and anti-melanogenic activity, together with cytotoxicity screening in human dermal fibroblasts, keratinocytes, melanocytes, and macrophages. These endpoints are directly relevant to cosmetic and dermatological applications, as oxidative stress, and hyperpigmentation are key mechanisms underlying skin aging and melanin overproduction. The findings of this work provide scientific support for the development of orchid flower-derived functional ingredients and establish a foundation for value-added utilization of surplus commercial Dendrobium flowers in dermatological and cosmetic formulations.
Materials and methods
Sample collection
All Dendrobium samples were obtained from the Mana Orchid Farm (3.9800° N, 100.0420° ± 0.005° E)., Nakhon Pathom, Thailand during October, 2023.
Preparation of flower extracts
Fresh flowers of D. Sonia ‘Jo Daeng’, D. Black Pearl (dark-colored), and D. Suree Peach, D. White Pearl (light-colored) were harvested at commercial maturity stage. Harvesting was conducted in the morning under standardized commercial conditions. For each cultivar, three independent biological batches were collected and processed separately. The flowers were thoroughly washed 3 - 4 times with tap water and oven-dried at 60 °C for 72 h or until constant weight and subsequently ground into fine powder to ensure microbial safety and consistency with previous Dendrobium studies. While partial degradation of heat-sensitive compounds may occur, identical processing conditions were applied to all samples to maintain comparative validity. For each extraction replicate, 1 g of dried flower powder was extracted with 99.5% methanol at a 1:10 (w/v) ratio (Powder: Solvent) for 24 h at room temperature with continuous agitation [7]. The resulting extracts were concentrated under reduced pressure using rotary evaporation until complete removal of methanol, thereby preventing solvent-related cytotoxic effects in subsequent cell-based assays. All extracts were handled under standardized conditions to ensure reproducibility and suitability for biological evaluation.
Phytochemical quantification
Total phenolic content (TPC)
A 0.1 mL aliquot of the orchid flower extract was mixed with 0.5 mL of 10% Folin-Ciocalteu reagent and dark incubated for 1 min. Subsequently, 1.5 mL of 7.5% Na2CO3 was added, and the volume was adjusted to 4 mL with distilled water. After dark incubation for 30 min, the OD765 was measured using an Optizen 3220UV UV-Vis spectrophotometer (K LAB Co., Ltd., Daejeon, and Republic of Korea). TPC was ascertained using a gallic acid standard curve and expressed as mg gallic acid equivalents (GAE)/g dry weight (DW) [7,17,18].
Total flavonoid content (TFC)
A 0.5 mL aliquot of the extract was mixed with 1.5 mL of methanol, 0.1 mL of 10% AlCl3, and 0.1 mL of 1.0 M potassium acetate. The volume was then adjusted to 5 mL with distilled water. The mixture was incubated for 30 min at room temperature, and the OD415 was determined on the Optizen 3220UV UV–Vis spectrophotometer. TFC was calculated employing a quercetin standard curve and expressed as mg quercetin equivalents (QE)/g DW [17].
Total tannin content (TTC)
TTC was determined by a Folin–Ciocalteu-based colorimetric method, adapted with modifications from Kanlayavattanakul et al. (2018) and Kannaian et al. (2020) [12, 19]. Briefly, a 0.2 mL aliquot of the extract was mixed with 2.5 mL of distilled water and 0.2 mL of Folin-Ciocalteu reagent, followed by 2 mL of 7% Na2CO3. After dark incubation for 90 min, the OD760 was recorded. TTC was estimated using a tannic acid standard curve and expressed as mg tannic acid equivalents (TAE)/g DW.
Total anthocyanin content (TAC) estimated using the pH differential method
TAC was quantified by the pH differential method with adapted from Obsuwan et al. (2019) [7], based on Lee et al. (2005) [20]. A 0.5 mL aliquot of the extract was mixed separately with 4.5 mL of KCl (pH 1.0) or sodium acetate (pH 4.5) buffer. After 15 min of incubation, the OD510 and OD700 were measured. Anthocyanin content was calculated utilizing cyanidin-3-glucoside as a standard and expressed as mg cyanidin-3-glucoside/100 g DW with the formula:
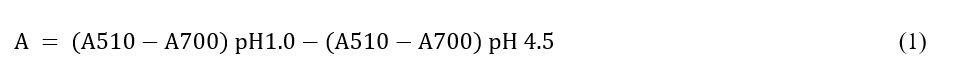
Monomeric anthocyanin content was expressed as mg/L using:
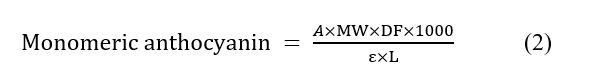
where MW = 449.2 g/mol, DF = 10, ε = 26,900 L·mol⁻¹·cm⁻¹, and L = 1 cm.
Total sugar content (TSC)
The total soluble carbohydrate content (TSC) was determined by the phenol–sulfuric acid method of Dubois et al. (1956), with slight modifications [21]. Briefly, a 0.5 mL aliquot of the extract or glucose standard was mixed with 0.5 mL of 5% phenol and 2.5 mL of concentrated sulfuric acid. The mixture was vortexed and incubated at room temperature for 30 min, after which absorbance was measured at 490 nm using an Optizen 3220UV UV–Vis spectrophotometer. Analyses were performed in triplicate, and results were expressed as mg glucose equivalents (GE)/g DW.
Reducing sugar content (RSC)
Reducing sugars were determined by a dinitrosalicylic acid (DNS)-based assay adapted from Miller (1959) and Leitão et al. (2023) with modifications [22,23]. A 0.5 mL aliquot of the extract was added to 0.5 mL of the DNS reagent, heated at 90 °C for 5 min, and cooled on an ice bath. After cooling, 2.5 mL of distilled water was added. The OD540 was ascertained with an Optizen 3220UV UV–Vis spectrophotometer. The assay was performed in triplicate, and results were expressed as mg glucose equivalents/g DW.
Vitamin C content
Vitamin C was determined by a ferric ion reduction–1,10-phenanthroline colorimetric method, adapted from Li et al. (2012) with modifications. [24]. For this, 1 g of dried orchid flowers was extracted with 10 mL of 5% TCA, vortexed, and centrifuged at 10,000 rpm for 10 min. Then, 0.5 mL of the supernatant was mixed with ethanol, 0.4% phosphoric acid–ethanol, 0.5% 1,10-phenanthroline–ethanol, and 0.03 g/L ferric chloride, and incubated at room temperature for 30 min. The OD534 was recorded with an Optizen 3220UV UV–Vis spectrophotometer. Vitamin C content was calculated based on an ascorbic acid standard curve and expressed as mg/g DW.
Antioxidant activity assay
2,2-diphenyl-1-picrylhydrazyl (DPPH)-based free radical scavenging assay
Free radical scavenging activity was determined by the DPPH assay, adapted from Obsuwan et al. (2019) with modifications [7]. Briefly, the extract, at different concentrations (2.5–40 mg/mL), was mixed with 2 mL of 0.2 mM DPPH in methanol and dark incubated for 30 min. The OD515 was recorded. Free radical scavenging activity was ascertained by applying the formula:
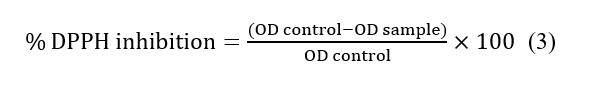
The IC50 values were determined, and the antioxidant activity was expressed as mg Trolox equivalents/g extract.
Ferric reducing antioxidant power (FRAP) Assay
FRAP activity was determined with slight modifications using 150 µL of the extract mixed with 2.85 mL of the FRAP reagent (acetate buffer, FeCl₃, and TPTZ). After incubation at 37 °C for 30 min, the OD593 was measured. The results were compared with a FeSO₄ standard curve and expressed as mM Fe²⁺/g DW [17].
Tyrosinase inhibition assay
Tyrosinase inhibitory activity was evaluated using mushroom tyrosinase (Agaricus bisporus), a commonly used screening enzyme for tyrosinase inhibition studies [25]. The assay was adapted from Maksup and Obsuwan (2020), with modifications [8]. Briefly, a0.1 mL aliquot of the extract or standard (kojic acid at 5 mg·L⁻¹, L‑ascorbic acid at 5 mg·L⁻¹, or quercetin at 3 mg·L⁻¹, corresponding to the concentrations used for the reference inhibitors in Figure 1(H) was mixed with 1.8 mL of sodium phosphate buffer (pH 6.8) and 0.1 mL of tyrosinase (138 U). The mixture was incubated for 10 min at room temperature and then added with 1 mL of 2.5 mM Levodopa (L-DOPA). After further incubation for 10 min, the OD475 was recorded with an Optizen 3220UV UV-Vis spectrophotometer. Inhibition was calculated employing the formula:
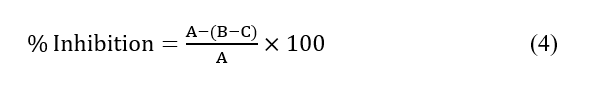
where A = absorbance of the control, B = absorbance of the sample with tyrosinase, and C = absorbance of the sample without tyrosinase.
Cell culture
The human monocytic leukemia cell line THP-1 (JCRB0112.1, lot#: 11102017) was obtained from the Japanese Collection of Research Bioresources Cell Bank, Osaka, Japan. The human keratinocyte cell line HaCaT and MRC-5 human fibroblasts were purchased from the American Type Culture Collection (ATCC), VA, USA. The murine melanoma cell line B16F10 was generously provided by Chulalongkorn University, Thailand. Cells were maintained in tissue culture flasks (T-25; Nunclon, Thermo Fisher Scientific, Shanghai, China) with complete growth medium (CGM). For the adherent cells, B16F10, MRC-5, and HaCaT, the CGM consisted of Dulbecco’s Modified Eagle’s Medium (DMEM; HyClone, Cytiva Life Sciences, UT, USA) supplemented with low or high levels of glucose, 10% fetal bovine serum (FBS; HyClone), 2 mM L-glutamine (Corning Inc., NY, USA), and 1% penicillin + streptomycin (HyClone). For the suspension cells, THP-1, Roswell Park Memorial Institute medium (RPMI-1640; Gibco, Thermo Fisher Scientific, MA, USA; cat#: 21870076, lot#: 2051389), added with 10% FBS, 2 mM L-glutamine, and 1% penicillin + streptomycin, was the CGM.
Cell viability assays and crystal violet staining
Cytotoxicity was assessed using the resazurin cell viability assay. HaCaT, MRC-5, or B16F10 cells were seeded in 96-well plates at 1.5×10⁴ cells/well. To induce differentiation into macrophage-like cells (THP-1m), THP-1 cells were seeded in 96-well plates at 3.5×10⁴ cells/well and treated with 100 ng/mL phorbol 12-myristate 13-acetate (PMA; Sigma-Aldrich, MO, USA; cat#: 16561298) for 48 h. The differentiated THP-1m cells were treated with 50 - 800 µg/mL of orchid extracts for 72 and 120 h (HaCaT, B16F10, and MRC-5) and 72 h for THP-1m. Next, 120 µL of 4.4 µM resazurin was added and dark incubated for 4 h. OD570 and OD600 were measured using a microplate reader.
Cytotoxicity of the crude extracts was assessed using crystal violet staining followed by morphological observation. After treatment for the required duration, the cells were washed twice with PBS, fixed in 70% ethanol for 10 min, and stained with 0.5 %w/v crystal violet for 30 min. Excess dye was removed by rinsing with PBS. After air-drying, cell morphology was examined under a CK30 inverted microscope (Olympus Tokyo, Japan).
Reactive oxygen species (ROS) quantification
Intracellular ROS levels were determined using 2′,7′-dichlorodihydrofluorescein diacetate (DCFH-DA) staining, as previously described for THP-1-derived macrophages with modifications [26]. THP-1-derived macrophages (THP-1m) were seeded in 96-well plates at a density of 3.5 × 10⁴ cells/well and pretreated with each orchid extract at 400 µg/mL for 72 h. This concentration was selected based on preliminary cytotoxicity screening in the present study, which showed >90% cell viability and therefore allowed assessment of antioxidant effects without cytotoxic interference. Inflammation was then induced with lipopolysaccharide (LPS, 100 ng/mL) for 24 h. Cells treated with tert-butyl hydroperoxide (TBHP, 50 µM for 30 min) served as a positive control for ROS generation. After treatment, cells were washed with PBS and incubated with 20 µM DCFH-DA for 45 min in the dark. Fluorescence images were captured using a fluorescence microscope, and intracellular ROS levels were quantified as corrected total cell fluorescence from at least three images per condition using ImageJ software and expressed relative to the untreated control.
Melanin contents
To evaluate the melanogenesis suppression capability of orchid extracts, the B16F10 cells were seeded in 6-well plates for 48 h at 1×106 cells/well. The medium was then replaced with a fresh one containing 100 nM melanocyte-stimulating hormone (α-MSH) and incubated for 24 h. The cells were then treated with orchid extracts at 400 µg/mL for 72 h. Then, the cells were disassociated with 0.05 µM trypsin and lysed employing 10% dimethyl sulfoxide (DMSO) in 1N NaOH, at 70 °C for 1 h. Next, 100 µL of each supernatant was transferred into a 96-well plate, and the OD405 was measured with a microplate reader.
Statistical analysis
All experiments were performed using three independent biological replicates (n = 3) and presented as the means ± standard deviation (SD). Statistical significance was evaluated using Duncan’s multiple range test (DMRT) at p < 0.05 with SPSS Statistics version 26 (IBM, NY, USA).
Results and discussion
Although the phytochemical composition and biological activities of Dendrobium species have been reported previously, most studies focus on individual compounds or single biological endpoints. In contrast, the present study adopts an integrated, application-oriented approach by valorizing surplus hybrid Dendrobium flowers as potential cosmetic bio-resources. By systematically comparing dark- and light-colored commercial cultivars, and by combining phytochemical profiling with multi-cellular safety assessment and melanogenesis-related bioactivity, this work provides new insight into cultivar-specific functional differences. Importantly, the study frames orchid flower by-products not merely as sources of antioxidants, but as differentiated bio-ingredients with distinct biological profiles, thereby supporting sustainable utilization within a circular economy framework.
Phytochemicals
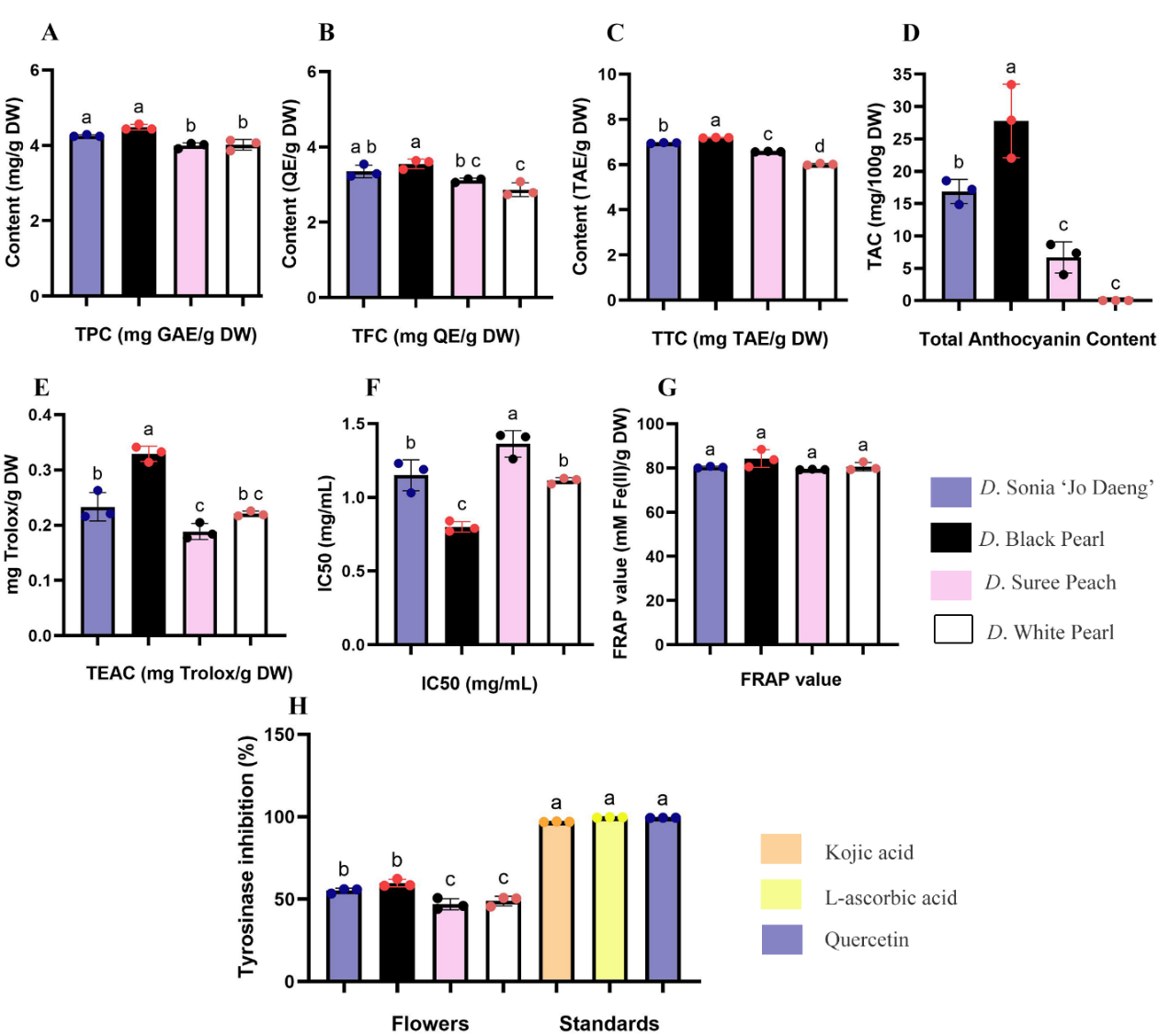
All Dendrobium hybrids contained phytochemicals at varying levels (Figures 1(A) - 1(C)). The TPC ranged from 4.00 ± 0.07 to 4.48 ± 0.07 mg GAE/g DW, TFC from 2.87 ± 0.18 to 3.55 ± 0.13 mg QE/g DW, and TTC from 6.01 ± 0.04 to 7.19 ± 0.02 mg TAE/g DW. Black Pearl exhibited the highest TPC (4.48 ± 0.07 mg GAE/g DW), TFC (3.55 ± 0.13 mg QE/g DW), and TTC (7.19 ± 0.02 mg TAE/g DW). TFC did not markedly differ between Black Pearl and ‘Sonia Jo Daeng’. The lowest TPC was observed in White Pearl, Suree Peach showed the lowest TFC and TTC. The higher phenolic and anthocyanin contents observed in the darker cultivar extracts may partially explain their stronger antioxidant activity, consistent with the known redox-active properties of these phytochemical groups.
These findings are consistent with previous reports showing that purple-flowered orchid hybrids may contain elevated levels of flavonoids and anthocyanin-related pigments in floral tissues [27]. These compounds exhibit free radical-scavenging activity and may contribute to cellular protection against oxidative damage, which is associated with chronic diseases such as cardiovascular disease, cancer, and diabetes [28,29]. In addition, tannins are polyphenolic constituents known to contribute to antioxidant, anti-inflammatory, and antimicrobial activities and are therefore of interest for pharmaceutical and cosmetic applications [6]. In Dendrobium flower extracts, tannin content has been quantified together with antioxidant and enzyme inhibitory activities, supporting their potential as specialty cosmetic ingredients [12].
The TAC was highest in Black Pearl (27.78 ± 5.67 mg cyanidin-3-glucoside/100 g DW), while anthocyanins were undetectable in White Pearl (Figure 1(D)). This pattern is consistent with findings in Rhynchostylis gigantea, in which anthocyanins were likewise not detected in the white-flowered form [30]. In Dendrobium flower extracts, pelargonidin and cyanidin have been identified among the major anthocyanins, and anthocyanins are widely recognized as pigments contributing to red-to-purple floral coloration and antioxidant activity [12,28]. Therefore, the greater accumulation of anthocyanins in the darker hybrid, particularly Black Pearl, may partly explain its stronger antioxidant performance relative to the paler hybrids. Beyond their general antioxidant capacity, anthocyanins such as cyanidin-3-O-glucoside may also contribute to antimelanogenic activity through interaction with the catalytic copper center of tyrosinase, although their inhibitory potency appears to depend on structural features and assay context [31]. In parallel, polyphenol- and flavonoid-rich extracts have been shown to suppress melanogenesis by downregulating MITF and downstream melanogenesis-related genes, including TYR, TRP-1, and TRP-2 [32]. Collectively, these mechanisms provide a plausible explanation for the strong antimelanogenic effect observed in the Black Pearl cultivar, which contained the highest anthocyanin content among the tested flowers.
The TSC and RSC ranged from 82.15 ± 4.00 to 100.00 ± 1.19 and from 93.27 ± 2.07 to 143.17 ± 0.83 mg/g DW, respectively. Black Pearl exhibited the maximum levels of both, while Suree Peach had the least. Vitamin C contents ranged from 1.35 ± 0.22 to 3.37 ± 0.04 mg/g DW in Black Pearl, with Sonia ‘Jo Daeng’ containing markedly higher amounts. The elevated sugar levels observed in the darker flowers may be linked to enhanced anthocyanin biosynthesis, as sucrose functions as a signaling molecule that induces anthocyanin accumulation and upregulates anthocyanin biosynthetic genes [33,34]. Vitamin C may also contribute to the antioxidant status of floral tissues, as ascorbate functions as a redox-active co-antioxidant involved in free radical scavenging and maintenance of cellular redox balance [35]. Collectively, these results indicate that enhanced sugar and vitamin C levels of Black Pearl may support enhanced pigmentation and antioxidant capacity. Nevertheless, these results should be interpreted with caution. Drying at elevated temperatures, as applied in the present study, may induce partial degradation of thermolabile compounds such as vitamin C and anthocyanins, potentially leading to an underestimation of their actual concentrations [36,37]. Furthermore, the colorimetric method used for vitamin C determination may overestimate true vitamin C levels due to interference from other reducing substances present in crude extracts. While suitable for comparative analysis across samples, this limitation highlights the need for more specific analytical approaches. Future studies employing high-performance liquid chromatography (HPLC) would enable compound-specific quantification and provide a more accurate assessment of vitamin C stability in relation to floral pigmentation and antioxidant activity [38]. This study is limited to flower-derived crude extracts and in vitro screening models. Future studies should further identify active compounds using chromatographic profiling (HPLC/LC–MS), investigate melanogenesis-related signaling pathways, and validate efficacy and safety in advanced skin models or clinical settings. In addition, formulation studies including stability and compatibility testing are necessary to support potential cosmetic application.
Biological activities
The antioxidant capacity of the extracts, assessed employing DPPH and FRAP assays, was maximal in Black Pearl (DPPH: 0.40 ± 0.17 mg Trolox/g DW and IC₅₀: 0.80 ± 0.36 mg/mL; FRAP: 84.26 ± 3.99 mM Fe(II)/g DW) (Figures 1(E) - 1(G)). These findings are consistent with the elevated TPC, TFC, TTC, and anthocyanin contents observed in this cultivar. Such a relationship is plausible because the hydroxyl (–OH) groups of phenolic compounds can donate hydrogen atoms to free radicals, thereby interrupting oxidative chain propagation and enhancing radical-scavenging activity [39]. Such antioxidant-rich extracts may contribute to suppressing oxidative stress-related disorders, including atherosclerosis, cancer, and cardiovascular diseases [40,41]. ROS suppression in activated macrophage-like cells suggests that the extracts may exert antioxidant effects under inflammatory oxidative conditions [42,43], supporting potential cosmetic relevance in protecting skin from oxidative and inflammation-related stress [11,15].
All extracts demonstrated notable tyrosinase inhibitory activity, ranging from 46.90 ± 3.37% to 59.71 ± 2.31% (Figure 1(H)). Black Pearl exhibited the strongest inhibition (59.71 ± 2.31%), comparable to Sonia ‘Jo Daeng’ (55.19 ± 1.50%), whereas Suree Peach and White Pearl showed the weakest suppressive effects. The positive association between tyrosinase inhibition and total flavonoid content suggests that flavonoids may contribute, at least in part, to melanogenesis suppression through direct tyrosinase inhibition. Proposed mechanisms include interaction with the copper-containing active site of the enzyme and competitive effects on substrate-related catalytic processes, depending on flavonoid structure [44,45]. While mushroom tyrosinase inhibition provides an initial screening of anti-melanogenic potential [9], melanocyte-based models offer greater biological relevance for evaluating pigmentation-related responses in the skin, particularly in the context of oxidative stress and photoaging [10,11]. In this study, the observed suppression of melanin accumulation in α-MSH-stimulated melanocytes therefore provides biologically relevant support for the anti-melanogenic potential of the extracts.
Table 1 Total sugar content, reducing sugar, and vitamin C contents of the flower extracts of four Dendrobium hybrids.
|
Total sugar (mg/g/DW) |
Reducing sugar (mg/g/DW) |
Vitamin C (mg/g/DW) |
D. Sonia ‘Jo Daeng’ |
96.52 ± 0.89a |
132.02 ± 0.83b |
3.33 ± 0.167a |
D. Black Pearl |
100.07 ± 0.97a |
143.17 ± 0.68a |
3.38 ± 0.03a |
D. Suree Peach |
82.15 ± 3.3b |
115.20 ± 0.61c |
1.54 ± 0.21b |
D. White Pearl |
86.88 ± 0.65b |
93.27 ± 1.68d |
Values are presented as the means ± standard deviation (SD) of three independent replicates. The various letters in superscripts within the same column indicate statistically significant differences among groups (DMRT, p < 0.05; n = 3).
![]()
Figure 1 Phytochemical contents and biological activities of the flower extracts of Dendrobium hybrids.
(A) Total phenolic content (TPC), (B) total flavonoid content (TFC), (C) total tannin content (TTC), and (D) total anthocyanin content (TAC). Antioxidant capacities were represented as (E) Trolox equivalent antioxidant capacity (TEAC), (F) DPPH radical scavenging activity expressed as IC₅₀ values, and (G) Ferric reducing antioxidant power (FRAP) assay. (H) Tyrosinase inhibitory activity of Dendrobium flower extracts compared with those of the standard compounds, kojic acid, L-ascorbic acid, and quercetin. Values represent mean ± SD (n = 3). Different letters indicate statistically significant differences (DMRT, p < 0.05). Abbreviations: TPC, total phenolic content; TFC, total flavonoid content; TTC, total tannin content; TAC, total anthocyanin content; FRAP, ferric reducing antioxidant power.
Morphological observations of the human fibroblasts, keratinocytes, and melanocytes, indicating cytotoxicity
Safety evaluation using different skin cell types is a critical step in developing plant-derived ingredients for cosmetic applications. The skin consists of several major cell populations, including fibroblasts, keratinocytes, and melanocytes. Assessing the effects of plant extracts on these representative cell types provides a realistic indication of their suitability for topical use. The crude extracts of Sonia ‘Jo Daeng’, Suree Peach, Black Pearl, and White Pearl flowers were evaluated for their cytotoxicity against MRC-5 human fibroblasts, HaCaT keratinocytes, and B16F10 melanocytes. After 3 and 5 days of exposure, flower extracts at concentrations ranging from 50 to 800 µg/mL were non-cytotoxic, with relative cell viability > 90% (Figures 2(A) - 2(C), Table 2). Crystal violet staining further confirmed these results; the OD570 did not vary significantly compared with the control, indicating that none of the extracts reduced cell viability (Figures 2(D) - 2(F)). Microscopic examinations supported these findings, revealing that cell morphology and confluency were preserved across all treatments. Images taken on day 5 showed MRC-5 fibroblasts with normal elongated shapes comparable to the control group, with no evidence of shrinkage or detachment (Figure 2(G)). These results suggest that Dendrobium flower extracts were biocompatible and safe for subsequent bioassays. This safety profile may be attributed, at least in part, to their phytochemical composition. Alkaloids are nitrogen-containing natural products that are widely distributed in plants and are known to function in ecological defense against herbivores and pathogens [46,47]. However, many alkaloids also exhibit strong bioactivity, including cytotoxic effects against cancer cells [47]. In D. officinale, transcriptomic analysis together with organ-specific alkaloid quantification showed that total alkaloids accumulate most strongly in leaves, whereas flowers contain lower levels, supporting organ-specific differences in alkaloid accumulation among tissues [48].
Table 2 Cell viability of MRC-5 human fibroblasts, HaCaT keratinocytes, and B16F10 melanocytes after 3 days of exposure to the flower extracts of four Dendrobium varieties.
|
%Cell viability (800 µg/mL of flower extracts) |
||
|
MRC-5 |
HaCaT |
B16F10 |
D. Sonia ‘Jo Daeng’ |
106.0 ± 2.88a |
97.6 ± 4.11a |
102.0 ± 2.57a |
D. Black Pearl |
102.0 ± 9.78a |
106.0 ± 7.09a |
98.4 ± 3.16a |
D. Suree Peach |
103.0 ± 5.01a |
107.0 ± 4.62a |
101.0 ± 2.35a |
D. White Pearl |
100.4 ± 5.59a |
102.0 ± 5.13a |
98.7 ± 5.18a |
Values are presented as the means ± standard deviation (SD) of three independent replicates. The various letters in superscripts within the same column indicate statistically significant differences among groups (DMRT, p < 0.05; n = 3).
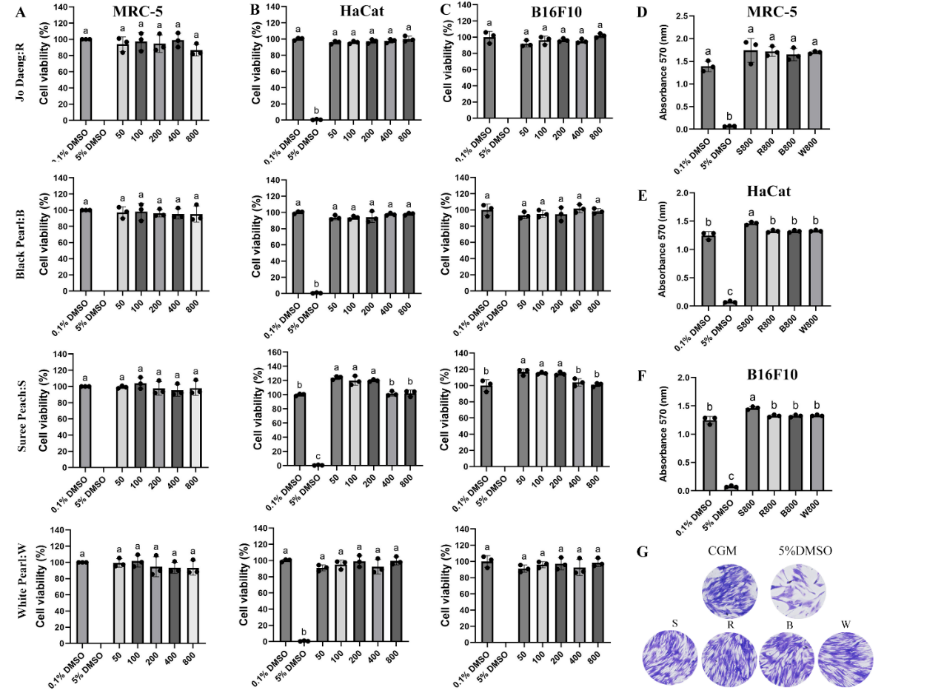
Figure 2 Cytotoxicity of Dendrobium flower extracts assessed using human fibroblasts (MRC-5), keratinocytes (HaCaT), and melanocytes (B16F10) based on cell morphology.
(A) - (C) Percent cell viability of MRC-5, HaCaT, and B16F10 cells after 5 days of treatment with various concentrations (50 - 800 µg/mL) of flower extracts of the Dendrobium cultivars Sonia ‘Jo Daeng’, Black Pearl, Suree Peach, and White Pearl. (D) - (F) Representative OD570 post-crystal violet staining, showing viable cell density on day 5, after treatment with orchid flower extracts. (G) Morphologically, the MRC-5 cells showed a typical shape on day 5 of exposure. Values represent mean ± SD (n = 3). Different letters indicate statistically significant differences (DMRT, p < 0.05).
Antioxidant/Redox-modulating effects
Before evaluating antioxidant/redox-modulating effects, the cytotoxicity of the four extracts was examined using THP-1-derived macrophages. After 3 days of exposure to the extracts at concentrations between 50 and 800 µg/mL, the macrophage viability remained > 90% for all treatments, confirming their safety toward immune cells (Figure 3(A)). The oxidative stress-inducing potential of the extracts was assessed by measuring their ROS-generating ability using DCFH-DA staining. None of the four extracts induced ROS formation in unstimulated macrophages after 3 days of treatment (Figure 3(B)), confirming their non-pro-oxidative and biocompatible nature. In contrast, the ROS generated by LPS pre-treatment was effectively suppressed by the extracts of Sonia ‘Jo Daeng’ and Black Pearl flowers at 400 µg/mL. Notably, with Black Pearl extracts, the ROS levels decreased the most, by 3.43-fold compared with the LPS control (Figure 3(C)). We acknowledge that intracellular ROS is an upstream mediator rather than a direct marker of the inflammatory response; therefore, in this study we interpret the LPS‑induced ROS reduction primarily as evidence of antioxidant and redox‑modulating activity in activated macrophages, which may secondarily contribute to reduced intracellular ROS levels profile rather than constituting definitive proof of anti‑inflammatory action. Accordingly, we have softened our wording and now refer to “ROS reduction in LPS-stimulated THP-1 macrophages” instead of claiming direct Antioxidant and redox-modulating effects, and we recognize the lack of canonical inflammatory readouts (e.g., TNF‑α, IL‑6, IL‑1β, COX‑2, or NF‑κB activation) as a limitation of the present work. Such a variation in the antioxidant influence of Dendrobium cultivars appears to be closely related to their pigment-associated phytochemical composition. Dark-colored orchids such as Black Pearl and Sonia ‘Jo Daeng’ exhibited more robust ROS scavenging than the light-colored varieties (Suree Peach and White Pearl), which can be attributed to their higher contents of anthocyanins, phenolics, and flavonoids, compounds with electron-donating and redox-modulating properties [14]. Anthocyanins act as chain-breaking antioxidants that directly neutralize reactive oxygen intermediates, while phenolic acids and flavonoids can chelate metal ions, thereby stabilizing the ROS contents through hydrogen atom and single electron transfer mechanisms [41]. By attenuating oxidative stress in LPS‑stimulated macrophages, these phytochemicals are likely to modulate redox‑sensitive signaling pathways such as NADPH oxidase and NF‑κB, which are critically involved in the production of pro‑inflammatory mediators [10,32], although specific cytokine and enzyme markers were not measured in this study and should be addressed in future work. Conversely, the cultivars with light-colored flowers, which contained fewer pigmented polyphenols, showed limited ROS-scavenging efficiency, consistent with their lower TPC and TAC.
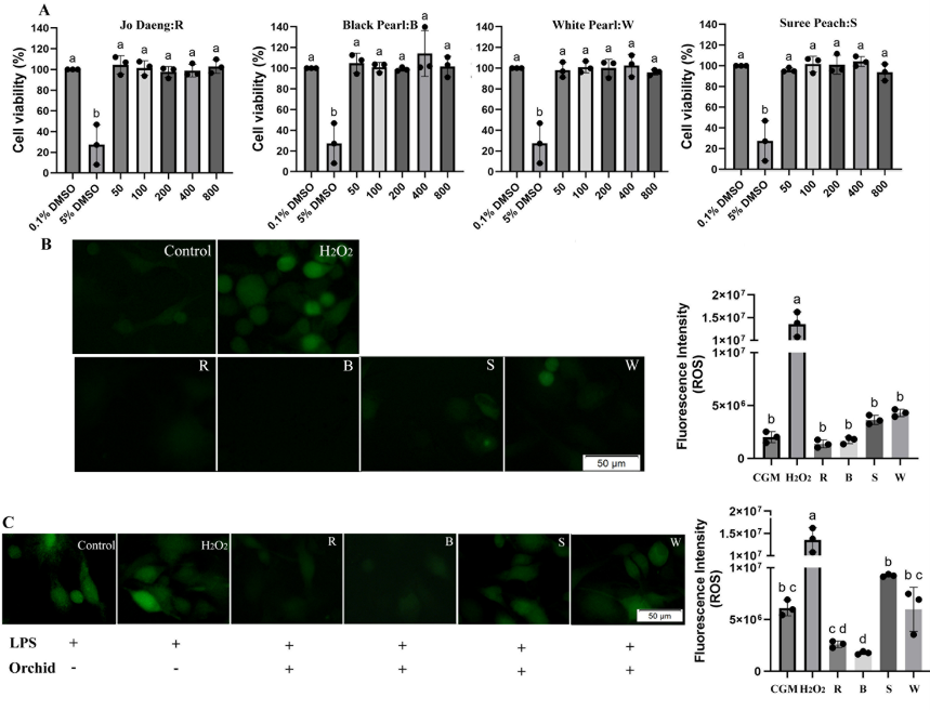
Figure 3 Antioxidant effects of Dendrobium flower extracts in THP-1-derived macrophages. (A) Cell viability (%) after 3 days of exposure to Sonia ‘Jo Daeng’ (R), Black Pearl (B), Suree Peach (S), and White Pearl (W) extracts at 50 - 800 µg/mL, determined via the resazurin assay. (B) Intracellular ROS generation in unstimulated macrophages after 3 days of treatment with orchid extracts (400 µg/mL), visualized using DCFH-DA fluorescence microscopy. (C) ROS levels in LPS-stimulated macrophages (100 ng/mL, 24 h) treated with orchid extracts at 400 µg/mL. Values represent mean ± SD (n = 3). Different letters indicate statistically significant differences (DMRT, p < 0.05).
Anti-melanogenic activity
The ability of Dendrobium flower extracts to inhibit melanin biosynthesis was investigated via melanin content assays. B16F10 melanocytes were treated with the crude flower extracts at 400 µg/mL, while for the positive controls, 100 µg/mL of kojic acid was added. These concentrations were selected based on the non-cytotoxic levels determined employing the cytotoxicity assays. Melanin synthesis was stimulated with 100 nM α-MSH, and melanin content was measured after 3 days. Among the tested cultivars, Black Pearl flower extracts significantly reduced melanin accumulation compared with the +α-MSH control, by ~41.17 ± 0.12%, indicating a 2.55- and 2.81-fold reduction relative to Suree Peach and White Pearl flower extracts, respectively. The inhibitory ability of Black Pearl flower extracts was comparable to that of kojic acid, whereas that of Sonia ‘Jo Daeng’ showed moderate suppression. Microscopic observations corroborated these results; cells treated with Black Pearl flower extracts and kojic acid exhibited lighter pigmentation and fewer intracellular melanin granules, while the +α-MSH, Suree Peach, and White Pearl flower extracts-treated groups remained strongly pigmented.
Notably, the White Pearl extract increased melanin content to approximately 180% of the baseline level, which was even higher than that of the +α MSH control (~170%), indicating a lack of anti-melanogenic activity and a potential pro melanogenic or α MSH–synergistic effect under the present experimental conditions.
Melanin, synthesized within the melanosomes of melanocytes, plays a critical photoprotective role by helping shield the skin from ultraviolet-induced damage and oxidative stress [10,11]. However, chronic UV exposure can dysregulate melanogenesis and contribute to hyperpigmentation [10,11,49]. Thus, identifying safe natural compounds that can modulate tyrosinase activity or suppress the expression of melanogenesis-related genes remains a major focus in skin-lightening research [50]. In the present study, Black Pearl flower extracts exhibited the maximal antimelanogenic effect, consistent with their highest phenolic, flavonoid, tannin, anthocyanin, and vitamin C contents. This effect may be related, at least in part, to the antioxidant-rich composition of the extract, because natural antioxidants can support melanocyte redox homeostasis [11]. In consistency, Sebok (SB) rice seed extracts, enriched in polyphenols (~21.6 mg TAE/g extract) and flavonoids (~14.1 mg QE/g extract), markedly suppressed melanogenesis in melan-a cells without affecting viability. At 100 µg/mL, SB reduced cellular tyrosinase activity by ~53.8% and downregulated MITF, TYR, TRP-1, and TRP-2 expression [32]. Additional studies have likewise shown that natural compounds can suppress melanogenesis in B16F10 cells through regulation of melanogenesis-related signaling pathways [50,52]. It should be noted that cell-free mushroom tyrosinase inhibition and cell-based melanogenesis assays evaluate different levels of biological response; cell-based systems additionally reflect intracellular regulation of tyrosinase activity, melanin accumulation, and melanogenic signaling [32,51,52]. This discrepancy may explain why some extracts that showed comparable mushroom tyrosinase inhibition, such as Suree Peach and White Pearl, did not reduce intracellular melanin accumulation in B16F10 cells; in the case of White Pearl, melanin content was even increased. These findings indicate that inhibition of mushroom tyrosinase in a cell-free assay does not necessarily translate into suppression of cellular melanogenesis, which is additionally influenced by intracellular uptake, metabolic transformation, substrate availability, and melanogenic signaling pathways, including the α-MSH/MITF axis and downstream melanogenesis-related genes [9,32,52]. It is therefore plausible that certain phytoconstituents present in these lighter-colored cultivars may stimulate melanogenic signaling or counteract direct enzyme inhibition under cellular conditions, thereby overriding the inhibitory effect observed in the cell-free tyrosinase assay.
Here, the cultivars with dark-colored flowers (Black Pearl and Sonia ‘Jo Daeng’), which have higher anthocyanin and phenolic levels, exhibited a more robust depigmenting influence than those with lighter-colored flowers (Suree Peach and White Pearl). This divergence between dark- and light-colored cultivars underscores the importance of cell-based assays for distinguishing truly depigmenting extracts from those with neutral or even pro melanogenic effects, despite similar performance in enzyme-based screening assays [9]. Anthocyanins most likely contribute through dual mechanisms: Direct tyrosinase inhibition and oxidative signaling modulation that promotes melanogenesis [31,41]. The strong association between antioxidant capacity and melanin synthesis suppression suggests that the whitening activity of Dendrobium extracts primarily operates through redox rather than cytotoxicity regulation. Collectively, these findings highlight dark-pigmented Dendrobium flowers, particularly of Black Pearl, as promising sources of safe and effective natural whitening agents. In contrast, Suree Peach and White Pearl are not considered suitable whitening candidates based on the present data and may warrant further investigation to identify specific constituents responsible for their potential pro melanogenic activity. Their combined antioxidant, and tyrosinase-inhibitory activities suggest the potential use of surplus cut-flower orchids in cosmeceutical formulations aimed at alleviating hyperpigmentation and photoaging.
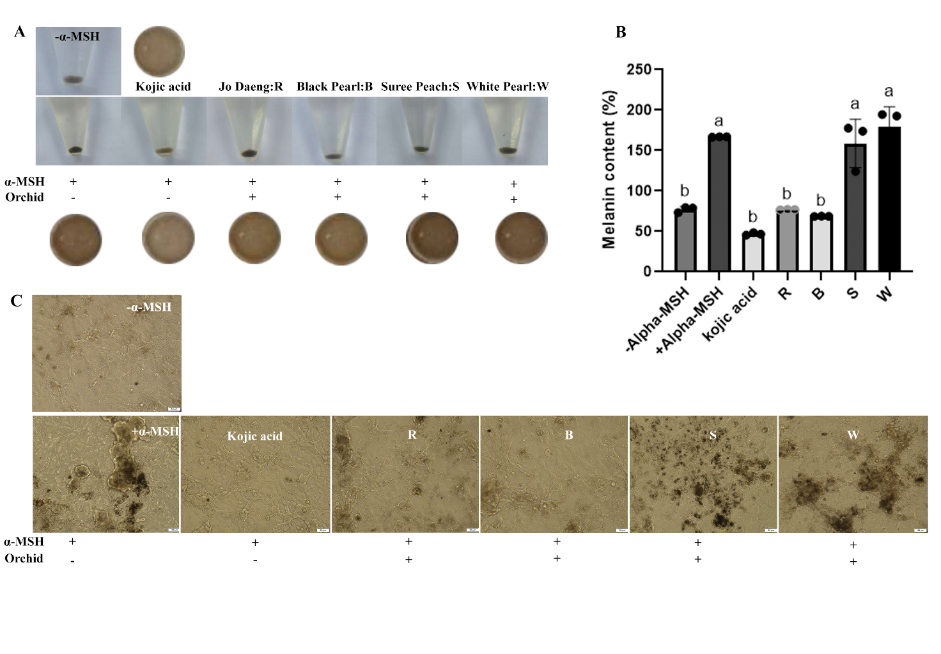
Figure 4 Anti-melanogenic effects of Dendrobium flower extracts determined using B16F10 melanocytes. (A) The appearance of melanin pellets in α-MSH-stimulated cells treated with Sonia ‘Jo Daeng’ (R), Black Pearl (B), Suree Peach (S), and White Pearl (W) flower extracts (400 µg/mL), compared with those exposed to α-MSH (control) and kojic acid (100 µg/mL). (B) Melanin content (%) relative to the −α-MSH control. (C) Representative micrographs showing intracellular melanin accumulation in α-MSH-stimulated B16F10 cells post-treatment with each extract. Data are expressed as the means ± SD (n = 3). Values represent mean ± SD (n = 3). Different letters indicate statistically significant differences (DMRT, p < 0.05).
Conclusions
This study highlights the remarkable potential of Dendrobium flowers, particularly of the hybrid Black Pearl, as natural sources of antioxidant and anti-melanogenic agents. Black Pearl flower extracts exhibited the highest phenolic, flavonoid, tannin, sugar, and vitamin C contents, resulting in a strong antioxidant ability. Its extracts were compatible with human fibroblasts, keratinocytes, melanocytes, and macrophages, maintaining > 90% cell viability at ≤ 800 µg/mL. Moreover, Black Pearl flower extracts effectively reduced ROS generation and melanin biosynthesis at levels comparable to those of kojic acid. Collectively, these findings establish a robust scientific foundation for the development of Dendrobium-derived cosmetic formulations targeting oxidative stress and hyperpigmentation. Beyond cosmetic applications, the demonstrated safety and potent antioxidant properties of Black Pearl flower extracts also suggest their potential for further exploration as functional bioingredients for food or nutraceutical applications. Utilizing surplus flowers of hybrid orchids as high value bioingredients could therefore enhance the economic and sustainable use of Thailand’s floricultural resources while advancing innovation in the cosmetic, nutraceutical, and healthcare industries.
This study is limited to flower-derived crude extracts and in vitro screening models. Future studies should further include cytokine profiling and inflammation-related signaling pathways to substantiate the anti-inflammatory potential suggested by ROS modulation, identify active compounds using chromatographic profiling (HPLC/LC–MS), investigate melanogenesis-related signaling pathways, and validate efficacy and safety in advanced skin models or clinical settings. In addition, formulation studies including stability and compatibility testing are necessary to support potential cosmetic application.
Acknowledgments
This work was supported by the Faculty of Science, Silpakorn University, Bangkok, Thailand, for research funding with serial number SRIF-PRG-2567-07.
Declaration of Generative AI in Scientific Writing
The authors acknowledge the use of generative AI tools (e.g., QuillBot and ChatGPT by OpenAI) in the preparation of this manuscript, specifically for language editing and grammar correction. No content generation or data interpretation was performed by AI. The authors take full responsibility for the content and conclusions of this work.
CRediT Author Statement
Pornwipa Phuangbubpha: Methodology, Investigation, Data analysis, Writing-original draft preparation. Patthicha Deesubin: Methodology, Data analysis, Writing-original draft preparation. Puretat Saetan: Methodology, Investigation. Sarunyaporn Maksup: Methodology, Investigation. Kullanart Obsuwan: Supervision, Conceptualization, Methodology, Investigation, Resource, Writing- Reviewing and Editing.
References
[1] P Li, T shen, L Li and Y Wang. Optimization of the selection of suitable harvesting periods for medicinal plants: Taking Dendrobium officinale as an example. Plant Methods 2024; 20(1), 43.
[2] R Choonong, W Sermpradita, T Kitisripanyac, B Sritularakd and W Putalun. The contents of bibenzyl derivatives, flavonoids and a phenanthrene in selected Dendrobium spp. and the correlation with their antioxidant activity. ScienceAsia 2019; 45(3), 245-252.
[3] M Chinsamy, J Finnie and JV Staden. Anti-inflammatory, antioxidant, anti-cholinesterase activity and mutagenicity of South African medicinal orchids. South African Journal of Botany 2014; 91, 88-98.
[4] GY Liu, L Tan, L Cheng, LS Ding, Y Zhou, Y Deng, YQ He, DL Guo and SJ Xiao. Dendrobine-type alkaloids and bibenzyl derivatives from Dendrobium findlayanum. Fitoterapia 2020; 142, 104497.
[5] S Zheng, Y Zhu, C Jiao, M Shi, L Wei, Y Zhou, Q Jin and Y Cai. Extraction and analysis of gigantol from Dendrobium officinale with response surface methodology. Molecules 2018; 23(4), 818.
[6] J Olivero-Verbel, P Quintero-Rincón and K Caballero. Aromatic plants as cosmeceuticals: benefits and applications for skin health. Planta 2024; 260, 132.
[7] K Obsuwan, JB Ryong and S Maksup. Analysis of bioactive compounds, polysaccharides and antioxidant activity in different parts of Dendrobium ‘Sonia Jo Daeng’. Science, Engineering and Health Studies 2019; 13(2), 73-82.
[8] S Maksup and K Obsuwan. Flavonoid contents and antityrosinase activity in various parts of Dendrobium Sonia ‘Jo Daeng’ and Dendrobium ‘Khao Sanan’ at different growth stages. Acta Hortic 2020; 1298, 517-522.
[9] M Hassan, S Shahzadi and A Kloczkowski. Tyrosinase inhibitors naturally present in plants and synthetic modifications of these natural products as anti-melanogenic agents: A review. Molecules 2023; 28(1), 378.
[10] X Jia and L He. Unveiling the dual-edged roles of melanocytes: Key insights into skin photoaging and therapeutic avenues. Pharmacological Research 2025; 218, 107848.
[11] M Mucha, E Skrzydlewska and A Gegotek. Natural protection against oxidative stress in human skin melanocytes. Communications Biology 2025; 8(1), 1283.
[12] M Kanlayavattanakul, N Lourith and P Chaikul. Biological activity and phytochemical profiles of Dendrobium: A new source for specialty cosmetic materials. Industrial Crops and Products 2018; 120, 61-70.
[13] S Li, J Wang, L Zhang, Y Zheng, G Ma, X Sun and J Yuan. Preparation of Dendrobium officinale flower anthocyanin and extended lifespan in Caenorhabditis elegans. Molecules 2022; 27(23), 8608.
[14] M Han, Y Zhao, J Meng, J Yin and H Li. Analysis of physicochemical and antioxidant properties of Malus spp. petals reveals factors involved in flower color change and market value. Scientia Horticulturae 2023; 310, 111688.
[15] S Warinhomhoun, C Muangnoi, V Buranasudja, W Mekboonsonglarp, P Rojsitthisak, K Likhitwitayawuid and B Sritularak. Antioxidant activities and protective effects of dendropachol, a new bisbibenzyl compound from Dendrobium pachyglossum, on hydrogen peroxide-induced oxidative stress in HaCaT keratinocytes. Antioxidants 2021; 10(2), 252.
[16] K Thammasiri. Current status of orchid production in Thailand. Acta Hortic 2014; 1078, 25-33.
[17] C Macwan, H Patel and K Kalia. A comparative evaluation of in vitro antioxidant properties of Bamboo Bambusa arundinacea leaves extracts. Journal of Cell and Tissue Research 2010; 10(3), 2413.
[18] VL Singleton and JA Rossi. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. American journal of Enology and Viticulture 1965; 16(3), 144-158.
[19] UPN Kannaian, JB Edwin, V Rajagopal, SN Shankar and B Srinivasan. Phytochemical composition and antioxidant activity of coconut cotyledon. Heliyon 2020; 6(2), e03411.
[20] J Lee, RW Durst and RE Wrolstad. Determination of total monomeric anthocyanin pigment content of fruit juices, beverages, natural colorants, and wines by the pH differential method: Collaborative study. J AOAC Int 2005; 88(5), 1269-1278
[21] M DuBois, KA Gilles, JK Hamilton, PA Rebers and F Smith. Colorimetric method for determination of sugars and related substances. Analytical Chemistry 1956; 28(3), 350-356.
[22] GL Miller. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Analytical Chemistry 1959; 31(3), 426-428.
[23] M Leitão, B Ferreira, B Guedes, D Moreira, PA García, L Barreiros and P Correia. Screening of antioxidant effect of spontaneous and bioinoculated with Gluconobacter oxydans fermented papaya: A comparative study. Fermentation 2023; 9(2), 124.
[24] H Li, C Tang, Z Xu, X Liu and X Han. Effects of different light sources on the growth of non-heading Chinese cabbage (Brassica campestris L.). Journal of Agricultural Science 2012; 4(4), 262.
[25] TS Chang. An updated review of tyrosinase inhibitors. Int J Mol Sci 2009; 10(6), 2440-2475.
[26] M Tanaka, Y Kishimoto, E Saita, N Suzuki-Sugihara, T Kamiya, C Taguchi, K Iida and K Kondo. Terminalia bellirica extract inhibits low-density lipoprotein oxidation and macrophage inflammatory response in vitro. Antioxidants 2016; 5(2), 20.
[27] HC Nguyen, KH Lin, MY Huang, CM Yang, TH Shih, TC Hsiung, YC Lin and FC Tsao. Antioxidant activities of the methanol extracts of various parts of Phalaenopsis orchids with white, yellow, and purple flowers. Notulae Botanicae Horti Agrobotanici Cluj-Napoca 2018; 46(2), 457-465.
[28] C Chaiyasut, BS Sivamaruthi, N Pengkumsri, S Sirilun, S Peerajan, K Chaiyasut and P Kesika. Anthocyanin profile and its antioxidant activity of widely used fruits, vegetables, and flowers in Thailand. Asian Journal of Pharmaceutical and Clinical Research 2016; 9(6), 218-224.
[29] AR Zuo, HH Dong, YY Yu, QL Shu, LX Zheng, XY Yu and SW Cao. The antityrosinase and antioxidant activities of flavonoids dominated by the number and location of phenolic hydroxyl groups. Chinese Medicine 2018; 13(1), 51.
[30] N Sumaythachotphong, W Bundithya and N Potapohn. Flower color inheritance of Rhynchostylis gigantea (Lindl.) Ridl. Thai J. Agric. Sci. 2020; 53(4), 178-191.
[31] KE Le, S Bharadwaj, AK Sahoo, U Yadava and
SG Kang. Determination of tyrosinase-cyanidin-3-O-glucoside and (-/+)-catechin binding modes reveal mechanistic differences in tyrosinase inhibition. Sci Rep 2021; 11, 24494.
[32] C Monmai, JS Kim, JH Chin, S Lee and SH Baek. Inhibitory effects of polyphenol- and flavonoid-enriched rice seed extract on melanogenesis in melan-a Cells via MAPK signaling-mediated MITF downregulation. International Journal of Molecular 2023; 24(14), 11841
[33] PK Das, DH Shin, SB Choi, SD Yoo, G Choi and YI Park. Cytokinins enhance sugar-induced anthocyanin biosynthesis in Arabidopsis. Molecules and Cells 2012; 34(1), 93-101.
[34] C Solfanelli, A Poggi, E Loreti, A Alpi and P Perata. Sucrose-specific induction of the anthocyanin biosynthetic pathway in Arabidopsis. Plant Physiology 2006; 140(2), 637-646.
[35] N Smirnoff. Ascorbic acid metabolism and functions: A comparison of plants and mammals. Free Radical Biology and Medicine 2018; 122, 116-129
[36] MP Zia and I Alibaş. The effect of different drying techniques on color parameters, ascorbic acid content, anthocyanin and antioxidant capacities of cornelian cherry. Food Chem 2021; 364, 130358.
[37] S Tan, Y Miao, C Zhou, Y Luo, Z Lin, R Xie and W Li. Effects of hot air drying on drying kinetics and anthocyanin degradation of blood-flesh peach. Foods 2022; 11(11), 1596.
[38] F Wu, F Xu, W Liu, S Chen, H Luo, N Cheng, H Zhao and W Cao. A high-performance liquid chromatography with electrochemical detection method developed for the sensitive determination of ascorbic acid: Validation, application, and comparison with titration, spectrophotometric, and high-performance liquid chromatography with diode-array detection methods. Foods 2023; 12(16), 3100.
[39] A Torres de Pinedo, P Peñalver and JC Morales. Synthesis and evaluation of new phenolic-based antioxidants: Structure-activity relationship. Food Chem 2007; 103(1), 55-61.
[40] MS Brewer. Natural antioxidants: Sources, compounds, mechanisms of action, and potential applications. Comprehensive Reviews in Food Science and Food Safety 2011; 10(4), 221-247.
[41] N Tena, J Martín and AG Asuero. State of the art of anthocyanins: Antioxidant activity, sources, bioavailability, and therapeutic effect in human health. Antioxidants 2020; 9(5), 451.
[42] J Ávila-Román, R Talaverón, L Hernández-Cáceres, A de los Reyes, FJ Fernández-Arche, A González-Gallego, C García-Mauriño, ME Martín and M Motilva. Up-regulation of the Nrf2/HO-1 antioxidant pathway in macrophages by an extract from a new halophilic archaea isolated in Odiel saltworks. Antioxidants 2023; 12(5), 1080. doi:10.3390/antiox12051080.
[43] S Carpentieri, M Pagano, M Russo, A Carullo, E Laezza, E Cervelli, P Formisano, M Treppiccione, G Panzella, G Giarra and M Scognamiglio. Antioxidant and anti-inflammatory effects of extracts from pulsed electric field-treated artichoke by-products in lipopolysaccharide-stimulated human THP-1 macrophages. Foods 2022; 11(15), 2250. doi:10.3390/foods11152250.
[44] JP Ebanks, RR Wickett and RE Boissy. Mechanisms regulating skin pigmentation: The rise and fall of complexion coloration. International Journal of Molecular Sciences 2009; 10(9), 4066-4087.
[45] R Sarkar, S Chugh and VK Garg. Newer and upcoming therapies for melasma. Indian Journal of Dermatology Venereology and Leprology 2012; 78, 417.
[46] S Letchuman, HDT Madhuranga, BLNK Madhurangi, AD Premarathna and M Saravanan. Alkaloids unveiled: A comprehensive analysis of novel therapeutic properties, mechanisms, and plant-based innovations. Intelligent Pharmacy 2024; 3(4), 268-276.
[47] Z Habli, G Toumieh, M Fatfat, ON Rahal and H Gali-Muhtasib. Emerging cytotoxic alkaloids in the battle against cancer: Overview of molecular mechanisms. Molecules 2017; 22(2), 250.
[48] C Shen, H Guo, H Chen, Y Shi, Y Meng, J Lu, S Feng and H Wang. Identification and analysis of genes associated with the synthesis of bioactive constituents in Dendrobium officinale using RNA-Seq. Scientific Reports 2017; 7(1), 187.
[49] Y Hushcha, I Blo, L Oton-Gonzalez, GD Mauro, F Martini, M Tognon and MD Mattei. MicroRNAs in the Regulation of Melanogenesis. International Journal of Molecular Sciences 2021; 22(11), 6104.
[50] P Khongkarat, P Sadangrit, S Puthong, T Meemongkolkiat, P Phuwapraisirisan and C Chanchao. Anti-tyrosinase and anti-melanogenic effects of piperine isolated from Piper nigrum on B16F10 mouse melanoma cells. Heliyon 2024; 10(12), e33423.
[51] Q Yu and L Fan. Antityrosinase and antioxidant activity of asparagus and its inhibition on B16F10 melanoma cells before and after hydrothermal treatment. Food Bioscience 2021; 41, 101026.
[52] ZJ Cheng, GF Dai, JL Hsu, JJ Lin, WT Wu, CC Su and YJ Wu. Antimelanogenesis effect of methyl gallate through the regulation of PI3K/Akt and MEK/ERK in B16F10 melanoma cells. Evidence‐Based Complementary and Alternative Medicine 2022; 2022, 509.