Trends
Sci.
2026;
23(5):
12629
Terpene-Based Phytochemicals as a Novel Strategy for Biofilm Disruption and Enhanced Wound Healing
Thanusha Perera1,2,*, Satheesh Babu Natarajan1, Anandarajagopal Kalusalingam3,
Shudipta Choudhury4 and Kamaleswaran Ramachandran4
1Department of Pharmaceutics, School of Pharmacy, Lincoln University College, Selangor 47301, Malaysia
2Department of Science and Technology, Faculty of Applied Sciences, Lincoln Institute of Graduate Studies,
Colombo, Sri Lanka
3Department of Pharmaceutical Chemistry, School of Pharmacy, KPJ Healthcare University, Nilali,
Negeri Sembilan, Malaysia
4V3Bio Sdn. Bhd, Penang 14100, Malaysia
(*Corresponding author’s e-mail: [email protected])
Received: 14 November 2025, Revised: 21 December 2025, Accepted: 30 December 2025, Published: 10 January 2026
Abstract
Wound healing is a complex, multi-phase process involving hemostasis, inflammation, proliferation, and remodeling. Chronic wounds, including diabetic ulcers and pressure sores, often stall in the inflammatory phase due to persistent infections, oxidative stress, and impaired tissue regeneration. Microbial biofilms, particularly from Staphylococcus aureus and Pseudomonas aeruginosa, shield bacteria from host defenses and antibiotics, contributing to the majority of chronic infections and hospital-acquired infections. This review evaluates the antibiofilm and antimicrobial potential of terpene-based phytochemicals and examines their formulation into nanoemulsion-based delivery systems. Published studies were analyzed for mechanisms of biofilm disruption, bacterial inhibition, and enhancement of antimicrobial activity via nanoemulsions. Terpene phytochemicals disrupt biofilm architecture, inhibit bacterial proliferation, compromise cell membrane integrity, and interfere with quorum sensing and adhesion processes. When delivered in nanoemulsions, their solubility, stability, and bioavailability are improved, enhancing antimicrobial efficacy. These compounds also promote tissue repair and regeneration, addressing both infection control and wound healing. By simultaneously targeting biofilm-protected pathogens and supporting tissue regeneration, terpene-based phytochemicals offer a biocompatible, nature-inspired therapeutic strategy for chronic wound management. Their combined antibiofilm, antimicrobial, and tissue-regenerative actions underscore their potential as next-generation agents for advanced wound care.
Keywords: Wound healing, Chronic wounds, Biofilm disruption, Terpene-based phytochemicals, Antibiofilm agents, Nanoemulsions
Introduction
Wound healing is a complex process involving hemostasis, inflammation, proliferation, and maturation [1]. Blood arteries first narrow, and platelets clump together to stop blood loss while releasing cytokines to draw immune cells [2]. Acute wounds, often arising from surgical interventions or severe traumas, move through a systematic sequence of phases: haemostasis, inflammation, proliferation, and remodelling [3]. Vascular constriction and platelet aggregation take place first to stop blood loss, and then there is an inflammatory response in which neutrophils and macrophages remove debris and fight off any infections [4]. During the proliferative phase, fibroblasts produce extracellular matrix components, particularly collagen, which aids in the development of granulation tissue. Simultaneously, angiogenesis guarantees sufficient oxygenation and nutrition delivery to the rebuilding tissue [5]. The concluding remodelling phase entails collagen maturation and reorganisation, resulting in the restoration of tissue integrity [6].
In case of chronic wounds, including diabetic foot ulcers and pressure ulcers, present a significant global health challenge, affecting approximately 0.78% of the population, with prevalence rates ranging from 0.18% to 0.32% [7]. Chronic wounds, such as diabetic ulcers or pressure sores, exhibit a prolonged and often aberrant healing process [8]. These wounds frequently stall in the inflammatory phase, characterized by persistent infiltration of neutrophils and elevated levels of pro-inflammatory cytokines [9]. The chronic inflammatory milieu leads to excessive protease activity, degrading essential extracellular matrix components and growth factors necessary for tissue repair [10]. Moreover, sustained oxidative stress from reactive oxygen species further damages cellular structures and impedes healing. The imbalance between tissue degradation and repair mechanisms results in the failure to progress through the normal healing stages, thereby perpetuating the chronicity of the wound [11].
Biofilms, complex communities of microorganisms adhering to surfaces, represent a significant global health challenge due to their resilience and role in persistent infections [12] This biofilm matrix, composed of polysaccharides, proteins, lipids, and extracellular DNA (eDNA), provides structural integrity and protection to the bacterial community [13]. It is estimated that approximately 80% of all human bacterial infections involve biofilm formation [14]. Common medical devices such as catheters, prosthetic heart valves, and intrauterine devices are prone to biofilm colonization, leading to device-related infections that are challenging to treat [15]. Biofilm-associated infections are a significant concern in healthcare settings due to their resistance to antibiotics and the immune system. The National Institutes of Health (NIH) estimates that biofilms are associated with approximately 65% of all microbial infections and 80% of chronic infections [16]. In the context of hospital-acquired infections (HAIs), biofilms play a substantial role. According to a 2022 editorial in Frontiers in Medicine, up to 80% of all microbial infections and over 60% of nosocomial (hospital-acquired) infections are due to biofilms [17]. Regarding the economic impact, a 2009 report by the Centers for Disease Control and Prevention (CDC) estimated that the overall annual direct medical costs of HAIs to U.S. hospitals ranged from $28.4 to $33.8 billion. Given that a significant proportion of these infections are biofilm-associated, the additional healthcare costs attributable to biofilm-associated HAIs are substantial [18]. Notably, pathogens such as Pseudomonas aeruginosa and Staphylococcus aureus are frequently implicated in these biofilm-related infections, contributing to chronic conditions like cystic fibrosis lung infections and persistent wound infections [19,20].
The initial stage of biofilm formation involves the reversible attachment of planktonic bacteria to a surface, mediated by appendages such as pili or fimbriae [21]. In case of Acinetobacter baumannii, pili play a crucial role in this attachment phase [22]. Subsequently, bacteria produce eDNA, which is essential for stabilizing the biofilm matrix and facilitating cell-to-cell adhesion [23]. Research led by Whitchurch et al. demonstrated that eDNA is a vital structural component in biofilm development, promoting self-organization within bacterial communities [24]. As the biofilm matures, bacteria undergo phenotypic changes, including altered gene expression and reduced metabolic activity, contributing to increased resistance to antimicrobial agents [25]. Bacteria within biofilms exhibit a significantly higher resistance to antibiotics compared to their free-floating (planktonic) counterparts. This increased resistance can range from 10 to 1,000 times greater, posing substantial challenges in treating biofilm-associated infections [26,27].
Phytochemicals, naturally occurring compounds in plants, have garnered attention for their potential to disrupt microbial biofilms. Research indicates that certain phytochemicals, including flavonoids, tannins, terpenoids, alkaloids, and essential oils, exhibit antibiofilm activity by inhibiting bacterial adhesion, disrupting quorum sensing, and degrading the biofilm matrix [28,29]. Terpenes, a diverse class of naturally occurring organic compounds, have garnered significant attention for their potential in antimicrobial and immune-modulatory therapies [30]. One notable example is totarol, a diterpene isolated from the heartwood of Podocarpus totara, a conifer native to New Zealand. Totarol has demonstrated potent antimicrobial properties against various microorganisms, including gram-positive bacteria such as Staphylococcus aureus and Mycobacterium tuberculosis [31]. Its mechanism of action is believed to involve the inhibition of bacterial cytokinesis by disrupting the assembly dynamics of the FtsZ protein, essential for bacterial cell division [32]. Another example is Berberine, an isoquinoline alkaloid from Berberis species, which exhibits potent antimicrobial activity against gram-positive bacteria like Staphylococcus aureus. It disrupts bacterial DNA synthesis and membrane integrity while inhibiting efflux pumps, enhancing antibiotic efficacy [33].
Recent advancements in eradicating biofilms have focused on innovative strategies that disrupt the structural integrity and resilience of these microbial communities [34]. One promising approach involves the development of light-sensitive mesoporous silica nanoparticles (MSNs) with photothermal properties. These nanoparticles, when functionalized with nitric oxide donors and loaded with antibiotics like levofloxacin, can effectively penetrate biofilms [35]. Upon near-infrared irradiation, they induce localized heating and enhanced nitric oxide release, leading to a significant reduction in biofilm viability, as demonstrated by a 90% decrease in Staphylococcus aureus biofilms [36]. Additionally, the exploration of natural antimicrobial proteins, such as those derived from the haemolymph of the Sydney rock oyster, has shown potential. These proteins exhibit bactericidal activity against pathogens like Streptococcus pneumoniae and can disrupt biofilm structures, thereby improving the effectiveness of conventional antibiotics [37].
Nanoemulsions (NEs) of essential oils (EOs), when formulated with an appropriate delivery system, offer a promising strategy to overcome these challenges. NEs are nanoscale emulsions consisting of oil and water phases, stabilized by surfactants, with particle sizes typically below 200 nm [38]. Among these, oil-in-water (O/W) NEs serve as exceptional carriers for EOs, owing to their remarkable ability to solubilize substantial amounts of essential oils while effectively shielding them from evaporation, hydrolysis, and degradation [39]. A study evaluating cinnamaldehyde nanoemulsion (CAN) revealed its potent antibiofilm activity against Listeria monocytogenes on food-contact surfaces. With stable nanoscale particles (< 200 nm), CAN demonstrated a minimum inhibitory concentration of 0.5 mg/mL and exerted a concentration- and time-dependent bactericidal effect, achieving over 90% biofilm inhibition [40]. This systematic review aims to evaluate the antibiofilm efficacy of terpene-based phytochemicals and their delivery strategies in chronic wound management.
Methodology
The literature included in this review was compiled using a structured approach to identify peer-reviewed studies on microbial biofilms in chronic wounds and the antibiofilm, antimicrobial, and wound-healing potential of terpene-based phytochemicals, including nanoemulsion-based delivery systems. Searches were conducted in PubMed, Scopus, Web of Science, and Google Scholar. Studies were selected based on their relevance to: (i) Biofilm-associated infections in chronic wounds, (ii) the antimicrobial and antibiofilm activity of terpene-based phytochemicals (e.g., totarol, berberine), and (iii) formulation strategies improving solubility, stability, and bioavailability. Original research articles and high-quality review papers providing mechanistic, in vitro, or in vivo evidence were prioritized. Non-peer-reviewed sources, conference abstracts, and studies unrelated to chronic wound biofilms were excluded.
Limitations include restriction to English-language publications, potential publication bias, and reliance on database indexing, which may have resulted in omission of some relevant studies. This methodology does not follow a systematic or scoping review framework but provides a transparent summary of the literature selection process.
Study Selection and Quality Assessment: Two independent reviewers screened titles and abstracts. Full-text studies were assessed for eligibility, and disagreements were resolved by a third reviewer. Study quality was evaluated using appropriate tools (e.g., Cochrane risk of bias, Newcastle-Ottawa Scale). A PRISMA 2020 flow diagram was used to document study selection.
Anti-biofilm mechanism
Planktonic bacteria connect to a surface by weak physicochemical interactions to initiate the dynamic, multistage process of biofilm development (Figure 1). After then, a microcolony forms, during which the connected cells multiply and start interacting with one another as well as producing extracellular polymeric material (EPS) [41]. The biofilm moves into the maturity stage as it develops into a sophisticated, 3-dimensional structure immersed in an EPS matrix that shields bacteria from external stressors and promotes nutrient exchange. Ultimately, during the dispersion stage, certain cells separate from the mature biofilm in response to internal or external stimuli, returning to a planktonic condition and facilitating the colonisation of new surfaces [42,43].
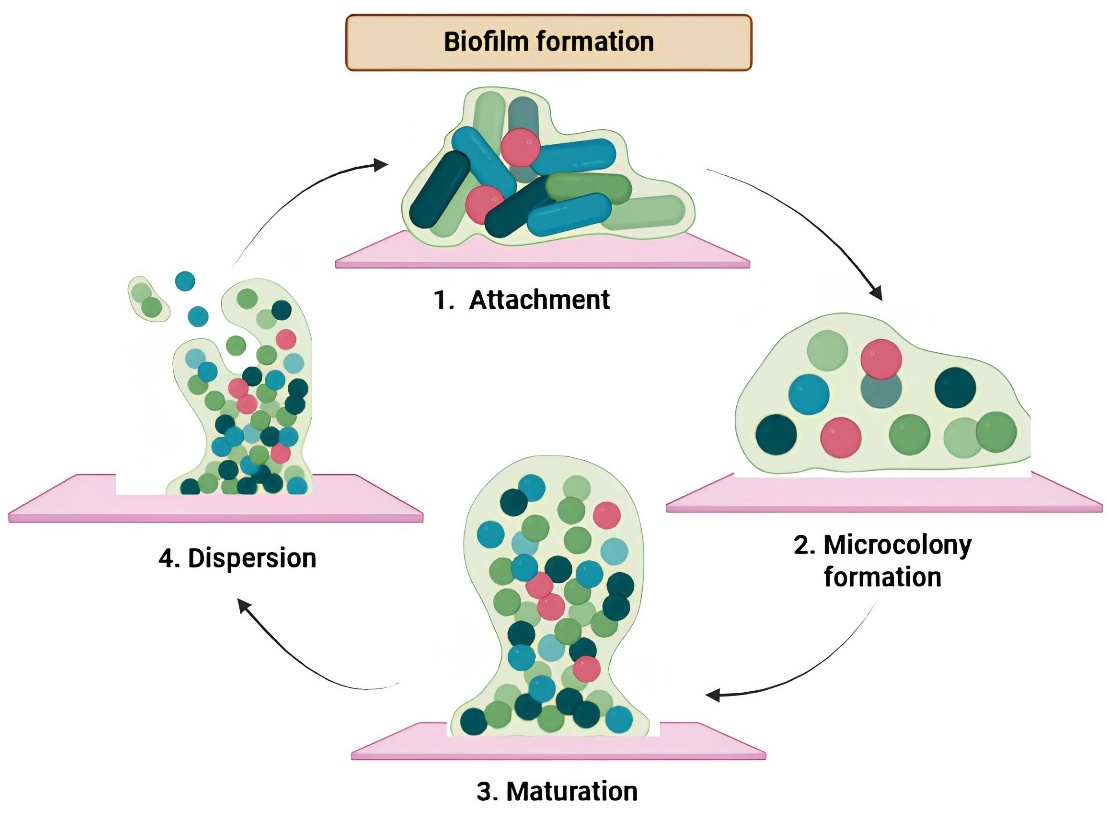
Figure 1 Mechanisms of Biofilm Formation and Strategies for Biofilm Inhibition. A) Schematic representation of the mechanism of biofilm formation. The process involves sequential stages: (1) initial attachment of planktonic cells to a surface, (2) microcolony formation, (3) maturation of the biofilm structure and (4) dispersion of cells to colonize new environments.
Chemical composition and their functions in biofilm
Biofilms are intricate microbial communities that attach to surfaces and are involved in a self-generated extracellular matrix, the chemical composition of which is crucial for biofilm development, stability, and usefulness [44]. The main constituents of the biofilm matrix include proteins, lipids, polysaccharides, eDNA, and occasionally extracellular enzymes. All of these elements help the biofilm withstand environmental stressors and antimicrobial therapies [45]. Structural stability is provided by polysaccharides such cellulose, alginate, and PSL, which promote cell-to-cell attachment and shield the biofilm from shear forces and desiccation [46]. In addition to mediating attachment to surfaces, proteins such as adhesins and hydrolases also help in nutrition uptake and host immune response defence. Environmental DNA, often derived from lysed cells, serves as both a structural scaffold and a medium for gene transfer and horizontal gene exchange, hence enhancing the genetic diversity of bacteria inside biofilms [47,48]. Lipids and lipopolysaccharides augment the hydrophobic characteristics of the matrix, hence increasing resistance to detergents and antibiotics [49]. The biofilm’s 3-dimensional architecture generates microenvironments with differing oxygen, pH, and nutrition levels, facilitating niche specialisation among microbial subpopulations, hence augmenting biofilm resistance and pathogenicity [50].
Biofilm formation in gram-positive and gram-negative bacteria:
Biofilm formation is a conserved, multistep process involving initial surface adhesion, extracellular polymeric substance (EPS) production, quorum sensing mediated regulation, and biofilm maturation, ultimately conferring enhanced tolerance to antimicrobial agents and host immune responses [23]. While the molecular regulators differ between Gram-positive and Gram-negative bacteria, the functional outcomes remain largely similar.
In Gram-positive bacteria, such as Staphylococcus aureus, biofilm development is mediated by surface-associated adhesion proteins that facilitate attachment to host tissues or abiotic surfaces, as illustrated in Figure 2 [51]. This is followed by the secretion of an EPS matrix composed primarily of polysaccharides, proteins, and extracellular DNA, which stabilises the biofilm architecture and enhances resistance to external stresses [23]. Biofilm maturation and dispersal are regulated by the agr quorum-sensing system, in which AgrD-derived autoinducing peptide (AIP), processed by AgrB, activates the AgrC–AgrA 2-component system, leading to RNAIII expression [52-55].
In Gram-negative bacteria, including Pseudomonas aeruginosa, surface attachment is typically facilitated by pili or fimbriae (Figure 3). Biofilm stability is maintained through species-specific EPS components such as alginate, Pel, and Psl polysaccharides [23,56]. Quorum sensing in these organisms primarily involves N-acyl homoserine lactone (AHL) signalling pathways, which regulate genes associated with EPS synthesis, motility, and biofilm maintenance by activating LuxR-type transcriptional regulators once a critical cell density is reached [57]. Additional regulatory mechanisms, including diffusible signal factors (DSFs) [58,59] and the Type VI secretion system (T6SS), further modulate interbacterial interactions, competition, and biofilm dynamics [60].
Despite these mechanistic differences, biofilms formed by both Gram-positive and Gram-negative bacteria exhibit reduced antimicrobial penetration, altered metabolic states, and enhanced persistence, thereby contributing to chronic and recurrent infections and posing significant challenges to effective clinical management [54,55,57].
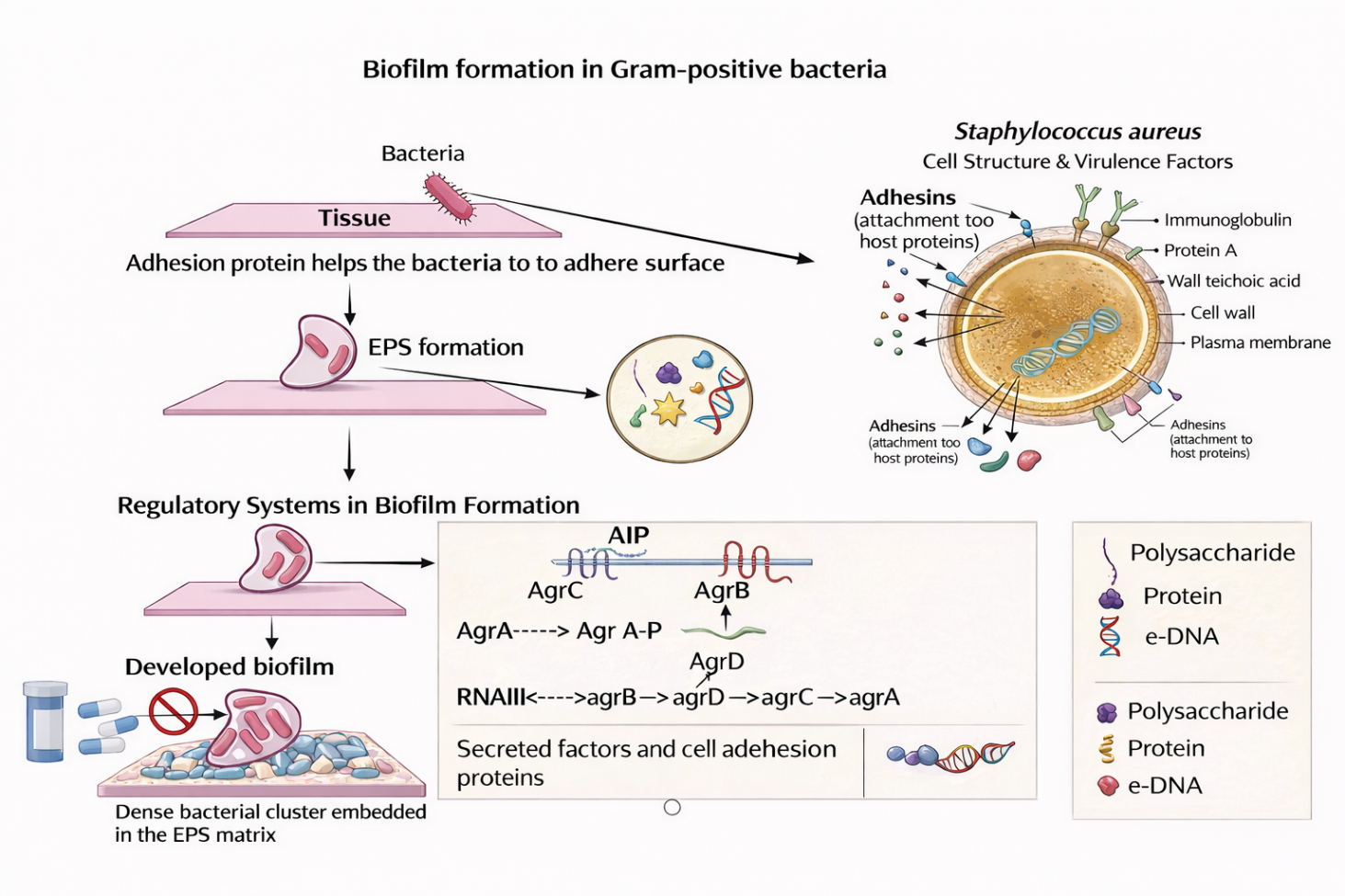
Figure 2 Schematic representation of biofilm formation in Gram-positive bacteria, highlighting surface adhesion, EPS matrix development, quorum sensing regulation, and biofilm maturation leading to antimicrobial resistance.
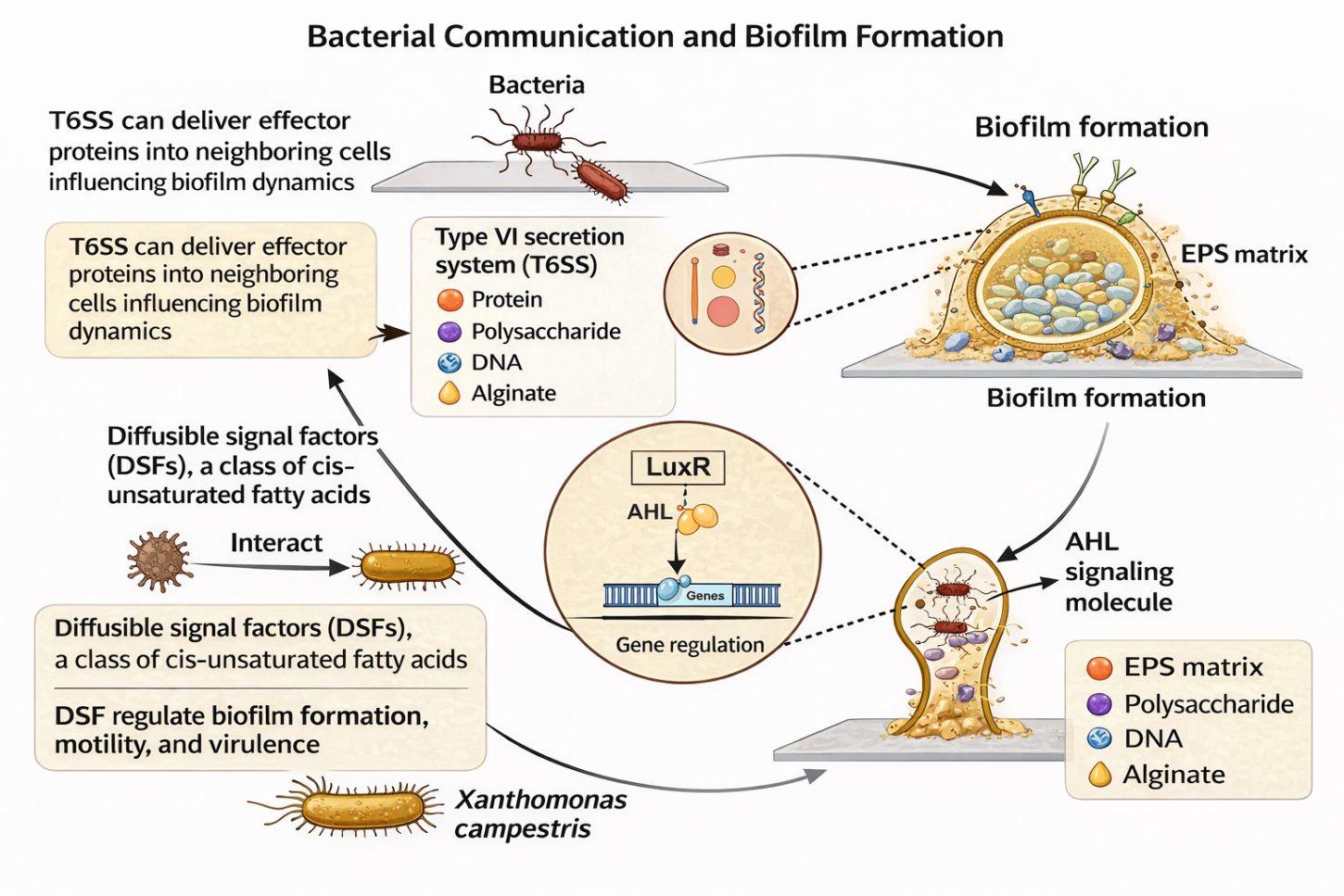
Figure 3 Overview of biofilm formation in Gram-negative bacteria, illustrating surface attachment, EPS synthesis, quorum sensing-mediated gene regulation, and biofilm stabilisation mechanisms.
Biofilm producing bacteria and infections associate with biofilms
Table 1 below outlines major bacterial species known for biofilm formation, along with their associated infections and key pathogenic traits. It highlights the structural and functional characteristics of biofilms that contribute to disease persistence. The mechanisms by which these bacteria evade host defenses and resist antibiotics are summarized. Treatment challenges, including reduced drug penetration and altered bacterial metabolism within biofilms, are also noted. Relevant references are provided to support each entry.
Table 1 Biofilm-producing bacteria and associated infections.
Microorganism |
Major Infections |
Key Biofilm / Virulence Features |
Clinical Challenges |
References |
Pseudomonas aeruginosa |
Chronic wounds, CF, UTI, pneumonia |
Alginate production, quorum sensing, eDNA |
High antibiotic resistance; poor drug penetration |
[61-63] |
Staphylococcus aureus |
Endocarditis, osteomyelitis, device-related infections |
Toxins, adhesion proteins, dense biofilms |
MDR and immune evasion in biofilms |
[64-66] |
Escherichia coli |
UTI, sepsis, wound infections |
Fimbriae, curli fibers, EPS |
Persistent catheter-associated infections |
[67-69] |
Streptococcus mutans |
Dental caries, oral infections |
Glucan synthesis, acid production |
Recurrent biofilm formation |
[70-72] |
Candida albicans |
Mucosal and bloodstream infections |
Yeast–hyphal transition, ECM production |
Antifungal resistance in biofilms |
[73-75] |
Acinetobacter baumannii |
VAP, wound and bloodstream infections |
Desiccation tolerance, strong biofilms |
Extreme MDR in hospital settings |
[76-78] |
Klebsiella pneumoniae |
Pneumonia, UTI, sepsis |
Capsule, fimbriae, EPS |
Persistent biofilms and carbapenem resistance |
[79,80] |
Enterococcus faecalis |
UTI, endocarditis, wounds |
ECM proteins, aggregation |
Vancomycin-resistant biofilms |
[81-83] |
Mycobacterium tuberculosis |
Pulmonary tuberculosis |
Dormancy, granuloma-associated biofilms |
Treatment persistence and relapse |
[84,85] |
Helicobacter pylori |
Gastritis, ulcers, gastric cancer |
Urease activity, motility, adhesion |
Reduced eradication by antibiotics |
[86,87] |
Proteus mirabilis |
UTI, catheter-associated infections |
Urease, swarming motility |
Biofilm-mediated UTI complications |
[88,89] |
Terpene-based phytochemicals and antibiofilm formation
Terpenes are a vast and varied category of chemical molecules synthesised by several plants, especially conifers, as well as some insects. They constitute the primary constituents of resin and turpentine derived from resin [90]. The amount of isoprene units that terpenes contain each of which has 5 carbon atoms determines their classification [91]. Table 2 presents the classifications of terpenes and their antibiofilm efficacy against bacteria. Monoterpenes are composed of 2 isoprene units and possess the chemical formula C10H16. They are the terpenes that are most volatile and are frequently in charge of giving many plants their unique fragrances [92]. Monoterpenes are present in the essential oils of several plants and are utilised in the manufacture of flavours, perfumes, and aromatherapy products. Examples are pinene, which is found in pine trees, and limonene, which is found in citrus fruits [93]. Sesquiterpenes have the chemical formula C15H24 because they are made up of 3 isoprene units. In many different plants and essential oils, they are less volatile than monoterpenes [91]. The ability of sesquiterpenes to reduce inflammation and cancer is well-known. The sesquiterpene lactone artemisinin, for example, is a powerful antimalarial medication derived from Artemisia annua [94]. Diterpenes include 4 isoprene units and possess the chemical formula C20H32. They are present in resins and serve as building blocks for physiologically significant substances including phytol, retinol, and retinal [95]. The Pacific yew tree (Taxus brevifolia) produces the diterpene taxol, a well-known chemotherapeutic drug used to treat a variety of malignancies [96]. Six isoprene units make up triterpenes, which have the chemical formula C30H48 [91]. White birch bark (Betula pubescens) contains a triterpene called betulinic acid, which has showed promise in cancer treatment [97].
Table 2 Classification and biological activities of terpenes.
Terpene |
Class |
Natural Source |
Target Microorganisms |
Key Antibiofilm Mechanisms |
Terpene |
References |
Limonene |
Monoterpene |
Citrus fruits, peppermint |
Staphylococcus aureus, Pseudomonas aeruginosa |
Inhibits quorum sensing and EPS production, reducing biofilm formation |
Limonene |
[98] |
Carvacrol |
Monoterpenoid |
Oregano, thyme |
Escherichia coli, Listeria monocytogenes, Candida albicans |
Disrupts cell membranes, reduces adhesion and motility, degrades EPS |
Carvacrol |
[99,100] |
α-Pinene |
Monoterpene |
Pine trees, rosemary |
Streptococcus mutans, Acinetobacter baumannii |
Downregulates EPS-related genes; shows synergy with antibiotics |
α-Pinene |
[101,102] |
β-Caryophyllene |
Sesquiterpene |
Cannabis, cloves |
Streptococcus pneumoniae, Helicobacter pylori |
Modulates bacterial signaling pathways, reducing biofilm viability |
β-Caryophyllene |
[103] |
Geraniol |
Monoterpenoid |
Geranium, lemongrass |
Staphylococcus aureus, Staphylococcus epidermidis |
Alters membrane integrity and surface hydrophobicity, limiting adhesion |
Geraniol |
[104,105] |
Eucalyptol (1,8-Cineole) |
Monoterpenoid |
Eucalyptus oil |
Streptococcus pyogenes, Pseudomonas aeruginosa |
Disrupts quorum sensing, reduces EPS synthesis, enhances antibiotic penetration |
Eucalyptol (1,8-Cineole) |
[106,107] |
Thymol |
Monoterpenoid |
Thyme, oregano |
Pseudomonas aeruginosa, Streptococcus mutans, Xanthomonas oryzae |
Damages membranes and suppresses biofilm-associated gene expression |
Thymol |
[108,109] |
Farnesol |
Sesquiterpenoid |
Citrus fruits, neroli oil |
Candida albicans, Staphylococcus aureus |
Inhibits hyphal development, interferes with quorum sensing and virulence |
Farnesol |
[110,111] |
Linalool |
Monoterpenoid |
Lavende, coriander |
Bacillus amyloliquefaciens, E. coli, C. albicans |
Increases membrane permeability, reduces adhesion; synergistic with antibiotics |
Linalool |
[112,113] |
Terpinen-4-ol |
Monoterpenoid |
Tea tree oil |
Propionibacterium acnes, Staphylococcus aureus |
Disrupts membranes, reduces aggregation, enhances antiseptic efficacy |
Terpinen-4-ol |
[114,115] |
Terpenes as a multi-pathogen biofilm disruptor in chronic wounds
Terpenes, a varied group of chemical molecules synthesised by several plants, have attracted interest for their possible function in regulating polymicrobial biofilms [116]. These biofilms, consisting of many bacteria species, offer considerable issues in clinical environments due to their increased resistance to antimicrobial treatments [117]. Table 3 presents a comparative review of diverse disorders linked to polymicrobial biofilms, emphasising the related pathogens, biofilm attributes, individual terpenes examined, and their antibiofilm effectiveness.
Table 3 Antibiofilm potential of terpenes against polymicrobial infections: a comparative analysis across diseases.
Disease / Condition |
Pathogens |
Biofilm Impact |
Terpenes Evaluated |
Antibiofilm Outcome |
Antibiofilm Mechanism & Efficacy |
References |
Chronic otitis media |
H. influenzae, S. pneumoniae, M. catarrhalis |
Persistent middle-ear biofilms |
Tea tree oil |
Reduced biofilm formation and antibacterial activity (in vitro) |
Disrupts biofilm EPS, reduces bacterial adhesion, antibacterial activity |
[118,119] |
Cystic fibrosis lung infection |
P. aeruginosa, S. aureus |
Dense multispecies respiratory biofilms |
Eucalyptus oil |
Inhibited biofilm formation and disrupted mature biofilms |
Disrupts quorum sensing, reduces EPS synthesis, enhances antibiotic penetration |
[106,120] |
Chronic wound infections |
S. aureus, P. aeruginosa, E. faecalis |
Delayed healing, high resistance |
Oregano oil |
Strong antibacterial and antibiofilm effects |
Compromises bacterial membranes, reduces adhesion, disrupts biofilm formation |
[121,122] |
Dental caries |
S. mutans, C. albicans |
Synergistic cariogenic biofilms |
Menthol |
Reduced biofilm biomass and cell viability |
Reduces biofilm biomass, inhibits adhesion, antibacterial activity |
[123,124] |
Urinary tract infections |
E. coli, E. faecalis |
Catheter-associated biofilms |
Thymol |
Suppressed biofilm formation and eradicated mature biofilms |
Disrupts membranes, inhibits EPS production, eradicates mature biofilms |
[108,125] |
CAUTIs |
E. coli, E. faecalis, P. mirabilis |
Recurrent catheter biofilms |
Carvacrol |
Inhibited formation and eliminated established biofilms |
Reduces adhesion and motility, disrupts EPS, eliminates existing biofilms |
[126] |
Gastric ulcers |
Helicobacter pylori |
Mucosal biofilms increase persistence |
Curcumin, α-pinene |
Reduced biofilm formation and virulence |
Inhibits quorum sensing, reduces EPS, decreases pathogenicity |
[127,128] |
Clostridioides difficile infection |
C. difficile |
Biofilm-protected spores |
Eugenol, thymol |
Significant antibacterial and antibiofilm activity |
Disrupts EPS, inhibits biofilm formation, antibacterial effect |
[129,130] |
Acne vulgaris |
Cutibacterium acnes |
Sebaceous biofilms |
Tea tree oil |
Disrupted biofilms and reduced bacterial load |
Disrupts membranes, reduces aggregation, decreases bacterial load |
[119,131] |
Periodontitis / gingivitis |
P. gingivalis, T. forsythia, T. denticola |
Gingival pocket biofilms |
Thymol, eugenol |
Inhibited plaque formation and bacterial adhesion |
Inhibits bacterial adhesion, reduces plaque biofilm formation |
[132-134] |
Osteomyelitis |
S. aureus, S. epidermidis |
Bone-associated chronic biofilms |
Carvacrol |
Disrupted biofilms and enhanced antibiotic susceptibility |
Disrupts biofilm matrix, enhances antibiotic susceptibility |
[135] |
Bacterial vaginosis |
G. vaginalis, A. vaginae |
Persistent vaginal biofilms |
Eugenol, thymol |
Disrupted biofilms and reduced adhesion |
Disrupts adhesion, inhibits biofilm formation |
[136] |
Pelvic inflammatory disease |
N. gonorrhoeae, C. trachomatis |
Protective reproductive tract biofilms |
Geraniol, carvacrol |
Inhibited pathogen growth and biofilm formation |
Reduces bacterial adhesion, inhibits biofilm formation |
[137] |
Synergistic effects of terpenes with antibiotics and antimicrobial peptides
Terpenes can significantly enhance the efficacy of conventional antibiotics and antimicrobial peptides (AMPs), allowing dose reduction and potentially limiting adverse effects. Their primary mode of action involves integration into microbial cell membranes, leading to increased membrane permeability and enhanced intracellular uptake of antibiotics or AMPs. In addition, certain terpenes inhibit bacterial efflux pumps, promoting intracellular retention of antimicrobial agents and reducing resistance development [30,138,139]. Monoterpenes such as α-terpineol and terpinen-4-ol exhibit strong antibacterial activity, particularly against Gram-negative bacteria, by disrupting membrane integrity, reducing membrane potential, and inducing leakage of intracellular macromolecules [140,141]. Beyond membrane disruption, terpenes can interfere with microbial metabolic pathways. For example, the tropolone monoterpene hinokitiol exerts antifungal activity by chelating intracellular iron, thereby inhibiting mitochondrial respiration and ATP synthesis, which are essential for fungal survival [142].
These synergistic interactions frequently result in reduced minimum inhibitory concentrations (MICs) of antibiotics [143]. Compounds such as citral and borneol have also demonstrated enhanced antibiofilm activity when combined with antibiotics by increasing membrane porosity and promoting bacterial cell lysis, thereby improving treatment efficacy against biofilm-associated infections [144]. Similarly, phenolic terpenes including thymol and carvacrol have shown enhanced antibacterial activity when combined with antibiotics such as tetracycline and penicillin, particularly against Staphylococcus aureus and Escherichia coli [145-148]. Synergy has also been reported between terpenes and AMPs. Cinnamaldehyde, the major component of cinnamon oil, exhibits enhanced bactericidal activity when combined with AMPs such as cecropin by accelerating membrane disruption and bacterial cell death [149]. In fungal infections, essential oils rich in terpenes, such as those from Lippia alba, demonstrate strong synergistic effects with conventional antifungal agents. Notably, combinations with ketoconazole showed potent fungicidal activity against Trichophyton rubrum and Candida species [150,151].
Synergistic effects of Terpenes with antibiotics on wound healing
In wound healing, terpene-antibiotic combinations are particularly valuable due to their dual antimicrobial and tissue-protective effects. Terpenes can suppress bacterial growth, inhibit biofilm formation, and modulate inflammatory responses, all of which are critical factors in chronic wound infections [152]. For instance, terpinen-4-ol has been shown to reduce bacterial burden and inflammation when used alongside antibiotics, thereby accelerating wound closure [153]. At the molecular level, this synergy involves modulation of inflammatory signaling pathways, including inhibition of NF-κB and NLRP3 inflammasome activation, as well as enhancement of antioxidant defenses through regulation of enzymes such as superoxide dismutase and glutathione-S-transferase [154]. Furthermore, incorporation of terpenes into wound dressings has been reported to improve antibiotic bioavailability and sustain drug release, resulting in improved therapeutic outcomes in both acute and chronic wounds [155].
Advances in the drug delivery system for wound healing
Significant clinical difficulties can arise from wound infections, especially in chronic wounds such pressure sores, diabetic ulcers, and venous leg ulcers [156]. These infections are frequently exacerbated by variables such as compromised blood flow, extended inflammation, and bacterial biofilm development, which impede proper healing and elevate the risk of systemic infections [157]. Conventional wound treatment techniques, such as topical antibiotics and dressings, frequently prove inadequate due to challenges such as restricted medication penetration, brief residence time, and inadequate control over drug release [158]. Traditional drug delivery systems (DDS) for wound healing, including ointments, creams, and hydrocolloid dressings, generally provide therapeutic chemicals by passive diffusion [159]. Although these systems are straightforward to implement and economically viable, they encounter many limitations: variable drug release patterns, diminished effectiveness in chronic wound conditions, and difficulties in sustaining appropriate drug concentrations at the wound site for prolonged durations [160]. These mechanisms are also frequently unable to handle the intricate pathophysiology of chronic wounds, including immunological dysfunction and compromised angiogenesis [161]. To address these issues, sophisticated DDS have been created, providing superior control over drug release and greater therapeutic results. Nanoparticles, liposomes, and solid lipid nanoparticles facilitate targeted distribution and sustained release of active drugs, improving bioavailability and minimising systemic adverse effects [162]. Microneedle devices provide painless, regulated medication administration straight to the skin layer, enhancing patient adherence and therapeutic effectiveness [163]. Hydrogels and electrospun nanofibers provide moist wound environments that promote healing and function as transporters for antibacterial agents and growth factors [164]. Moreover, scaffold-mediated delivery methods, utilising natural and synthetic polymers, facilitate tissue regeneration by offering structural support and enabling localised drug release. These sophisticated systems not only overcome the shortcomings of traditional DDS but also provide personalised and adaptive treatment solutions customised to the unique requirements of chronic wound patients [165].
Figure 4 thoroughly depicts the antimicrobial mechanisms of nanoparticles against bacterial cells, emphasising oxidative stress induced by the nanoparticle-mediated generation of reactive oxygen species (ROS), resulting in metabolic enzyme dysfunction and disruption of the electron transport chain [166]. The accumulation of nanoparticles in the bacterial microenvironment leads to membrane destabilisation through hole formation and a reduction in membrane potential, hence exacerbating cellular distress [167]. Moreover, macromolecular degradation resulting from nanoparticle absorption is illustrated by direct interference with genetic material and ribosomal activity, resulting in DNA damage, suppression of protein synthesis, and disturbances in quorum sensing (QS) pathways, ultimately compromising bacterial signalling, biofilm development, and virulence [168].
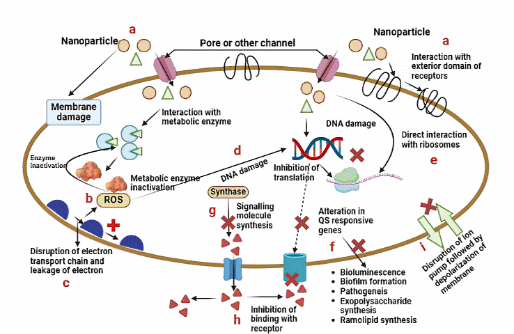
Figure 4 Mechanisms of antibacterial activity caused by metal nanoparticles: Disruptions in cellular processes and quorum sensing. This graphic depicts many antibacterial processes of nanoparticles targeting bacterial cells. Nanoparticles induce membrane damage by creating holes or compromising membrane integrity, resulting in the release of cellular contents. (a) They engage with metabolic enzymes, frequently producing reactive oxygen species (ROS), which deactivate these enzymes and disrupt metabolism. (c) This results in the disruption of the electron transport pathway, leading to energy depletion and oxidative stress. (d) Nanoparticles induce DNA damage, either directly or through reactive oxygen species (ROS). (e)Inhibition of translation by the alteration of ribosomal function. (f) They modify quorum sensing (QS) sensitive genes, diminishing bioluminescence, biofilm development, pathogenicity, and the production of exopolysaccharides and rhamnolipids. (g) Nanoparticles impede the production of signalling molecules by obstructing QS synthase enzymes. (h) They inhibit these molecules from attaching to receptors, hence affecting quorum sensing signals and bacterial communication. (i) They induce disruption of the ion pump and membrane depolarisation.
Nanocarriers for terpene delivery
Nanocarriers have become essential systems for the efficient transport of terpenes, improving their solubility, stability, and bioavailability [169]. Liposomes, ethosomes, polymeric nanoparticles, and solid lipid nanoparticles (SLNs) have attracted considerable interest, as seen in Figure 5. Liposomes are spherical vesicles consisting of one or more phospholipid bilayers that encase an aqueous core, enabling the encapsulation of both hydrophilic and lipophilic molecules. Their biocompatibility and structural adaptability render them appropriate for terpene delivery [170,171]. Ethosomes are a specialised type of liposomes that contain elevated levels of ethanol, which improves their flexibility and ability to penetrate biological membranes, making them particularly advantageous for the transdermal administration of terpenes [172]. The incorporation of ethanol compromises the lipid bilayers of the stratum corneum, enabling enhanced penetration into the skin [173]. Polymeric nanoparticles, often measuring between 10 and 1,000 nm, are fabricated from biodegradable polymers like poly(lactic-co-glycolic acid) (PLGA) or chitosan [174]. These nanoparticles can encapsulate terpenes, safeguarding them from degradation and facilitating regulated release patterns. Their surface can be altered to increase targeted specificity, therefore boosting therapeutic effectiveness [175]. Nonetheless, issues including possible cytotoxicity and the requirement for intricate production procedures demand meticulous evaluation in their use [176]. Solid lipid nanoparticles (SLNs) comprise solid lipids stabilised by surfactants, creating a matrix that may incorporate lipophilic chemicals such as terpenes. SLNs can improve the bioavailability of terpenes by promoting lymphatic absorption and provide protection against environmental degradation [177]. They provide benefits like enhanced medication stability, regulated release, and the practicality of large-scale manufacturing. However, challenges such as restricted drug-loading capacity and possible polymorphic transitions during storage necessitate optimisation to guarantee consistent performance [178,179].
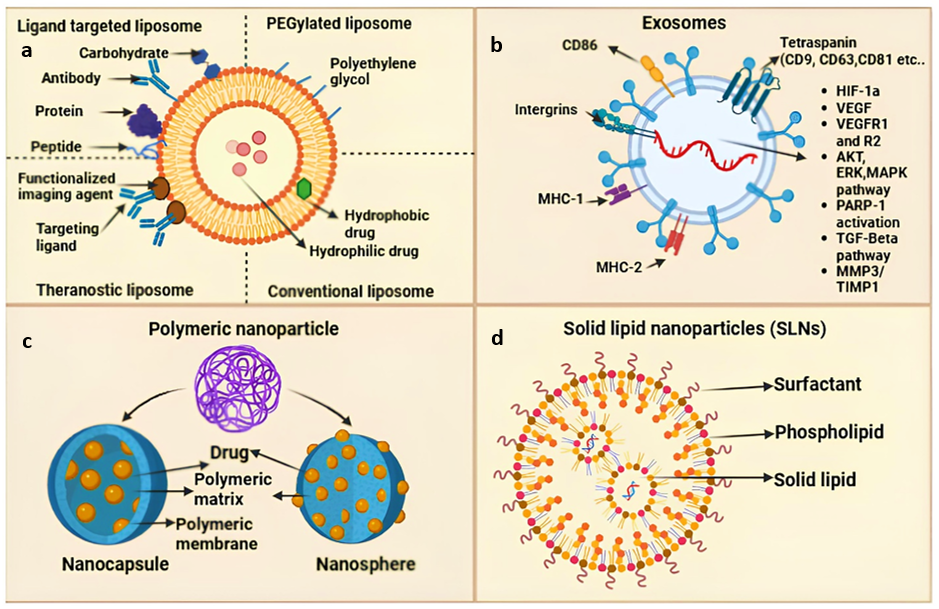
Figure 5 Advanced Drug Delivery Nanocarrier Systems: Solid Lipid Nanoparticles, Liposomes, Exosomes, and Polymeric Nanoparticles. This graphic provides a comprehensive overview of sophisticated nanocarrier systems employed in medication delivery. (a) Liposomes can take many different forms: theranostic liposomes combine therapeutic and diagnostic agents; ligand-targeted liposomes are functionalised with ligands, peptides, or antibodies for targeted delivery; PEGylated liposomes are coated with polyethylene glycol for increased stability; and conventional liposomes carry both hydrophilic and hydrophobic drugs. (b) Proteins, lipids, and RNAs are found in exosomes, which are naturally occurring vesicles that facilitate intercellular communication through pathways including VEGF, MAPK, and TGF-β as well as surface indicators like tetraspanins. (c) Polymeric nanoparticles include core-shell nanocapsules that provide controlled release and nanospheres with the medication distributed throughout the matrix. (d) Biocompatibility, medication stability, and effective administration are offered by solid lipid nanoparticles (SLNs), which are made up of solid lipids, phospholipids, and surfactants.
Nanoemulsion based drug delivery system
In the fight against biofilms in wound healing, nanoemulsion-based drug delivery devices have shown great promise (Figure 6) [180]. Due to their nanoscale droplet size, NEs have improved features including increased surface area, better solubility of hydrophobic medicines, and efficient penetration of biofilms. Antimicrobial drugs that are hydrophilic or lipophilic can be encapsulated in these systems, allowing for their targeted and prolonged release at the wound site [181,182]. For example, biodegradable NEs loaded with 2 antimicrobials have shown synergistic benefits, efficiently breaking down biofilms and encouraging tissue regeneration [183]. NEs can be designed to react to certain stimuli in the wound microenvironment, facilitating the regulated release of therapeutic medicines [184]. The use of nanoemulsion-based delivery methods in wound care boosts antibacterial effectiveness while facilitating the natural wound healing process by diminishing inflammation and encouraging re-epithelialization [185]. Nonetheless, issues including the durability of NEs, possible cytotoxicity, and the scalability of production must be resolved to fully harness their therapeutic potential. Current research aims to optimise formulations and investigate innovative biocompatible materials to address these constraints and enhance the use of nanoemulsion-based therapeutics in wound care [186].
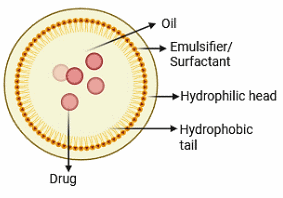
Figure 6 Nanoemulsion-based drug delivery.
Hydrogel and scaffold-based delivery systems
Hydrogels, defined by their 3-dimensional, hydrophilic polymer networks, have been thoroughly investigated for their potential to encapsulate and release bioactive substances in a regulated fashion [187]. Their elevated water content and adjustable characteristics provide them optimal candidates for the delivery of hydrophobic compounds, including terpenes. These improvements highlight the capability of hydrogels to provide prolonged terpene release for diverse biological applications [188]. Zagórska-Dziok et al. [189] have created a hydrogel-based delivery system that provides an innovative method for wound healing by incorporating cannabidiol (CBD) and α-terpineol (TER) into a polymeric matrix. These hydrogels facilitate regulated drug release, exhibit significant antioxidant and anti-inflammatory actions, and possess antibacterial characteristics, therefore establishing an optimum environment for tissue regeneration. By blocking collagenase and elastase activity, they maintain skin integrity and expedite the healing process. This novel technology offers a promising, multimodal approach to enhanced wound treatment [189]. In tissue engineering, bioactive scaffolds are essential for supporting and augmenting the body’s inherent wound healing mechanisms [190]. These scaffolds, typically made from biocompatible materials, offer structural support and can be modified with bioactive chemicals to enhance cellular functions critical for tissue healing [191]. The application of ovine forestomach matrix (OFM) as a biomaterial scaffold has demonstrated potential in soft tissue healing, offering a natural extracellular matrix that facilitates cell infiltration and tissue integration. Recent breakthroughs in the design and application of bioactive scaffolds are facilitating more profitable and effective treatments for wound regeneration [192].
Microneedle and smart drug delivery systems
Microneedles (MNs) have emerged as a revolutionary technique in wound healing, namely for the administration of therapeutic drugs to address biofilm-associated illnesses (Figure 7) [193]. MNs can infiltrate these biofilms and the underlying tissue, enabling targeted and effective administration of medicines directly to the afflicted region. This focused strategy not only improves therapeutic effectiveness but also reduces systemic adverse effects often linked to traditional antibiotic therapies [194]. Integrating stimuli-responsive mechanisms into microneedle designs enhances medication release patterns in accordance with certain wound microenvironments. For example, pH-responsive microneedles have been engineered to utilise the acidic characteristics of chronic wound environments. These MNs are designed with coatings that maintain stability at the physiological skin pH of 4.5 but swiftly disintegrate and release their payload upon exposure to the higher pH values (about 7.5) characteristic of infected wounds. These advanced delivery methods guarantee the exact release of therapeutic substances at the appropriate time and location, hence improving treatment efficacy [195,196].
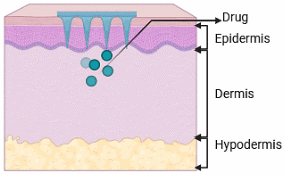
Figure 7 Microneedle drug delivery.
One noteworthy advancement is the application of self-assembling nanoreactors that respond to biofilm microenvironments. To initiate therapeutic responses, these systems take advantage of the hydrogen peroxide-rich and acidic conditions characteristic of biofilm habitats. One example is the use of copper-doped polyoxometalate clusters (Cu-POM) in chemodynamic treatment to transform endogenous hydrogen peroxide into hydroxyl radicals. This mechanism causes bacterial mortality by metabolic disruption, effectively dismantling biofilms. Furthermore, these nanoreactors can reactivate macrophage immune responses, improving the elimination of planktonic bacteria that may evade dissolving biofilms [197]. The use of microneedles for terpene administration in wound healing remains under investigation, however the potential of these systems is considerable. Terpenes, recognised for their antibacterial and anti-inflammatory properties, may be efficiently administered using microneedle platforms to address biofilm-associated illnesses. Further study is required to comprehensively clarify the efficacy and safety of terpene-loaded microneedles in clinical environments [198].
Preclinical and clinical insights into terpene-based biofilm therapy
In a study conducted by Dalleau et al., researchers evaluated the antibiofilm effectiveness of ten terpenes against 3 species of Candida: Candida albicans, Candida parapsilosis, and Candida glabrata. Their research indicated that the application of carvacrol, geraniol, or thymol at a concentration of 0.06% to 24-hour-old C. albicans biofilms led to more than 80% suppression of biofilm development. Carvacrol at 0.03% consistently attained a minimum of 75% inhibition, regardless of the biofilm age. Furthermore, carvacrol at a concentration of 0.125% demonstrated more than 75% suppression of biofilms formed by C. parapsilosis and C. glabrata. The results emphasise the strong antibiofilm efficacy of certain terpenes, notably carvacrol, geraniol, and thymol, against Candida species, highlighting their potential as antifungal agents [199]. This study investigated the antibiofilm mechanisms of phenolic and aldehydic terpenes against Cryptococcus neoformans. Their findings demonstrated that these terpenes not only successfully reduced biofilm development but also destroyed established C. neoformans biofilms. The research offered significant insights into the molecular basis of their antibiofilm action, revealing that these terpenes undermine the structural integrity and survival of fungal biofilms [200].
This study assessed the effectiveness of orange terpene (OT) in eradicating Escherichia coli biofilm cells on beef and several food contact surfaces, including stainless steel (SS), silicone rubber (SR), polyethylene terephthalate (PET), and low-density polyethylene (LDPE). The results indicated that OT had substantial antibiofilm efficacy on all examined surfaces at 37 °C, significantly diminishing E. coli biofilm cells. The findings indicate that OT may serve as a viable natural disinfectant for managing E. coli biofilms in food processing settings, presenting a possible alternative to traditional sanitisation techniques [201]. Rresearch examined the effects of specific terpenes: (−)-trans-Caryophyllene, (S)-cis-Verbenol, (S)-(−)-Limonene, (R)-(+)-Limonene, and Linalool, along with their combinations, on biofilm formation in 2 clinical isolates of Staphylococcus aureus (ST30-t019 and ST5-t311). The results indicated that all terpene combinations suppressed biofilm formation by more than 50% without hindering bacterial growth. The most effective combination (−)-trans-Caryophyllene and Linalool, both at a dosage of 500 μg/mL attained an outstanding 88% inhibition. This formulation decreased the expression of genes related to initial cell adhesion and quorum sensing (sdrD, spa, agr, and hld), while increasing the expression of genes linked to capsular polysaccharide synthesis (cap5B and cap5C). The results underscore the potential of particular terpene combinations as effective anti-biofilm drugs aimed at the first phases of S. aureus biofilm formation [202].
Optimizing terpene-based formulations for enhanced antimicrobial efficacy
Improving the antimicrobial effectiveness of terpene-based formulations necessitates strategic optimisation methods that enhance their stability, bioavailability, and penetration into target areas. Cyclodextrin complexation is one of the more efficacious methods, involving the encapsulation of terpenes within cyclodextrin molecules. This incorporation enhances water solubility, diminishes volatility, and alleviates possible cytotoxicity, hence improving the therapeutic profile while preserving prolonged antibacterial efficacy. Cyclodextrin complexes safeguard terpenes against environmental degradation, so ensuring sustained effectiveness and regulated release, rendering them suitable for medicinal and cosmetic uses [203]. Additionally, terpenes serve as natural penetration enhancers, rendering them advantageous for transdermal and topical antibacterial uses [204]. By compromising the lipid architecture of the stratum corneum, they enhance the deeper and more effective penetration of active medicinal compounds. This characteristic is especially beneficial for addressing localised infections, since improved skin absorption can boost treatment results [205]. Terpenes capacity to regulate membrane fluidity positions them as possible enhancers of medication transport across bacterial cell membranes, hence augmenting their antimicrobial efficacy [206]. The antibacterial effectiveness of terpenes is mainly due to their capacity to damage microbial cell membranes. Moreover, several terpenes have been identified as inhibitors of bacterial efflux pumps, hence augmenting the efficacy of traditional antibiotics against resistant species [138]. Terpenes enhance membrane permeability by integrating into lipid bilayers, resulting in the leaking of cellular contents and eventual cell death [207].
Synergistic effects of terpenes and photodynamic therapy for antimicrobial treatment
Szymczak et al. [208] performed a comprehensive study on the antibacterial efficacy of zinc (II) phthalocyanine regioisomers conjugated with menthol, a monoterpene alcohol. Their research indicated that the incorporation of menthol markedly improved the phototoxic reaction of the photosensitiser against Staphylococcus aureus. This enhancement is ascribed to menthol’s capacity to compromise bacterial membrane integrity, therefore promoting greater diffusion of the photosensitiser into bacterial cells. Upon internalisation, the photosensitiser produces ROS when activated by light, resulting in oxidative stress-induced cellular damage. This work highlights the significance of terpenes in enhancing PDT results by facilitating improved interactions between photosensitisers and bacteria, hence augmenting the efficacy of bacterial elimination.
Kossakowska-Zwierucho et al. [209] investigated the function of farnesol, a sesquiterpene alcohol, in augmenting the effectiveness of photodynamic inactivation (PDI) against Staphylococcus aureus. Their research demonstrates that farnesol markedly enhances the bactericidal efficacy of TMPyP, a cationic porphyrin photosensitiser triggered by red light. The research demonstrated that farnesol pre-treatment enhanced bacterial cell vulnerability to photodynamic injury by compromising lipid bilayers, therefore increasing membrane permeability to the photosensitiser. The augmented permeability enabled more infiltration of the photosensitiser into bacterial structures, resulting in heightened ROS production and consequent oxidative damage following light exposure. The findings strongly indicate that farnesol-induced membrane disruption is a crucial element in enhancing the efficacy of PDT-based antimicrobial approaches.
Li and Wu [210] examined the synergistic effects of 5-aminolevulinic acid (ALA), carvacrol, and blue light on the eradication of polymicrobial biofilms. Their research highlighted the efficacy of carvacrol, a monoterpenoid phenol derived from oregano and thyme, in improving PDT results. Carvacrol was seen to compromise the structural integrity of biofilms by disrupting the EPS matrix, thereby enhancing the penetration of ALA, a precursor to the endogenous photosensitiser protoporphyrin IX (PpIX). When exposed to blue light, PpIX experiences photochemical excitation, producing cytotoxic ROS that efficiently eliminate bacterial cells within biofilms. The research indicates that carvacrol’s antibacterial efficacy, when paired with photodynamic therapy (PDT), can markedly enhance biofilm disintegration, presenting a viable strategy for addressing persistent biofilm-related illnesses.
Lu et al. [211] elucidated the antibacterial activities of carvacrol in combination with photodynamic therapy (PDT). Their research concentrated on carvacrol’s capacity to augment the photodynamic response when exposed to blue light irradiation. The study revealed that carvacrol has intrinsic antibacterial properties and functions as a potentiator of photodynamic therapy by enhancing ROS production in bacterial cells. This dual mechanism leads to heightened oxidative stress, lipid peroxidation, and cellular injury, finally resulting in bacterial cell death. The results highlight the efficacy of integrating carvacrol into PDT methods to more efficiently target multidrug-resistant bacteria [211-212].
Future perspectives and challenges
Personalized and precision medicine approaches
The human microbiome is crucial in health and illness, affecting medication metabolism and treatment results. Recent breakthroughs in microbiome research have shown the possibility of altering microbial assemblages to improve therapeutic success. A study examines the impact of gut microbiota on the progression of infectious illnesses and cancer, highlighting the swift advancement of microbiome-based therapeutic interventions designed to enhance host health. This highlights the necessity of evaluating individual microbiome profiles in the formulation of terpene-based medicines, since the microbiome’s makeup can profoundly influence the therapeutic efficacy of these chemicals [213].
Artificial intelligence (AI) and machine learning (ML) are being used in healthcare to improve predictive analytics and therapeutic approaches. In the domain of biofilm-associated infections, AI has demonstrated potential in forecasting biofilm development and enhancing treatment strategies. Research underscores the use of AI in antimicrobial stewardship, revealing its capacity to enhance decision-making and patient outcomes through the analysis of intricate information to forecast antibiotic resistance trends. Utilising AI and ML, doctors may formulate more efficacious, individualised strategies to prevent and manage biofilm-associated illnesses [214,215].
Integration with regenerative medicine
The integration of stem cell treatment with bioactive chemicals such as terpenes presents considerable potential in regenerative medicine. Research has concentrated on creating biomimetic scaffolds that enhance stem cell survival and differentiation. For instance, a paper describes the production of nanofibrous scaffolds made of polycaprolactone-collagen that have been loaded with terpene-rich Arnebia euchroma extract. These scaffolds’ potential for tissue engineering applications was suggested by their increased stem cell proliferation and preservation of stemness. Thus, by offering both structural support and bioactive stimulation, terpene-based bioactives may improve regeneration outcomes when included into scaffolds loaded with stem cells [216].
By encouraging tissue remodelling, cell migration, and proliferation, growth factors play a crucial role in coordinating the wound healing process. Because terpenes have anti-inflammatory and antibacterial qualities, their use in wound healing techniques has been investigated. A review highlights the potential of several growth factors to improve tissue repair while discussing their involvement in wound healing. Although there aren’t many research directly combining terpenes and growth factors, the advantages of terpenes on their own in lowering inflammation and fighting microbial infections imply that their incorporation into growth factor-mediated treatments may improve wound healing results [217].
Overcoming biofilm resistance while enhancing patient compliance in terpene-based therapies
The durability of biofilms is mostly attributed to their thick structure, which obstructs the entry of therapeutic agents, together with a microenvironment capable of neutralising the effectiveness of these substances. The acidic and hydrogen peroxide-laden environment within biofilms might reduce the efficacy of certain antimicrobials, enabling bacterial survival despite therapeutic interventions [218] Innovative techniques are being studied to reduce bacterial resistance development in biofilms. A potential strategy is the use of biofilm microenvironment-responsive nanoreactors capable of inducing bacterial cell death via processes akin to cuproptosis pathways. These nanoreactors not only compromise the biofilm architecture but also augment the immune response, resulting in more effective elimination of biofilm-associated illnesses [219]. In the context of patient compliance and acceptability, especially in chronic wound treatment, the sensory attributes and possible discomfort associated with terpene formulations are essential factors to consider. Extracellular vesicles (EVs) generated from plants, namely those extracted from Aloe saponaria, exhibit anti-inflammatory and pro-angiogenic characteristics while maintaining little cytotoxicity. It has been demonstrated that these EVs induce angiogenesis, improve cell migration, and encourage fibroblast proliferation all of which are critical for efficient wound healing. Crucially, their biocompatibility and natural origin may lower the likelihood of irritation and sensory pain, increasing patient compliance with chronic wound care [220].
Conclusions
In the fight against infections linked to biofilms in wound healing, terpene-based phytochemicals provide a potential new avenue. Their diverse antibacterial, anti-inflammatory, and antioxidant activities break robust biofilm formations and expedite tissue regeneration, fostering an environment conducive to fast wound closure. Terpenes become natural, strong, and biocompatible therapeutic agents by encouraging host tissue regeneration and focussing on important microbial virulence pathways. Utilising modern drug delivery methods and synergistic formulations to maximise their potential might transform wound management practices, leading to safer, more effective, and sustainable therapies. Terpene-based phytochemicals are promising antibiofilm agents for chronic wound management. Their efficacy is enhanced when combined with conventional antibiotics and advanced delivery systems. Future studies should concentrate on improving their clinical applicability and pharmacokinetics in order to fully realise their therapeutic promise in contemporary medicine.
Acknowledgements
The authors wish to thank all the member of School of Pharmacy, Lincoln University College, Malaysia.
Declaration of generative AI in scientific writing
The authors declare that no generative AI tools were employed in drafting, editing or assisting with any part of the manuscript. The entire work is the result of the authors’ effort and expertise.
CRediT author statement
Shudipta Choudhury and Kamaleswaran Ramachandran: Conceptualized the study and developed the framework for the review. Thanusha Perera: Conducted the literature search and drafted the initial manuscript. Satheesh Babu Natarajan and Anandarajagopal Kalusalingam: Supervised the work and provided feedback throughout the writing process. All authors contributed to manuscript revision, read and approved the final version.
References
M Bartold and S Ivanovski. Biological processes and factors involved in soft and hard tissue healing. Periodontology 2000 2025; 97(1), 16-42.
A Ahmad and SA Zainal. Photobiomodulation therapy for wound healing: A narrative review. IIUM Medical Journal Malaysia 2025; 24(1), 31-39.
S Enoch and DJ Leaper. Basic science of wound healing. Surgery 2005; 23(2), 37-42.
L Cañedo-Dorantes and M Cañedo-Ayala. Skin acute wound healing: a comprehensive review. International Journal of Inflammation 2019; 2019(1), 3706315.
L Häkkinen, H Larjava and L Koivisto. Granulation tissue formation and remodeling. Endodontic Topics 2011; 24(1), 94-129.
J M Reinke and H Sorg. Wound repair and regeneration. European Surgical Research 2012; 49(1), 35-43.
L Zhang, C Weng, Z Zhao and X Fu. Extracorporeal shock wave therapy for chronic wounds: A systematic review and meta-analysis of randomized controlled trials. Wound Repair and Regeneration 2017; 25(4), 697-706.
G Han and R Ceilley. Chronic wound healing: A review of current management and treatments. Advances in Therapy 2017; 34, 599-610.
H Strang, A Kaul, U Parikh, L Masri, S Saravanan, H Li, Q Miao and S Balaji. Role of cytokines and chemokines in wound healing. In: D Bagchi, A Das and S Roy (Eds.). Wound healing, tissue repair, and regeneration in diabetes. Academic Press, Massachusetts, United States, 2020, p. 197-235.
SM McCarty and SL Percival. Proteases and delayed wound healing. Advances in Wound Care 2013; 2(8), 438-447.
M Cano Sanchez, S Lancel, E Boulanger and R Neviere. Targeting oxidative stress and mitochondrial dysfunction in the treatment of impaired wound healing: A systematic review. Antioxidants 2018; 7(8), 98.
A Almatroudi. Biofilm resilience: Molecular mechanisms driving antibiotic resistance in clinical contexts. Biology 2025; 14(2), 165.
H Ragupathi, MM Pushparaj, SM Gopi, DK Govindarajan and K Kandaswamy. Biofilm matrix: A multifaceted layer of biomolecules and a defensive barrier against antimicrobials. Archives of Microbiology 2024; 206(11), 432.
T Bjarnsholt, K Buhlin, YF Dufrêne, M Gomelsky, A Moroni, M Ramstedt, KP Rumbaugh, T Schulte, L Sun, B Åkerlund and U Römling. Biofilm formation - what we can learn from recent developments. Journal of Internal Medicine 2018; 284(4), 332-345.
N Khardori and M Yassien. Biofilms in device-related infections. Journal of Industrial Microbiology and Biotechnology 1995; 15(3), 141-147.
MM Zafer, GA Mohamed, SR Ibrahim, S Ghosh, C Bornman and MA Elfaky. Biofilm-mediated infections by multidrug-resistant microbes: a comprehensive exploration and forward perspectives. Archives of Microbiology 2024; 206(3), 101.
NK Ragupathi, B Veeraraghavan, E Karunakaran and PN Monk. Biofilm-mediated nosocomial infections and its association with antimicrobial resistance: Detection, prevention, and management. Frontiers in Medicine 2022; 9, 987011.
RD Scott. The direct medical costs of healthcare-associated infections in US hospitals and the benefits of prevention, Available at: https://stacks.cdc.gov/view/cdc/11550/cdc_11550_DS1.pdf, accessed October 2025.
V Jean-Pierre, A Boudet, P Sorlin, Q Menetrey, R Chiron, JP Lavigne and H Marchandin. Biofilm formation by Staphylococcus aureus in the specific context of cystic fibrosis. International Journal of Molecular Sciences 2022; 24(1), 597.
C Moser, PØ Jensen, K Thomsen, M Kolpen, M Rybtke, AS Lauland, H Trøstrup and T Tolker-Nielsen. Immune responses to Pseudomonas aeruginosa biofilm infections. Frontiers in Immunology 2021; 12, 625597.
AS Pugazhendhi, F Wei, M Hughes and M Coathup. Bacterial adhesion, virulence, and biofilm formation. In: M Coathup (Ed.). Musculoskeletal infection. Springer International Publishing, Cham, Switzerland, 2022, p. 19-64.
I Ahmad, A Nadeem, F Mushtaq, N Zlatkov, M Shahzad, AV Zavialov, SN Wai and BE Uhlin. Csu pili dependent biofilm formation and virulence of Acinetobacter baumannii. npj Biofilms and Microbiomes 2023; 9(1), 101.
T Das, S Sehar and M Manefield. The roles of extracellular DNA in the structural integrity of extracellular polymeric substance and bacterial biofilm development. Environmental Microbiology Reports 2013; 5(6), 778-786.
CB Whitchurch, T Tolker-Nielsen, PC Ragas and JS Mattick. Extracellular DNA required for bacterial biofilm formation. Science 2002; 295, 1487.
PS Stewart, B White, L Boegli, T Hamerly, KS Williamson, MJ Franklin, B Bothner, GA James, S Fisher, FG Vital-Lopez and A Wallqvist. Conceptual model of biofilm antibiotic tolerance that integrates phenomena of diffusion, metabolism, gene expression, and physiology. Journal of Bacteriology 2019; 201(22), e00128-19.
K Azeem, S Fatima, A Ali, A Ubaid, FM Husain and M Abid. Biochemistry of bacterial biofilm: Insights into antibiotic resistance mechanisms and therapeutic intervention. Life 2025; 15(1), 49.
A Prinzi and R Rohde. The role of bacterial biofilms in antimicrobial resistance. American Society for Microbiology 2023; Available from: https://asm.org/articles/2023/march/the-role-of-bacterial-biofilms-in-antimicrobial-re
A Jain and DK Parihar. Antibacterial, biofilm dispersal and antibiofilm potential of alkaloids and flavonoids of Curcuma. Biocatalysis and Agricultural Biotechnology 2018; 16, 677-682.
X Song, YX Xia, ZD He and HJ Zhang. A review of natural products with anti-biofilm activity. Current Organic Chemistry 2018; 22(8), 789-817.
NA Mahizan, SK Yang, CL Moo, AAM Song, CM Chong, CW Chong, A Abushelaibi, SH Lim and KS Lai. Terpene derivatives as potential agents against antimicrobial resistance (AMR) pathogens. Molecules 2019; 24(14), 2631.
C Shi, M Che, X Zhang, Z Liu, R Meng, X Bu, H Ye and N Guo. Antibacterial activity and mode of action of totarol against Staphylococcus aureus in carrot juice. Journal of Food Science and Technology 2018; 55, 924-934.
R Jaiswal, TK Beuria, R Mohan, SK Mahajan and D Panda. Totarol inhibits bacterial cytokinesis by perturbing the assembly dynamics of FtsZ. Biochemistry 2007; 46(14), 4211-4220.
M Majid, H Naveed, SA Mohsin, MT Khan, W Hussain, Y Ruan, H Lu, H Fatima and X Hu. Nature’s answer to superbugs: Berberis lycium and its potent isoquinoline alkaloid, Berberine-A, can target bacterial enzymome to combat antibiotic resistance. Journal of Antibiotic Research 2024. http://dx.doi.org/10.2139/ssrn.4984042
Z Javanmard, M Pourhajibagher and A Bahador. Advancing anti-biofilm strategies: innovations to combat biofilm-related challenges and enhance efficacy. Journal of Basic Microbiology 2024; 64(12), e2400271.
A García, B González, C Harvey, I Izquierdo-Barba and M Vallet-Regí. Effective reduction of biofilm through photothermal therapy by gold core@shell based mesoporous silica nanoparticles. Microporous and Mesoporous Materials 2021; 328, 111489.
E Álvarez, B González, D Lozano, AL Doadrio, M Colilla and I Izquierdo-Barba. Nanoantibiotics based in mesoporous silica nanoparticles: new formulations for bacterial infection treatment. Pharmaceutics 2021; 13(12), 2033.
K Summer, Q Guo, L Liu, B Barkla, S Giles and K Benkendorff. Antimicrobial proteins from oyster hemolymph improve the efficacy of conventional antibiotics. PLoS One 2025; 20(1), e0312305.
O Alliod, JP Valour, S Urbaniak, H Fessi, D Dupin and C Charcosset. Preparation of oil-in-water nanoemulsions at large scale using premix membrane emulsification and Shirasu Porous Glass (SPG) membranes. Colloids and Surfaces A 2018; 557, 76-84.
S Noori, F Zeynali and H Almasi. Antimicrobial and antioxidant efficiency of nanoemulsion-based edible coating containing ginger (Zingiber officinale) essential oil and its effect on safety and quality attributes of chicken breast fillets. Food Control 2018; 84, 312-320.
H Cheng, Q Zhu and J Wan. Impact of cinnamaldehyde nanoemulsion on Listeria monocytogenes biofilms on broth-treated food contact surfaces. LWT 2025; 117500.
S Yao, L Hao, R Zhou, Y Jin, J Huang and C Wu. Multispecies biofilms in fermentation: biofilm formation, microbial interactions, and communication. Comprehensive Reviews in Food Science and Food Safety 2022; 21(4), 3346-3375.
CR Armbruster and MR Parsek. New insight into the early stages of biofilm formation. Proceedings of the National Academy of Sciences 2018; 115(17), 4317-4319.
T Tolker-Nielsen. Biofilm development. In: M Ghannoum, M Parsek, M Whiteley and PK Mukherjee (Eds.). Microbial biofilms. Wiley, New Jersey, United States, 2015, p. 51-66.
MS Khan, MM Altaf and I Ahmad. Chemical nature of biofilm matrix and its significance. In: I Ahmad and FM Husain. Biofilms in plant and soil health. Wiley, New Jersey, United States, 2017, p. 151-177.
L Karygianni, Z Ren, H Koo and T Thurnheer. Biofilm matrixome: Extracellular components in structured microbial communities. Trends in Microbiology 2020; 28(8), 668-681.
E Balducci, F Papi, DE Capialbi and LD Bino. Polysaccharides’ structures and functions in biofilm architecture of antimicrobial-resistant (AMR) pathogens. International Journal of Molecular Sciences 2023; 24(4), 4030.
J S Ramírez-Larrota and U Eckhard. An introduction to bacterial biofilms and their proteases, and their roles in host infection and immune evasion. Biomolecules 2022; 12(2), 306.
D Campoccia, L Montanaro and CR Arciola. Extracellular DNA (eDNA): A major ubiquitous element of the bacterial biofilm architecture. International Journal of Molecular Sciences 2021; 22(16), 9100.
DR Dixon and RP Darveau. Lipopolysaccharide heterogeneity: Innate host responses to bacterial modification of lipid A structure. Journal of Dental Research 2005; 84(7), 584-595.
J Jo, A Price-Whelan and LE Dietrich. Gradients and consequences of heterogeneity in biofilms. Nature Reviews Microbiology 2022; 20(10), 593-607.
SN Abraham, N Sharon and I Ofek. Adhesion and colonisation. Molecular Medical Microbiology 2002; 1, 629-644.
KY Le and M Otto. Quorum-sensing regulation in staphylococci-an overview. Frontiers in Microbiology 2015; 6, 1174.
TJ Polaske, KH West, K Zhao, DL Widner, JT York and HE Blackwell. Chemical and biomolecular insights into the Staphylococcus aureus agr quorum sensing system: current progress and ongoing challenges. Israel Journal of Chemistry 2023; 63(5-6), e202200096.
T Abee, Á T Kovács, OP Kuipers and S Van der Veen. Biofilm formation and dispersal in Gram-positive bacteria. Current Opinion in Biotechnology 2011; 22(2), 172-179.
L Zhou, Y Zhang, Y Ge, X Zhu and J Pan. Regulatory mechanisms and promising applications of quorum sensing-inhibiting agents in control of bacterial biofilm formation. Frontiers in Microbiology 2020; 11, 589640.
MJ Franklin, DE Nivens, JT Weadge and PL Howell. Biosynthesis of the Pseudomonas aeruginosa extracellular polysaccharides, alginate, Pel, and Psl. Frontiers in Microbiology 2011; 2, 167.
A Hartmann. Quorum sensing N-acyl-homoserine lactone signal molecules of plant beneficial Gram-negative rhizobacteria support plant growth and resistance to pathogens. Rhizosphere 2020; 16, 100258.
Z Wang, X Hou, G Shang, G Deng, K Luo and M Peng. Exploring fatty acid β-oxidation pathways in bacteria: from general mechanisms to DSF signaling and pathogenicity in Xanthomonas. Current Microbiology 2024; 81(10), 336.
YW He, Y Deng, Y Miao, S Chatterjee, TM Tran, J Tian and S Lindow. DSF-family quorum sensing signal-mediated intraspecies, interspecies, and inter-kingdom communication. Trends in Microbiology 2023; 31(1), 36-50.
M Gallique, M Bouteiller and A Merieau. The type VI secretion system: A dynamic system for bacterial communication? Frontiers in Microbiology 2017; 8, 1454.
A Vetrivel, M Ramasamy, P Vetrivel, S Natchimuthu, S Arunachalam, GS Kim and R Murugesan. Pseudomonas aeruginosa biofilm formation and its control. Biologics 2021; 1(3), 312-336.
Z Pang, R Raudonis, BR Glick, TJ Lin and Z Cheng. Antibiotic resistance in Pseudomonas aeruginosa: mechanisms and alternative therapeutic strategies. Biotechnology Advances 2019; 37(1), 177-192.
HC Neu. The role of Pseudomonas aeruginosa in infections. Journal of Antimicrobial Chemotherapy 1983; 11(SB), 1-3.
A Nasser, MM Dallal, S Jahanbakhshi, T Azimi and L Nikouei. Staphylococcus aureus: Biofilm formation and strategies against it. Current Pharmaceutical Biotechnology 2022; 23(5), 664-678.
A Pantosti, A Sanchini and M Monaco. Mechanisms of antibiotic resistance in Staphylococcus aureus. Future Microbiology 2007; 2(3), 323-334.
MZ David and RS Daum. Treatment of Staphylococcus aureus infections. In: MZ David and RS Daum (Eds.). Staphylococcus aureus: Microbiology, pathology, immunology, therapy and prophylaxis. Springer, Cham, Swizerland, 2017, p. 325-383.
G Laverty, SP Gorman and BF Gilmore. Biomolecular mechanisms of Pseudomonas aeruginosa and Escherichia coli biofilm formation. Pathogens 2014; 3(3), 596-632.
A Ito, A Taniuchi, T May, K Kawata and S Okabe. Increased antibiotic resistance of Escherichia coli in mature biofilms. Applied and Environmental Microbiology 2009; 75(12), 4093-4100.
TA Russo and JR Johnson. Medical and economic impact of extraintestinal infections due to Escherichia coli: focus on an increasingly important endemic problem. Microbes and Infection 2003; 5(5), 449-456.
D Senadheera and DG Cvitkovitch. Quorum sensing and biofilm formation by Streptococcus mutans. In: Bacterial signal transduction: Networks and drug targets. Springer, New York, 2008, p. 178-188.
J Nan, E Brostromer, XY Liu, O Kristensen and XD Su. Bioinformatics and structural characterization of a hypothetical protein from Streptococcus mutans: implication of antibiotic resistance. PLoS One 2009; 4(10), e7245.
SM Daboor, FS Masood, MS Al-Azab and EE Nori. A review on Streptococcus mutans with its diseases dental caries, dental plaque and endocarditis. Indian Journal of Microbiology Research 2015; 2(2), 76-82.
T Atriwal, K Azeem, FM Husain, A Hussain, MN Khan, MF Alajmi and M Abid. Mechanistic understanding of Candida albicans biofilm formation and approaches for its inhibition. Frontiers in Microbiology 2021; 12, 638609.
LE Cowen, JB Anderson and LM Kohn. Evolution of drug resistance in Candida albicans. Annual Review of Microbiology 2002; 56(1), 139-165.
CJ Nobile and AD Johnson. Candida albicans biofilms and human disease. Annual Review of Microbiology 2015; 69(1), 71-92.
JA Gaddy and LA Actis. Regulation of Acinetobacter baumannii biofilm formation. Future Microbiology 2009; 4(3), 273-278.
I Kyriakidis, E Vasileiou, ZD Pana and A Tragiannidis. Acinetobacter baumannii antibiotic resistance mechanisms. Pathogens 2021; 10(3), 373.
A Gedefie, W Demsis, M Ashagrie, Y Kassa, M Tesfaye, M Tilahun, H Bisetegn and Z Sahle. Acinetobacter baumannii biofilm formation and its role in disease pathogenesis: A review. Infection and Drug Resistance 2021; 10, 3711-3719.
C Vuotto, F Longo, MP Balice, G Donelli and PE Varaldo. Antibiotic resistance related to biofilm formation in Klebsiella pneumoniae. Pathogens 2014; 3(3), 743-758.
ME Guerra, G Destro, B Vieira, AS Lima, LF Ferraz, AP Hakansson, M Darrieux and TR Converso. Klebsiella pneumoniae biofilms and their role in disease pathogenesis. Frontiers in Cellular and Infection Microbiology 2022; 12, 877995.
A Toledo-Arana, J Valle, C Solano, MJ Arrizubieta, C Cucarella, M Lamata, B Amorena, J Leiva, JR Penadés and I Lasa. The enterococcal surface protein, Esp, is involved in Enterococcus faecalis biofilm formation. Applied and Environmental Microbiology 2001; 67(10), 4538-4545.
IU Rathnayake, M Hargreaves and F Huygens. Antibiotic resistance and virulence traits in clinical and environmental Enterococcus faecalis and Enterococcus faecium isolates. Systematic and Applied Microbiology 2012; 35(5), 326-333.
SB Bhardwaj. Enterococci: An important nosocomial pathogen. Pathogenic Bacteria 2019; 12, 1-15.
P Chakraborty, S Bajeli, D Kaushal, BD Radotra and A Kumar. Biofilm formation in the lung contributes to virulence and drug tolerance of Mycobacterium tuberculosis. Nature Communications 2021; 12(1), 1606.
CA Peloquin and SE Berning. Infection caused by Mycobacterium tuberculosis. Annals of Pharmacotherapy 1994; 28(1), 72-84.
H Yonezawa, T Osaki and S Kamiya. Biofilm formation by Helicobacter pylori and its involvement in antibiotic resistance. BioMed Research International 2015; 2015(1), 914791.
P Sipponen and H Hyvärinen. Role of Helicobacter pylori in the pathogenesis of gastritis, peptic ulcer and gastric cancer. Scandinavian Journal of Gastroenterology 1993; 28(S196), 3-6.
SM Jacobsen and ME Shirtliff. Proteus mirabilis biofilms and catheter-associated urinary tract infections. Virulence 2011; 2(5), 460-465.
E Alqurashi, K Elbanna, I Ahmad and HH Abulreesh. Antibiotic resistance in Proteus mirabilis: mechanism, status, and public health significance. Journal of Pure and Applied Microbiology 2022; 16(3), 1191-1203.
JM Kopaczyk, J Warguła and T Jelonek. The variability of terpenes in conifers under developmental and environmental stimuli. Environmental and Experimental Botany 2020; 180, 104197.
FD Mabou and IB Yossa. Terpenes: Structural classification and biological activities. IOSR Journal of Pharmacy and Biological Sciences 2021; 16, 25-40.
K Wijesekera and AS Dissanayake. Terpenes. In: M Napagoda and L Jayasinghe (Eds.). Chemistry of natural products: Phytochemistry and pharmacognosy of medicinal plants. De Gruyter, Berlin, Germany, 2022, p. 65-90.
H Loza-Tavera. Monoterpenes in essential oils: biosynthesis and properties. In: F Shahidi, P Kolodziejczyk, JR Whitaker, AL Munguia and G Fuller (Eds.). Chemicals via higher plant bioengineering. Advances in experimental medicine and biology. Springer, Boston, United States, 1999, p. 49-62.
F de Faveri Favero, R Grando, FR Nonato, IM Sousa, NC Queiroz, GB Longato, RR Zafred, JE Carvalho, HM Spindola and MA Foglio. Artemisia annua L.: evidence of sesquiterpene lactones’ fraction antinociceptive activity. BMC Complementary and Alternative Medicine 2014; 14(1), 1-10.
ANM Alamgir. Secondary metabolites: Secondary metabolic products consisting of C and H; C, H, and O; N, S, and P elements; and O/N heterocycles. In: ANM Alamgir (Ed.). Therapeutic use of medicinal plants and their extracts, volume 2: Phytochemistry and bioactive compounds. Springer, Cham, Switzerland, 2018, p. 165-309.
JM DeLong and RK Prange. Taxus spp.: Botany, horticulture, and source of anti-cancer compounds. Horticultural Reviews 2006; 32, 299-327.
S Banerjee, S Banerjee, A Bishayee, MN Da Silva, OA Sukocheva, E Tse, N Casarcia and A Bishayee. Cellular and molecular mechanisms underlying the potential of betulinic acid in cancer prevention and treatment. Phytomedicine 2024; 132, 155858.
E Gambino, A Maione, M Guida, L Albarano, F Carraturo, E Galdiero and V Di Onofrio. Evaluation of the pathogenic mixed biofilm formation of Pseudomonas aeruginosa/Staphylococcus aureus and treatment with limonene on 3 different materials by a dynamic model. International Journal of Environmental Research and Public Health 2022; 19(6), 3741.
V Poscente, L Di Gregorio, M Costanzo, C Nobili, R Bernini, L Garavaglia and A Bevivino. Lactiplantibacillus plantarum monolayer enhanced bactericidal action of carvacrol: Biofilm inhibition of viable foodborne pathogens and spoilage microorganisms. Frontiers in Microbiology 2023; 14, 1296608.
S Asadi, B Nayeri-Fasaei, T Zahraei-Salehi, R Yahya-Rayat, N Shams and A Sharifi. Antibacterial and anti-biofilm properties of carvacrol alone and in combination with cefixime against Escherichia coli. BMC Microbiology 2023; 23(1), 55.
SY Wei, YL Li, L Wang, ZY Chu, YC Qin and H Zeng. α-Pinene: inhibitor of Acinetobacter baumannii biofilms and potential therapeutic agent for pneumonia. International Immunopharmacology 2025; 151, 114287.
P Jurado, C Uruén, S Martínez, E Lain, S Sánchez, A Rezusta and J Arenas. Essential oils of Pinus sylvestris, Citrus limon and Origanum vulgare exhibit high bactericidal and anti-biofilm activities against Neisseria gonorrhoeae and Streptococcus suis. Biomedicine & Pharmacotherapy 2023; 168, 115703.
HJ Yoo and SK Jwa. Inhibitory effects of β-caryophyllene on Streptococcus mutans biofilm. Archives of Oral Biology 2018; 88, 42-46.
K Gu, P Ouyang, Y Hong, Y Dai, T Tang, C He, G Shu, X Liang, H Tang, L Zhu and Z Xu. Geraniol inhibits biofilm formation of methicillin-resistant Staphylococcus aureus and increases the therapeutic effect of vancomycin in vivo. Frontiers in Microbiology 2022; 13, 960728.
A Kannappan, M Sivaranjani, R Srinivasan, J Rathna, SK Pandian and AV Ravi. Inhibitory efficacy of geraniol on biofilm formation and development of adaptive resistance in Staphylococcus epidermidis RP62A. Journal of Medical Microbiology 2017; 66(10), 1506-1515.
P K Sagar, P Sharma and R Singh. Inhibition of quorum sensing-regulated virulence factors and biofilm formation by Eucalyptus globulus against multidrug-resistant Pseudomonas aeruginosa. Journal of Pharmacopuncture 2022; 25(1), 37-45.
K Vijayakumar, V Manigandan, D Jeyapragash, V Bharathidasan, B Anandharaj and M Sathya. Eucalyptol inhibits biofilm formation of Streptococcus pyogenes and its mediated virulence factors. Journal of Medical Microbiology 2020; 69(11), 1308-1318.
T Liu, J Kang and L Liu. Thymol as a critical component of Thymus vulgaris L. essential oil combats Pseudomonas aeruginosa by intercalating DNA and inactivating biofilm. LWT 2021; 136, 110354.
ST Khan, M Khan, J Ahmad, R Wahab, OH Abd-Elkader, J Musarrat, HZ Alkhathlan and AA Al-Kedhairy. Thymol and carvacrol induce autolysis, stress, growth inhibition and reduce the biofilm formation by Streptococcus mutans. AMB Express 2017; 7(1), 1-10.
P Gupta, M Sharma, N Arora, V Pruthi and KM Poluri. Chemistry and biology of farnesol and its derivatives: quorum sensing molecules with immense therapeutic potential. Current Topics in Medicinal Chemistry 2018; 18(22), 1937-1954.
B Gaálová-Radochová, S Kendra, L Jordao, L Kursawe, J Kikhney, A Moter and H Bujdáková. Effect of quorum sensing molecule farnesol on mixed biofilms of Candida albicans and Staphylococcus aureus. Antibiotics 2023; 12(3), 441.
L Wang, J Wang, K Zhang, J Zhang, D Cui, J Wang, P Ji, Y Wei and J Li. Linalool as a potential agent for inhibiting Escherichia coli biofilm formation and exopolysaccharide production. BMC Veterinary Research 2025; 21(1), 235.
G Shen, L Yang, X Lv, Y Zhang, X Hou, M Li, M Zhou, L Pan, A Chen and Z Zhang. Antibiofilm activity and mechanism of linalool against food spoilage Bacillus amyloliquefaciens. International Journal of Molecular Sciences 2023; 24(13), 10980.
PM Maquera-Huacho, CC Tonon, MF Correia, RS Francisconi, EA Bordini, É Marcantonio and DM Spolidorio. In vitro antibacterial and cytotoxic activities of carvacrol and terpinen-4-ol against biofilm formation on titanium implant surfaces. Biofouling 2018; 34(6), 699-709.
L Cordeiro, P Figueiredo, H Souza, A Sousa, F Andrade-Júnior, D Medeiros, J Nóbrega, D Silva, E Martins, J Barbosa-Filho and E Lima. Terpinen-4-ol as an antibacterial and antibiofilm agent against Staphylococcus aureus. International Journal of Molecular Sciences 2020; 21(12), 4531.
D Cox-Georgian, N Ramadoss, C Dona and C Basu. Therapeutic and medicinal uses of terpenes. In: N Joshee, S Dhekney and P Parajuli (Eds.). Medicinal plants: From farm to pharmacy. Springer, Cham, Switzerland, 2019, p. 333-359.
D Dufour, V Leung and CM Lévesque. Bacterial biofilm: Structure, function, and antimicrobial resistance. Endodontic Topics 2010; 22(1), 2-16.
TB Farnan, J McCallum, A Awa, AD Khan and SJ Hall. Tea tree oil: In vitro efficacy in otitis externa. The Journal of Laryngology & Otology 2005; 119(3), 198-201.
R Iseppi, M Mariani, S Benvenuti, E Truzzi and P Messi. Effects of Melaleuca alternifolia Chell (Tea Tree) and Eucalyptus globulus Labill. essential oils on antibiotic-resistant bacterial biofilms. Molecules 2023; 28(4), 1671.
C Brinsi, A Abidi, K Hosni, H Tounsi and H Sebai. Protective effect of Eucalyptus globulus extracts against bleomycin-induced pulmonary fibrosis in rats. Journal of Medicinal Food 2022; 25(7), 741–750.
B Saoudi, K Bariz, S Saci, Y Belounis, H Ait Issad, M Abbaci, M A Mustapha, E H Nabti, R Alenazy, M S Alhussaini and A A Alyahya. Enhancing antibiotic efficacy and combating biofilm formation: evaluating the synergistic potential of Origanum vulgare essential oil against multidrug-resistant Gram-negative bacteria. Microorganisms 2024; 12(8), 1651.
C Rossi, C Chaves-López, A Serio, F Anniballi, L Valbonetti and A Paparella. Effect of Origanum vulgare essential oil on biofilm formation and motility capacity of Pseudomonas fluorescens strains isolated from discoloured mozzarella cheese. Journal of Applied Microbiology 2018; 124(5), 1220-1231.
S Kajjari, BK Vanishree, S Janardhanan, VH Patil, C Uppin and SM Hugar. Antimicrobial efficacy of Mangifera indica, Mentha arvensis, and chlorhexidine mouthwashes on Streptococcus mutans and Candida albicans in children: A comparative in vivo study. International Journal of Clinical Pediatric Dentistry 2024; 17(S1), S78.
R Karicheri and B Antony. Antibacterial and antibiofilm activities of peppermint (Mentha piperita Linn) and menthol mint (Mentha arvensis Linn) essential oils on Aggregatibacter actinomycetemcomitans isolated from orodental infections. European Journal of Pharmaceutical and Medical Research 2016; 3, 577-581.
SO Egwuatu, TM Obuotor, OS Taiwo, WE Ike, AE Ojo, PO Isibor, OF Adeniji, FM Oyeyipo, OA Awotoye, P Akinduti and P Egwuatu. Antibacterial efficacy of Thymus vulgaris essential oil against extended-spectrum beta-lactamase-producing Escherichia coli in urinary tract infections. In: Biotechnological Approaches to Sustainable Development Goals. Cham: Springer Nature Switzerland; 2023. p. 159–172.
TW Kang, G Chandrasekaran, EC Hwang, HS Kim and VK Lakshmanan. Characterization and antibacterial activity of PVA-PVP-CS carvacrol-loaded polymer composite films for urinary catheter. International Journal of Polymeric Materials and Polymeric Biomaterials 2018; 67(17), 1016-1027.
R De, P Kundu, S Swarnakar, T Ramamurthy, A Chowdhury, GB Nair and AK Mukhopadhyay. Antimicrobial activity of curcumin against Helicobacter pylori isolates from India and during infections in mice. Antimicrobial Agents and Chemotherapy 2009; 53(4), 1592-1597.
UJ Tsopmene, CRT Kuaté, PN Kayoka-Kabongo, BN Bisso, A Metopa, CT Mofor and JP Dzoyem. Antibiofilm activity of curcumin and piperine and their synergistic effects with antifungals against Candida albicans clinical isolates. Scientifica 2024; 2024(1), 2025557.
A Jerkovic-Mujkic, K Masic and TJ Dzelilovic. Antimicrobial potential of herbal products against Clostridioides difficile. Health 2024; 4, 39-49.
S Justin and B Antony. Antibacterial activity of the essential oils of Syzygium aromaticum (L.) Merr. Perry (Clove), Myristica fragrans Houtt. (Nutmeg) and Zingiber officinale Roscoe (Ginger) against clinical isolates of Clostridium difficile: An in vitro study. International Journal of Contemporary Medical Research 2016; 3, 1085-1089.
R Najafi-Taher, AJ Kohneloo, VE Farsani, NM Rayeni, HR Moghimi, A Ehsani and A Amani. A topical gel of tea tree oil nanoemulsion containing adapalene versus adapalene marketed gel in patients with acne vulgaris: A randomized clinical trial. Archives of Dermatological Research 2022; 314(9), 1-7.
D Lauritano, D Pazzi, A Iapichino, RM Gaudio, MD Muzio, LL Russo and F Pezzetti. Evaluation of the efficacy of a new oral gel containing carvacrol and thymol for home oral care in the management of chronic periodontitis using pcr analysis: A microbiological pilot study. Journal of Biological Regulators & Homeostatic Agents 2016; 30(2S1), 129-134.
VC Patole, SP Chaudhari, AP Pandit and PP Lokhande. Thymol and eugenol loaded chitosan dental film for treatment of periodontitis. Indian Drugs 2019; 56(6), 51-58.
H Hamzah, S U Pratiwi and T Hertiani. Efficacy of thymol and eugenol against polymicrobial biofilm. Indonesian Journal of Pharmacy 2018; 29(4), 214-221.
W Mączka, M Twardawska, M Grabarczyk and K Wińska. Carvacrol - a natural phenolic compound with antimicrobial properties. Antibiotics 2023; 12(5), 824.
F Murina, F Vicariotto and S Di Francesco. Thymol, eugenol and lactobacilli in a medical device for the treatment of bacterial vaginosis and vulvovaginal candidiasis. New Microbiologica 2018; 41(3), 220-224.
C Liu, Y Zhang, S Kong, I Tsui, Y Yu and F Han. Applications and therapeutic actions of complementary and alternative medicine for women with genital infection. Evidence-Based Complementary and Alternative Medicine 2014; 2014(1), 658624.
AC Guimarães, LM Meireles, MF Lemos, MC Guimarães, DC Endringer, M Fronza and R Scherer. Antibacterial activity of terpenes and terpenoids present in essential oils. Molecules 2019; 24(13), 2471.
KJ Dias, GM Miranda, JR Bessa, AC Araújo, PR Freitas, RS Almeida, CL Paulo, J B Neto, HD Coutinho and J Ribeiro-Filho. Terpenes as bacterial efflux pump inhibitors: a systematic review. Frontiers in Pharmacology 2022; 13, 953982.
S Burt. Essential oils: Their antibacterial properties and potential applications in foods - a review. International Journal of Food Microbiology 2004; 94(3), 223-253.
A Masyita, RM Sari, AD Astuti, B Yasir, NR Rumata, TB Emran, F Nainu and J Simal-Gandara. Terpenes and terpenoids as main bioactive compounds of essential oils, their roles in human health and potential application as natural food preservatives. Food Chemistry: X 2022; 13, 100217.
X Jin, M Zhang, J Lu, X Duan, J Chen, Y Liu, W Chang and H Lou. Hinokitiol chelates intracellular iron to retard fungal growth by disturbing mitochondrial respiration. Journal of Advanced Research 2021; 34, 65-77.
M Mikulášová, R Chovanová and Š Vaverková. Synergism between antibiotics and plant extracts or essential oils with efflux pump inhibitory activity in coping with multidrug-resistant staphylococci. Phytochemistry Reviews 2016; 15, 651-662.
W Wang, X Bao, M Bové, P Rigole, X Meng, J Su and T Coenye. Antibiofilm activities of borneol-citral-loaded Pickering emulsions against Pseudomonas aeruginosa and Staphylococcus aureus in physiologically relevant chronic infection models. Microbiology Spectrum 2022; 10(5), e01696-22.
L Gutiérrez, A Escudero, R Batlle and C Nerín. Effect of mixed antimicrobial agents and flavors in active packaging films. Journal of Agricultural and Food Chemistry 2009; 57(18), 8564-8571.
C Gan, E Langa, A Valenzuela, D Ballestero and MR Pino-Otín. Synergistic activity of thymol with commercial antibiotics against critical and high WHO priority pathogenic bacteria. Plants 2023; 12(9), 1868.
B Huerta Lorenzo, Á Galán-Relaño, E Barba-Sánchez, A Romero-Salmoral, AL Solarte Portilla, L Gómez-Gascón and RJ Astorga Márquez. Potentiation of the antimicrobial effect of oxytetracycline combined with cinnamon, clove, oregano, and red thyme essential oils against MDR Salmonella enterica strains. Animals 2024; 14(9), 1347.
H Si, J Hu, Z Liu and ZL Zeng. Antibacterial effect of oregano essential oil alone and in combination with antibiotics against extended-spectrum β-lactamase-producing Escherichia coli. FEMS Immunology & Medical Microbiology 2008; 53(2), 190-194.
F Fratini, C Pecorini, I Resci, E Copelotti, FP Nocera, B Najar and S Mancini. Evaluation of the synergistic antimicrobial activity of essential oils and cecropin A natural peptide on Gram-negative bacteria. Animals 2025; 15(2), 282.
PS Costa, SS Oliveira, EB Souza, EH Brito, CS Cavalcante, SM Morais, AL Leal, HM Barreto, AM Teixeira, CE Nogueira and RO Fontenelle. Antifungal activity and synergistic effect of essential oil from Lippia alba against Trichophyton rubrum and Candida spp. Revista Virtual de Química 2020; 12(6), 0000-0000.
D C Costa, A B Vermelho, C A Almeida, E P Dias, S M Cedrola, M D Arrigoni-Blank, A F Blank, C S Alviano and D S Alviano. Inhibitory effect of linalool-rich essential oil from Lippia alba on the peptidase and keratinase activities of dermatophytes. Journal of Enzyme Inhibition and Medicinal Chemistry 2014; 29(1), 12-17.
J Reichling. Anti-biofilm and virulence factor-reducing activities of essential oils and oil components as a possible option for bacterial infection control. Planta Medica 2020; 86(8), 520-537.
Y Ge, J Tang, H Fu and Y Fu. Terpinen-4-ol liposomes-incorporated chitosan/polyethylene oxide electrospun nanofibrous film ameliorates the external microenvironment of healing cutaneous wounds. Journal of Applied Polymer Science 2021; 138(2), 49670.
JI Deen, AS Zawad, M Uddin, MA Chowdhury, SQ Al Araby and MA Rahman. Terpinen-4-ol, a volatile terpene molecule, extensively electrifies biological systems against oxidative stress-linked pathogenesis. Advances in Redox Research 2023; 9, 100082.
T Kennewell, S Mashtoub, GS Howarth, AJ Cowin and Z Kopecki. Antimicrobial and healing-promoting properties of animal and plant oils for the treatment of infected wounds. Wound Practice & Research: Journal of the Australian Wound Management Association 2019; 27(4), 175-183.
RG Frykberg and J Banks. Challenges in the treatment of chronic wounds. Advances in Wound Care 2015; 4(9), 560-582.
A Scalise, A Bianchi, C Tartaglione, E Bolletta, M Pierangeli, M Torresetti, M Marazzi and GI Di Benedetto. Microenvironment and microbiology of skin wounds: the role of bacterial biofilms and related factors. Seminars in Vascular Surgery 2015; 28(3-4), 151-159.
JS Boateng, KH Matthews, HN Stevens and GM Eccleston. Wound healing dressings and drug delivery systems: A review. Journal of Pharmaceutical Sciences 2008; 97(8), 2892-2923.
G Gainza, S Villullas, JL Pedraz, RM Hernandez and M Igartua. Advances in drug delivery systems to release growth factors for wound healing and skin regeneration. Nanomedicine: Nanotechnology, Biology and Medicine 2015; 11(6), 1551-1573.
A Sultana, M Zare, V Thomas, TS Kumar and S Ramakrishna. Nano-based drug delivery systems: Conventional routes, recent developments, and future prospects. Medicine in Drug Discovery 2022; 15, 100134.
H Bhardwaj, S Khute, R Sahu and RK Jangde. Advanced drug delivery systems for management of chronic diabetic wound healing. Current Drug Targets 2023; 24(16), 1239-1259.
L Pachuau. Recent developments in novel drug delivery systems for wound healing. Expert Opinion on Drug Delivery 2015; 12(12), 1895-1909.
J Jose, KB Khot, SP Thomas, H Chopra, G Gopan, A Bandiwadekar, A Babu, SR Ugare, GM Ashraf, A Dhyani and V Vora. Advances in microneedles-based drug delivery systems on promoting wound healing. Journal of Drug Delivery Science and Technology 2023; 90, 105163.
Y Li, J Wang, Y Wang and W Cui. Advanced electrospun hydrogel fibers for wound healing. Composites Part B: Engineering 2021; 223, 109101.
M Sufiyan, P Kushwaha, M Ahmad, P Mandal and KK Vishwakarma. Scaffold-mediated drug delivery for enhanced wound healing: A review. AAPS PharmSciTech 2024; 25(5), 137.
J K Tee, CN Ong, BH Bay, HK Ho and DT Leong. Oxidative stress by inorganic nanoparticles. Wiley Interdisciplinary Reviews: Nanomedicine and Nanobiotechnology 2016; 8(3), 414-438.
SM Häffner and M Malmsten. Interplay between amphiphilic peptides and nanoparticles for selective membrane destabilization and antimicrobial effects. Current Opinion in Colloid & Interface Science 2019; 44, 59-71.
AK Tiwari, PC Pandey, MK Gupta and RJ Narayan. Nano–bio interaction and antibacterial mechanism of engineered metal nanoparticles: fundamentals and current understanding. Journal of Cluster Science 2025; 36(1), 1-33.
A R Bilia, V Piazzini, L Risaliti, G Vanti, M Casamonti, M Wang and M C Bergonzi. Nanocarriers: a successful tool to increase solubility, stability and optimize bioefficacy of natural constituents. Current Medicinal Chemistry 2019; 26(24), 4631-4656.
D Yadav, K Sandeep, D Pandey and RK Dutta. Liposomes for drug delivery. Journal of Biotechnology & Biomaterials 2017; 7(4), 276.
M Saffari, FH Shirazi and HR Moghimi. Terpene-loaded liposomes and isopropyl myristate as chemical permeation enhancers toward liposomal gene delivery in lung cancer cells: a comparative study. Iranian Journal of Pharmaceutical Research 2016; 15(3), 261.
B Kumar, M Pandey, R Aggarwal and PK Sahoo. A comprehensive review on invasomal carriers incorporating natural terpenes for augmented transdermal delivery. Future Journal of Pharmaceutical Sciences 2022; 8(1), 50.
AC Paiva-Santos, AL Silva, C Guerra, D Peixoto, M Pereira-Silva, M Zeinali, F Mascarenhas-Melo, R Castro and F Veiga. Ethosomes as nanocarriers for the development of skin delivery formulations. Pharmaceutical Research 2021; 38(6), 947-970.
EM Elmowafy, M Tiboni and ME Soliman. Biocompatibility, biodegradation and biomedical applications of poly(lactic acid)/poly(lactic-co-glycolic acid) micro and nanoparticles. Journal of Pharmaceutical Investigation 2019; 49, 347-380.
MM El-Hammadi, AL Small-Howard, M Fernández-Arévalo and L Martín-Banderas. Development of enhanced drug delivery vehicles for 3 cannabis-based terpenes using poly(lactic-co-glycolic acid)-based nanoparticles. Industrial Crops and Products 2021; 164, 113345.
P Patel, N Vyas and M Raval. Safety and toxicity issues of polymeric nanoparticles: A serious concern. In: M Rai, M Patel and R Patel (Eds.). Nanotechnology in medicine: Toxicity and safety. Wiley, New Jersey, United States, 2021, p. 156-173.
E Lasoń. Topical administration of terpenes encapsulated in nanostructured lipid-based systems. Molecules 2020; 25(23), 5758.
R H Müller, K Mäder and S Gohla. Solid lipid nanoparticles (SLN) for controlled drug delivery - a review of the state of the art. European Journal of Pharmaceutics and Biopharmaceutics 2000; 50(1), 161-177.
SV Khairnar, P Pagare, A Thakre, AR Nambiar, V Junnuthula, MC Abraham, P Kolimi, D Nyavanandi and S Dyawanapelly. Review on the scale-up methods for the preparation of solid lipid nanoparticles. Pharmaceutics 2022; 14(9), 1886.
M Kumari and D K Nanda. Potential of curcumin nanoemulsion as antimicrobial and wound healing agent in burn wound infection. Burns 2023; 49(5), 1003-1016.
CR Garcia, MH Malik, S Biswas, VH Tam, KP Rumbaugh, W Li and X Liu. Nanoemulsion delivery systems for enhanced efficacy of antimicrobials and essential oils. Biomaterials Science 2022; 10(3), 633-653.
TN Barradas and KG de Holanda e Silva. Nanoemulsions of essential oils to improve solubility, stability and permeability: A review. Environmental Chemistry Letters 2021; 19(2), 1153-1171.
A Nabawy, JM Makabenta, S Schmidt-Malan, J Park, CH Li, R Huang, S Fedeli, AN Chattopadhyay, R Patel and VM Rotello. Dual antimicrobial-loaded biodegradable nanoemulsions for synergistic treatment of wound biofilms. Journal of Controlled Release 2022; 347, 379-388.
L Li, Y Xie, J Wang, Q Sun, M Gao and C Li. Biofilm microenvironment-activated multimodal therapy nanoplatform for effective antibacterial treatment and wound healing. Acta Biomaterialia 2024; 183, 221-234.
S Fahimirad, F Fattahi, M Hatami and S Shabani. Nanotechnology-based biotherapeutics for physiological wound healing phases. Industrial Crops and Products 2025; 226, 120608.
SS Garg, R Dubey, S Sharma, A Vyas and J Gupta. Biological macromolecule-based nanoformulations for improving wound healing and bacterial biofilm-associated infection: A review. International Journal of Biological Macromolecules 2023; 247, 125636.
M Li, X He, R Zhao, Q Shi, Y Nian and B Hu. Hydrogels as promising carriers for the delivery of food bioactive ingredients. Frontiers in Nutrition 2022; 9, 1006520.
R Ajdary, BL Tardy, BD Mattos, L Bai and O J Rojas. Plant nanomaterials and inspiration from nature: Water interactions and hierarchically structured hydrogels. Advanced Materials 2021; 33(28), 2001085.
M Zagórska-Dziok, A Nowak, A Zgadzaj, E Oledzka, K Kędra, A E Wiącek and M Sobczak. New polymeric hydrogels with cannabidiol and α-terpineol as potential materials for skin regeneration synthesis and physicochemical and biological characterization. International Journal of Molecular Sciences 2024; 25(11), 5934.
I Negut, G Dorcioman and V Grumezescu. Scaffolds for wound healing applications. Polymers 2020; 12(9), 2010.
S Stratton, NB Shelke, K Hoshino, S Rudraiah and SG Kumbar. Bioactive polymeric scaffolds for tissue engineering. Bioactive Materials 2016; 1(2), 93-108.
WJ Duncan. Establishment and evolution of the Otago sheep animal models: 40 years of periodontal, implant and bone regeneration research. New Zealand Dental Journal 2024; 120(4), 149-161.
H Haidari, R Bright, Y Yu, K Vasilev and Z Kopecki. Development of microneedles for antimicrobial drug delivery: A comprehensive review on applications in wound infection management. Small Science 2024; 4(10), 2400158.
TS Alster and PM Graham. Microneedling: a review and practical guide. Dermatologic Surgery 2018; 44(3), 397-404.
L Sun, Z Zhang, Y Cao, Z Lu, Y Zhang, X He, Z Wang and R Pei. pH-responsive and on-demand hydrogen sulfide-releasing injectable hydrogel for wound healing promotion. Chemical Engineering Journal 2025; 509, 160904.
A Ullah, M Jang, H Khan, HJ Choi, S An, D Kim, YR Kim, UK Kim and GM Kim. Microneedle array with a pH-responsive polymer coating and its application in smart drug delivery for wound healing. Sensors and Actuators B: Chemical 2021; 345, 130441.
J Mei, D Xu, L Wang, L Kong, Q Liu and Q Li. Biofilm microenvironment-responsive self-assembly nanoreactors for all-stage biofilm-associated infection through bacterial cuproptosis-like death and macrophage re-rousing. Advanced Materials 2023; 35(36), e2303432.
D Kumar, S Pandey, J Shiekmydeen, M Kumar, S Chopra and A Bhatia. Therapeutic potential of microneedle-assisted drug delivery for wound healing: Current state of the art, challenges, and future perspective. AAPS PharmSciTech 2025; 26(1), 25.
S Dalleau, E Cateau, T Bergès, JM Berjeaud and C Imbert. In vitro activity of terpenes against Candida biofilms. International Journal of Antimicrobial Agents 2008; 31(6), 572-576.
P Kumari, N Arora, A Chatrath, R Gangwar, V Pruthi, KM Poluri and R Prasad. Delineating the biofilm inhibition mechanisms of phenolic and aldehydic terpenes against Cryptococcus neoformans. ACS Omega 2019; 4(18), 17634-17648.
MA Chowdhury, M Ashrafudoulla, SI Mevo, AS Mahamud, CS Reem, AJ Ha and SD Ha. Efficacy of orange terpene against Escherichia coli biofilm on beef and food contact surfaces. Food Research International 2024; 197, 115204.
C Salinas, G Florentín, F Rodríguez, N Alvarenga and R Guillén. Terpenes combinations inhibit biofilm formation in Staphylococcus aureus by interfering with initial adhesion. Microorganisms 2022; 10(8), 1527.
PSS Lima, AM Lucchese, HG Araujo-Filho, PP Menezes, AAS Araujo, LJ Quintans-Junior and JSS Quintans. Inclusion of terpenes in cyclodextrins: preparation, characterization and pharmacological approaches. Carbohydrate Polymers 2016; 151, 965-987.
MAR Martins, LP Silva, O Ferreira, B Schröder, JAP Coutinho and SP Pinho. Terpenes solubility in water and their environmental distribution. Journal of Molecular Liquids 2017; 241, 996-1002.
B Sapra, S Jain and AK Tiwary. Percutaneous permeation enhancement by terpenes: Mechanistic view. AAPS Journal 2008; 10, 120-132.
F Nourbakhsh, M Lotfalizadeh, M Badpeyma, A Shakeri and V Soheili. From plants to antimicrobials: Natural products against bacterial membranes. Phytotherapy Research 2022; 36(1), 33-52.
AY Abramov, MV Zamaraeva, AI Hagelgans, RR Azimov and OV Krasilnikov. Influence of plant terpenoids on the permeability of mitochondria and lipid bilayers. Biochimica et Biophysica Acta (BBA) - Biomembranes 2001; 1512(1), 98-110.
J Szymczak, L Sobotta, J Dlugaszewska, M Kryjewski and J Mielcarek. Menthol-modified zinc(II) phthalocyanine regioisomers and their photoinduced antimicrobial activity against Staphylococcus aureus. Dyes and Pigments 2021; 193, 109410.
M Kossakowska-Zwierucho, G Szewczyk, T Sarna and J Nakonieczna. Farnesol potentiates photodynamic inactivation of Staphylococcus aureus with the use of red light-activated porphyrin TMPyP. Journal of Photochemistry and Photobiology B: Biology 2020; 206, 111863.
Y Li and M X Wu. Visualization and elimination of polymicrobial biofilms by a combination of ALA-carvacrol-blue light. Journal of Photochemistry and Photobiology B: Biology 2022; 234, 112525.
M Lu, Y Li and MX Wu. Bacteria-specific pro-photosensitizer kills multidrug-resistant Staphylococcus aureus and Pseudomonas aeruginosa. Communications Biology 2021; 4(1), 408.
M Lu, S Wang, T Wang, S Hu, B Bhayana, M Ishii, Y Kong, Y Cai, T Dai, W Cui and MX Wu. Bacteria-specific phototoxic reactions triggered by blue light and phytochemical carvacrol. Science Translational Medicine 2021; 13(575), eaba3571.
P Manrique, I Montero, M Fernandez-Gosende, N Martinez, CH Cantabrana and D Rios-Covian. Past, present, and future of microbiome-based therapies. Microbiome Research Reports 2024; 3(2), 23.
MB Habib, G Batool, NA Shah, T Muhammad, NS Akbar and A Shahid. Biofilm-mediated infections: Novel therapeutic approaches and harnessing artificial intelligence for early detection and treatment. Microbial Pathogenesis 2025; 203, 107497.
F Pennisi, A Pinto, G E Ricciardi, C Signorelli and V Gianfredi. Artificial intelligence in antimicrobial stewardship: A systematic review and meta-analysis of predictive performance and diagnostic accuracy. European Journal of Clinical Microbiology & Infectious Diseases 2025; 44(3), 463-513.
Natouri, A Barzegar, A Nobakht, A Bagheri, F Eslami, S Jafarirad, N Tohidifar and B Nikzad. Bioactive stem cell-laden 3D nanofibrous scaffolds for tissue engineering. Heliyon 2024; 10(19), e38462.
S Yamakawa and K Hayashida. Advances in surgical applications of growth factors for wound healing. Burns & Trauma 2019; 7, 1-9.
IA Jang, J Kim and W Park. Endogenous hydrogen peroxide increases biofilm formation by inducing exopolysaccharide production in Acinetobacter oleivorans DR1. Scientific Reports 2016; 6(1), 21121.
J Mei, D Xu, L Wang, L Kong, Q Liu, Q Li, X Zhang, Z Su, X Hu, W Zhu and M Ye. Biofilm microenvironment-responsive self-assembly nanoreactors for all-stage biofilm-associated infection through bacterial cuproptosis-like death and macrophage re-rousing. Advanced Materials 2023; 35(36), 2303432.
M Kim and J H Park. Isolation of Aloe saponaria-derived extracellular vesicles and investigation of their potential for chronic wound healing. Pharmaceutics 2022; 14(9), 1905.