Trends
Sci.
2026;
23(8):
12879
Comparative Study on Anti-Inflammatory Activity and Phytochemical Constituents of Essential Oil, Steamed and Non-Steamed Extracts of Zingiber Ottensii Valeton
Patcharaporn Muanrit1, Sumalee Panthong2, Saovapak Poomirat2,
Ubonwan Saesiw2 and Intouch Sakpakdeejaroen2,*
1Student of Doctor of Philosophy in Applied Thai Traditional Medicine, Faculty of Medicine, Thammasat University, Pathum Thani 12120, Thailand
2Department of Applied Thai Traditional Medicine, Faculty of Medicine, Thammasat University,
Pathum Thani 12120, Thailand
(*Corresponding author’s e-mail: [email protected])
Received: 3 December 2025, Revised: 9 January 2026, Accepted: 16 January 2026, Published: 15 March 2026
Abstract
The rhizome of Zingiber ottensii Valeton (ZT) has traditionally been used to treat wounds and various digestive disorders, including flatulence, stomachache, and peptic ulcers, as well as to alleviate inflammatory conditions. However, the impact of the steaming process - commonly employed in Thai folk medicine to prepare ZT rhizomes - on their anti-inflammatory activity and chemical constituents has not yet been scientifically investigated. Therefore, this study aimed to compare the anti-inflammatory activities, analyze the chemical composition, and quantify bioactive constituents of essential oil, including steamed and non-steamed extracts. In vitro anti-inflammatory activities were assessed by measuring NO inhibition using Griess reagent in RAW 264.7 cells, as well as IL-6, TNF-α and PGE2 production using an ELISA test kit. Chemical composition was analyzed using GC/MS, and bioactive constituents in the extracts were quantified by HPLC. Among all ZT extracts, the essential oil (ZTEO) exhibited the strongest inhibitory effect on NO and IL-6 production. In comparison, the non-steamed hexane (ZTNH) and ethanol (ZTNE) extracts were significantly more effective than the steamed hexane (ZTSH) and ethanol (ZTSE) extracts in suppressing these inflammatory mediators. Zerumbone also demonstrated remarkable efficacy in inhibiting NO and IL-6, being at least 57-fold more potent than diclofenac. GC-MS analysis revealed notable differences in the chemical profiles of steamed and non-steamed ZT extracts and essential oil. Zerumbone and α-humulene were identified as the major sesquiterpenes in all tested samples, while terpinen-4-ol was absent in both the steamed hexane and ethanol extracts. The HPLC results further confirmed a significant decrease in zerumbone content in the steamed extracts compared to their non-steamed counterparts. Overall, the steaming process appeared to reduce both the anti-inflammatory activity and zerumbone content of Zingiber ottensii rhizomes, suggesting that non-steamed rhizomes are preferable for preserving their bioactive compounds and therapeutic potential.
Keywords: Zingiber ottensii, Thai traditional medicine, Chemical constituent, Steamed extract, Anti-inflammation, HPLC, GC/MS, Zerumbone
Introduction
Inflammation is a complex biological response of the immune system to infection, injury, or harmful stimuli. This defense mechanism functions to eliminate the underlying cause of cellular damage, remove
necrotic tissue, and initiate tissue repair [1]. The process involves the migration of leukocytes, including neutrophils, monocytes, and macrophages, to the affected site. Simultaneously, various inflammatory mediators—such as nitric oxide (NO), prostaglandin E2 (PGE2), serotonin, bradykinin and histamine—are produced, along with pro-inflammatory cytokines including interleukin-6 (IL-6) and tumor necrosis factor-alpha (TNF-α) [2]. However, chronic or uncontrolled inflammation can lead to various inflammatory disorders, such as arthritis, cardiovascular disease, diabetes, neurodegenerative disorders, and cancer [3]. Although conventional non-steroidal anti-inflammatory drugs (NSAIDs) are effective, their long-term use is often associated with adverse side effects, including gastrointestinal lesions, bleeding, and peptic ulcers [4], prompting extensive research into safer, naturally derived anti-inflammatory agents.
The genus Zingiber (family Zingiberaceae) is a well-recognized source of potent medicinal plants widely used in traditional medicine. Among its species, Zingiber ottensii Valeton (ZT), known in Thai as Phlai Dum, is used locally throughout Southeast Asia to treat symptoms associated with fever, body aches, and general pain [5]. In Thailand, the rhizome of ZT has traditionally been used in folk medicine to treat wounds and digestive ailments, such as flatulence, stomachache, and peptic ulcers, as well as to relieve inflammatory conditions including bruises, sprains, and muscle pain [6,7].
Several studies have reported that ZT rhizome and leaf extracts exhibit anti-inflammatory [6,8,9], antiproliferative [7,10,11], antimicrobial [10], and antioxidant [12] activities. Previous phytochemical analyses of ZT extracts and essential oils have identified key bioactive constituents, including monoterpenes, sesquiterpenes and diarylheptanoids [5,13,14]. Among these, the sesquiterpene zerumbone appears to play a significant role in mediating these biological activities.
According to traditional practices employed by Thai folk doctors, the rhizomes of Zingiber species, such as ginger (Z. officinale) and Bengal ginger (Z. montanum), are commonly subjected to a steaming process before being used in remedies [15,16]. This technique is believed to enhance therapeutic efficacy, reduce potential irritability, and improve storage stability. It is well established that thermal processing can substantially alter the composition and phytochemical content of plant materials. Such modifications may either enhance or impair pharmacological properties through degradation or structural transformation, depending on the characteristics of the compounds and the thermal treatment conditions (e.g., duration, temperature, and moisture). For example, steaming has been reported to increase 6-gingerol content [16] and enhance the anti-inflammatory activity of ginger rhizome extract compared to its non-steamed counterpart [17]. However, no comprehensive investigation has compared the chemical composition and anti-inflammatory activities of extracts obtained from steamed versus non-steamed ZT rhizomes, leaving a significant gap in understanding the scientific basis of this traditional practice.
Therefore, in this study, we aimed to compare the anti-inflammatory activities of hexane and ethanol extracts obtained from steamed versus non-steamed ZT rhizomes, as well as the essential oil extracted via hydrodistillation from non-steamed ZT rhizomes. In addition, we analyzed the chemical composition of these samples using gas chromatography/mass spectrometry (GC/MS), and quantified their bioactive constituents through a validated high-performance liquid chromatography (HPLC) method recently developed by our research group [18].
Materials and methods
Chemicals and reagents
Dulbecco's Modified Eagle Medium (DMEM), Minimum Essential Medium (MEM), fetal bovine serum (FBS), Penicillin-Streptomycin (P/S) (10,000 U/mL), and 0.4% Trypan blue solution were purchased from Gibco, (New York, USA). Phosphoric acid (85 %w/w), dimethyl sulfoxide (DMSO), Albumin bovine fraction V powder, anti-dinitrophenyl bovine serum albumin (DNP-BSA), DNP-specific IgE (monoclonal anti-DNP), 4-nitrophenyl N-acetyl-β-D-glucosaminide (PNAG), lipopolysaccharide (LPS) from E. coli O55:B5 (Catalog No. L4005, Lot No. 034k4112), sulfanilamide, phosphate buffered saline (PBS), N-(1-Naphthy) ethylenediamine dihydrochloride, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl- tetrazolium bromide (MTT), hydrochloric acid (HCl), diclofenac (sodium salt) and all other chemicals not specifically mentioned were purchased from Merck Millipore (Darmstadt, Germany). Standard zerumbone was purchased from TCI (Product No. Z0060, Lot No. OMU8F, Tokyo, Japan). Solvents—including acetonitrile (HPLC grade), methanol (HPLC grade), purified water (HPLC grade), n-hexane (analytical grade), isopropanol (analytical grade) and 95% ethanol (analytical grade)—were obtained from RCI Labscan (Bangkok, Thailand).
Plant materials
Fresh rhizomes of ZT were obtained from a Thai herbal farmer in Wiang Haeng District, Chiang Mai Province, Thailand, in January 2023. A voucher specimen (TTM-c No. 1000774) was identified by an herbal medicine expert from the Department of Thai Traditional and Alternative Medicine (DTAM) and deposited at the herbarium of the Thai Traditional Medicine Research Institute, Department of Thai Traditional and Alternative Medicine, Bangkok, Thailand.
Preparation of ZT rhizome extracts and its essential oil
Fresh rhizomes of ZT were cleaned and divided into 3 portions. The first portion (non-steamed) was dried at 50 °C for 48 h and ground into coarse powder. The second portion was subjected to steamed using an adapted method, as previously reported [15,19]. To ensure high reproducibility and consistency, a standardized high-intensity model was employed using an autoclave at 121 °C for 20 min. This controlled environment was chosen to eliminate variables common in traditional steaming, such as fluctuations in ambient pressure, temperature and steam density. Following the steaming treatment, the ZT rhizomes were dried at 50 °C for 48 h and ground into a coarse powder. Both non-steamed and steamed ZT powders were then partitioned for extraction via maceration and decoction.
For maceration, the crude ZT powders were extracted with either n-hexane or 95% ethanol for 3 days. The extracts were filtered through Whatman No. 1 filter paper. This maceration process was repeated twice, for a total of 3 extractions. Finally, the extracts were combined, concentrated under reduced pressure using a rotary evaporator, and dried in a vacuum dryer [20].
For decoction, the crude ZT powders were boiled with deionized water for 15 min following traditional folk medicine methods. The extracts were then filtered through a nylon filter sheet (200 mesh; 75 μm pore-size). The decoction process was repeated twice, for a total of 3 extractions. Finally, the extracts were combined and dried using a lyophilizer [20].
The remaining portion of the fresh rhizomes was used to extract the essential oil by hydrodistillation for 6 h using a Clevenger-type apparatus. The isolated ZT essential oils were dried over anhydrous sodium sulfate. All extracts were stored at −20 °C for further experiments.
Anti-inflammatory activity
Cell culture
The RAW 264.7 murine macrophage cell line was obtained from the American Type Culture Collection (ATCC TIB-71). Cells were grown as a monolayer in DMEM supplemented with 10% (v/v) fetal bovine serum (FBS), 100 U/mL penicillin and 100 μg/mL streptomycin. Cultures were maintained at 37 °C in a humidified incubator with a 5% carbon dioxide (CO2). The culture medium was changed every 3 - 4 days, and cells were subcultured or used for experiments when they reached approximately 80% confluence. All experiments were performed using cells between passages 7 and 20, which were confirmed to be negative for mycoplasma contamination.
Inhibitory effect on NO production in LPS-stimulated RAW 264.7 cells
The inhibition of NO production was evaluated by measuring the concentration of the end product, nitrite (NO2-), using Griess reagent (containing 0.1% N-(1-naphthyl) ethylenediamine dihydrochloride and 1% sulfanilamide in 2.5% phosphoric acid solution). Briefly, RAW 264.7 cells were seeded into a 96-well plate at a density of 1×105 cells/well and incubated for 24 h at 37 °C with 5% CO2 to allow cell attachment. Stock solutions of ZT extracts, zerumbone and the positive control (diclofenac sodium) were prepared in 100% DMSO. These were further diluted in fresh culture medium to obtain 5 different final concentrations ranging between 1.5625 and 50 µg/mL; zerumbone was diluted to a range of 0.390625 to 6.25 µg/mL. Aqueous extracts were dissolved in sterile deionized water, filtered through a 0.22 µm sterile filter, and similarly diluted in fresh culture medium to concentrations between 1.5625 and 50 µg/mL. The medium was then replaced with 100 µL of fresh medium containing 10 ng/mL LPS, and cells were treated with 100 µL of test samples at different concentrations. The maximum final concentration of DMSO in all wells, including the vehicle control, did not exceed 0.1% (v/v). After 24 h of incubation, 100 µL of supernatant was transferred to a new 96-well plate and mixed with an equal volume of Griess reagent. The optical density was measured at 570 nm [21,22].
Data obtained from 3 independent experiments (each concentration performed in quadruplicate wells) were analyzed using GraphPad Prism v.9 (GraphPad Software, San Diego, CA, USA). Percent inhibition values were calculated using the equation below. Dose-response curves were used to determine the half-maximal inhibitory concentration for 50% (IC50) of NO production by plotting percent inhibition against the sample concentrations using non-linear regression. Results are expressed as mean IC50 values ± standard deviation (SD).

Meanwhile, the potential cytotoxic effects of the test samples on RAW 264.7 cells were assessed using the MTT colorimetric assay to ensure that the inhibition of NO inhibition, including other inflammatory mediators, was not confounded by cell death. Briefly, 10 µL of MTT solution (0.5% w/v in PBS) was added to each well of the 96-well plate. After 2 h of incubation protected from light, the supernatant was removed, and 100 µL of isopropanol containing 0.04 M HCl was added to each well to dissolve the purple formazan product. Optical density was measured at 570 nm using a microplate reader [21,22]. Results were expressed as percentage cell viability relative to untreated cells according to the equation below. Samples were considered non-cytotoxic when cell viability exceeded 70%.
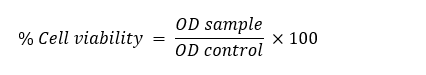
Inhibitory effect on IL-6 and TNF-α production in LPS-stimulated RAW 264.7 cells
The inhibition of IL-6 and TNF-α production was determined using ELISA kits (Cat. No. DY406 for IL-6 and DY410 for TNF-α, R&D Systems Inc., MN, USA) according to the manufacturer’s instructions. RAW 264.7 cells were grown and treated with LPS and test samples as described in the previous section. After 24 h of treatment, 100 µL of supernatant was collected to measure IL-6 and TNF-α production.
A 96-well plate was coated overnight at 4 °C with mouse IL-6 or TNF-α capture antibodies. The plate was then washed 3 times with wash buffer and blocked with 5% bovine serum albumin in PBS for 2 h at room temperature. After blocking, 100 µL of supernatant was added to each well and incubated for 2 h at room temperature, followed by washing. Streptavidin-HRP was subsequently added and incubated for 30 minutes protected from light, followed by another washing step. TMB (3,3′,5,5′-Tetramethylbenzidine) substrate was then added and incubated for 30 minutes, and the reaction was stopped by adding 50 µL of 1 M sulfuric acid (H2SO4). Optical density was measured at 450 nm [21,23].
IL-6 and TNF-α concentrations were calculated from the absorbance of each sample using a standard curve of the recombinant protein. Results were expressed as the percentage inhibition of IL-6 and TNF-α production according to the equation below. Dose-response curves were used to determine the IC50 values for of IL-6 and TNF-α production, as previously described.

Results were expressed as percentage inhibition according to the equation below. Dose-response curves were used to determine the IC50 of IL-6 and TNF-α production by plotting percent inhibition against the sample concentrations using non-linear regression, as previously described. Results are expressed as mean IC50 values ± standard deviation (SD).
Inhibitory effect on PGE2 production in LPS-stimulated RAW 264.7 cells
The inhibition of PGE2 production was determined using ELISA kits (Cat. No. 514010, Cayman Chemical, MI, USA) according to the manufacturer’s instructions. The RAW 264.7 cells were grown and treated with LPS and test samples as described in the previous section. After 24 h of treatment, 100 µL of supernatant was collected to measure PGE2 production.
In a 96-well plate, 50 µL of supernatant was mixed with 50 µL of PGE2 tracer and 50 µL of PGE2 monoclonal antibody. The plate was covered with plastic film and incubated at 4 °C for 18 h. After incubation, the plate was washed 5 times with 400 µL/well of wash buffer. Subsequently, 200 µL of Ellman's reagent was added to each well. The plate was wrapped in plastic film and incubated in the dark for 60 - 90 min. Optical density was measured at 412 nm [23]. Experiments were performed using quadruplicate wells for each concentration, and 3 independent experiments were performed.
Results were expressed as percentage inhibition according to the equation below. Dose-response curves were used to determine the IC50 of PGE2 production, as previously described. Results are expressed as mean IC50 values ± standard deviation (SD).
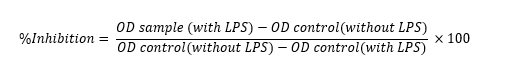
Chemical profiling by GC/MS analysis
The chemical profiling of the ZT extracts and essential oil was analyzed using a GC/MS system (GCMS-TQ™ 8040 NX; Shimadzu Corporation, Kyoto, Japan) coupled with an autosampler (AOC-20s Plus) and an auto injector (AOC-20s Plus), following an adaptation of previously reported methodology [14]. Briefly, samples were injected into a GC capillary column, Rxi-5Sil MS (30 m×0.25 mm, 0.25 µm film thickness). The oven temperature was held at 60 °C for 3 min, then programmed as follows: 1 °C/min to 80 °C, 3 °C/min to 120 °C, 4 °C/min to 220 °C, 10 °C/min to 300 °C and held at 300 °C for 5 min.
The injector temperature was 250 °C in split mode (split ratio 1:7), with a linear velocity of 40.0 cm/s pressure of 72.6 kPa; total flow of 12.6 mL/min, column flow of 1.20 mL/min, and purge flow of 3.0 mL/min. Helium was used as the carrier gas. Mass spectra were acquired in electron ionization (EI) mode at detector volage relative to the tuning results, with an ion source temperature of 270 °C and interface temperature of 300 °C, scanning from m/z 45 to m/z 700 (solvent cut time 3 min). GC/MS analysis was done in duplicate. Compounds were identified by comparing each peak’s mass spectrum with a standard library (the Wiley Registry 11th edition/NIST 2017 library, W11N17).
Determination of zerumbone in ZT extracts and essential oil using HPLC
The amount of zerumbone in the ZT extracts and essential oil was determined using a validated method, as previously reported [18]. The HPLC system (Agilent® 1200; Agilent Technologies, CA, USA) consisted of a solvent degasser (G1322A), a quaternary pump (G1311A), an autosampler (G1329A), a thermostatted column compartment (G1316A) and a diode array detector (G1315D). The validated HPLC method yield a linear response (R2 = 0.9993) from 10 to 1,000 µg/mL. The LOD and LOQ were determined to be 2.89 and 8.75 µg/mL, respectively. Method accuracy was evidenced by recoveries of 96.29–103.25%, and both intra-day and inter-day precision showed a %RSD < 2.0%, confirming the method’s reliability [18].
To do so, samples were dissolved in methanol (5 mg/mL) and filtered through a 0.45 μm nylon membrane filter. They were then injected into a reversed-phase column, Shim-pack GIST (4.60×150 mm, 5 µm, 100 Å), with a guard column of the same material. The mobile phases consisted of 0.1% v/v phosphoric acid in water (A) and acetonitrile (B) with the following gradient elution: 0 min, 90:10; 10 min, 90:10; 30 min, 70:30; 45 min, 60:40; 53 min, 95:5; 60 min, 95:5; 60.1 min, 90:10; 65 min, 90:10. The flow rate was 1 mL/min with UV detection at 215 nm. The operating temperature was maintained at room temperature.
A calibration curve of standard zerumbone was prepared with concentrations ranging from 10 to 1,000 µg/mL and analyzed in triplicate. Chromatographic data were analyzed using ChemStation® software (revision B.04.02 SP1). The content of zerumbone in the ZT extracts and essential oil was calculated by correlating the peak area with the calibration curve of standard zerumbone.
Statistical analysis
All experiments were performed in 3 independent experiments, and results are expressed as the Mean ± SD. Statistical analysis was conducted using an independent samples t-test or a 2-Way analysis of variance (ANOVA) followed by Tukey’s post-hoc test., depending on the experimental design. Differences were considered statistically significant at p < 0.05.
Results and discussion
Percentage yields of ZT rhizome extracts and its essential oil
Among all samples, the water extracts of the ZT rhizome produced the highest yields, followed by 95% ethanol and n-hexane. Furthermore, the steaming process appeared to reduce the extraction efficiency across all solvents compared to their non-steamed counterparts. Meanwhile, the essential oil obtained via hydrodistillation resulted in the lowest yield, as shown in Table 1.
Table 1 Percentage yields of non-steamed and steamed ZT extracts, and essential oil
Samples |
Extracts |
Code |
% Yield |
ZTNH |
1.67 |
||
ZTNE |
6.87 |
||
Water |
ZTNW |
11.90 |
|
n-Hexane |
ZTSH |
1.01 |
|
95% Ethanol |
ZTSE |
4.52 |
|
Water |
ZTSW |
8.83 |
|
ZT essential oil |
- |
ZTEO |
0.35 |
Anti-inflammatory effects of ZT rhizome extracts and its essential oil
The ZT rhizome extracts and their essential oil were evaluated for their inhibitory effects on inflammatory mediator production in LPS-stimulated RAW 264.7 cells. Among all tested samples, the essential oil (ZTEO; IC50 = 2.26 ± 0.31 μg/mL) exhibited the most potent inhibition of NO production (Table 2). Both non-steamed hexane and ethanol extracts (IC50 = 2.74 ± 0.10 μg/mL for ZTNH and 3.99 ± 0.14 μg/mL for ZTNE) also showed strong NO inhibition, whereas their steamed counterparts were less effective (IC50 = 4.02 ± 0.71 μg/mL for ZTSH and 8.01 ± 0.94 μg/mL for ZTSE). Zerumbone (IC50 = 0.82 ± 0.06 μg/mL) demonstrated particularly strong activity, exceeding that of diclofenac (IC50 = 46.72 ± 1.74 μg/mL). In contrast, both water extracts did not suppress NO production at the tested concentrations (IC50 > 50 μg/mL).
Regarding cytotoxicity, MTT assay results confirmed that all tested samples were non-toxic to RAW 264.7 cells, with cell viability consistently greater than 70%.
For IL-6 production, ZTEO (IC50 = 9.42 ± 0.07 μg/mL) exhibited the highest inhibitory activity among all extracts (Figure 1). Consistent with the NO results, non-steamed extracts showed stronger IL-6 inhibition (IC50 = 9.71 ± 0.13 μg/mL for ZTNH and 21.81 ± 0.33 μg/mL for ZTNE) compared to the steamed extracts (IC50 = 12.15 ± 0.46 μg/mL for ZTSH, 34.16 ± 0.18 μg/mL for ZTSE). Zerumbone (IC50 = 1.37 ± 0.06 μg/mL) again demonstrated superior inhibitory potency, whereas diclofenac and both water extracts could not suppress IL-6 production at the tested concentrations (IC50 > 50 μg/mL).
Regarding TNF-α production, all ZT extracts exhibited only limited inhibitory activity at their maximum non-toxic concentrations tested. As shown in Table 3, the non-steamed extracts showed slightly higher inhibition (% inhibition = 38.34 ± 1.11% for ZTNH and 33.86 ± 0.88% for ZTNE) compared with the steamed extracts (% inhibition = 32.56 ± 1.17% for ZTSH and 27.53 ± 1.23% for ZTSE; % inhibition at 50 μg/mL) and ZTEO (% inhibition = 15.62 ± 0.35%). Both water extracts (% inhibition = 5.22 ± 0.48 μg/mL for ZTNW and 0.62 ± 0.29 μg/mL for ZTNW) barely inhibited TNF-α production. Moreover, zerumbone (% inhibition = 10.69 ± 0.90%) and diclofenac (% inhibition = 15.62 ± 0.35%) also showed weaker TNF-α inhibition compared to most ZT extracts.
A similar trend was observed for PGE2 production. All ZT extracts, ZTEO, and zerumbone showed negligible effects, with percent inhibition ranging from 0.27% to 13.09% at their maximum non-toxic concentrations tested (Table 3). By contrast, diclofenac, a known cyclooxygenase (COX) inhibitor, strongly suppressed PGE₂ production (IC50 = 2.05 ± 0.00 ng/mL).
Table 2 Inhibitory effect of ZT extracts and essential oil on NO production in LPS-stimulated RAW 264.7 cells (n = 3).
Sample |
% Inhibition on NO production at various concentrations (µg/mL) |
IC50 (µg/mL) |
||||||
50 |
25 |
12.5 |
6.25 |
3.125 |
1.5625 |
|||
ZTNH |
- |
96.82 ± 1.05 |
97.18 ± 0.73 |
77.65 ± 2.56 |
56.00 ± 2.52 |
32.55 ± 2.00 |
2.74 ± 0.10 a,*,†,# |
|
|
|
(73.65 ± 1.97) |
(78.62 ± 3.00) |
(83.30 ± 2.25) |
(84.43 ± 2.49) |
(91.46 ± 3.97) |
|
|
ZTSH |
- |
98.54 ± 0.74 |
98.25 ± 1.18 |
86.96 ± 2.64 |
61.94 ± 1.48 |
33.34 ± 1.36 |
4.02 ± 0.71 b,*,†,# |
|
|
|
(77.68 ± 3.24) |
(86.10 ± 0.33) |
(88.06 ± 3.40) |
(91.78 ± 2.37) |
(96.69 ± 1.69) |
|
|
ZTNE |
96.33 ± 0.65 |
95.13 ± 1.95 |
85.61 ± 3.43 |
63.12 ± 1.83 |
44.79 ± 2.00 |
- |
3.99 ± 0.14 b,*,†,# |
|
|
(95.87 ± 1.50) |
(96.77 ± 1.31) |
(92.83 ± 1.59) |
(91.90 ± 0.61) |
(92.12 ± 1.95) |
|
|
|
ZTSE |
96.82 ± 0.82 |
92.86 ± 1.29 |
74.61 ± 2.08 |
42.91 ± 3.42 |
23.15 ± 0.94 |
- |
8.01 ± 0.94 c,*,†,# |
|
|
(85.72 ± 0.62) |
(88.56 ± 1.64) |
(88.99 ± 1.35) |
(90.31 ± 0.69) |
(93.32 ± 2.11) |
|
|
|
ZTNW |
13.63 ± 0.36 |
13.56 ± 0.82 |
12.50 ± 0.09 |
10.52 ± 1.49 |
9.54 ± 0.90 |
- |
> 50 |
|
|
(86.67 ± 1.56) |
(85.86 ± 1.74) |
(89.34 ± 1.51) |
(91.40 ± 2.08) |
(91.03 ± 1.17) |
|
|
|
ZTSW |
6.85 ± 1.56 |
6.43 ± 1.79 |
6.61 ± 1.37 |
5.51 ± 1.82 |
4.64 ± 1.32 |
- |
> 50 |
|
|
(84.56 ± 2.88) |
(86.57 ± 1.36) |
(87.64 ± 1.40) |
(87.43 ± 2.07) |
(93.63 ± 2.57) |
|
|
|
ZTEO |
- |
94.95 ± 1.99 |
96.22 ± 1.61 |
86.12 ± 3.41 |
62.42 ± 1.68 |
45.77 ± 2.51 |
2.26 ± 0.32 †,# |
|
|
|
(81.58 ± 1.58) |
(86.13 ± 2.00) |
(93.19 ± 2.07) |
(93.66 ± 3.92) |
(95.09 ± 3.95) |
|
|
Diclofenac |
52.45 ± 0.73 |
35.23 ± 2.29 |
25.08 ± 2.07 |
18.09 ± 2.96 |
14.63 ± 2.46 |
- |
46.72 ± 1.74 |
|
|
(74.48 ± 3.63) |
(83.57 ± 0.97) |
(86.80 ± 2.32) |
(86.43 ± 3.78) |
(94.10 ± 0.93) |
|
|
|
Sample |
6.25 |
3.125 |
1.5625 |
0.78125 |
0.390625 |
|
IC50 (µg/mL) |
|
Zerumbone |
97.67 ± 0.41 |
96.32 ± 0.82 |
74.43 ± 3.68 |
48.21 ± 2.89 |
23.96 ± 0.18 |
|
0.82 ± 0.06 # |
|
|
(79.29 ± 1.77) |
(83.96 ± 3.02) |
(86.87 ± 2.98) |
(89.02 ± 2.50) |
(92.39 ± 3.46) |
|
|
|
Data are expressed as Mean ± SD (n = 3 independent experiments). Values followed by different superscript letters (a-c) indicate significant differences (p < 0.05) based on 2-Way ANOVA (solvent × steaming interaction) followed by Tukey’s post-hoc test. Independent samples t-tests were used to compare all individual ZT extracts with ZTEO (*, p < 0.05), all individual ZT samples with zerumbone (†, p < 0.05), and all samples with diclofenac (#, p < 0.05). Values reported as > 50 µg/mL were considered non-inhibitory at the highest concentration tested and were excluded from the statistical analysis.

Figure 1 Inhibitory effect of ZT extracts, essential oil and zerumbone on IL-6 production in LPS-stimulated RAW 264.7 cell. Data are expressed as Mean ± SD (n = 3 independent experiments). Cell viability at all tested concentrations was exceed 70%. Values followed by different superscript letters (a-d) indicate significant differences (p < 0.05) based on 2-Way ANOVA (solvent×steaming interaction) followed by Tukey’s post-hoc test. Independent samples t-tests were used to compare all individual ZT extracts with ZTEO (*, p < 0.05) and all individual ZT samples with zerumbone (#, p < 0.05). Values reported as > 50 µg/mL were considered non-inhibitory at the highest concentration tested and were excluded from the statistical analysis
Table 3 Inhibitory effect of ZT extracts and essential oil on TNF-α and PGE2 production in LPS-stimulated RAW 264.7 cells (n = 3).
Sample |
Tested concentration |
% Inhibition on inflammatory mediators (% Cell viability at various concentrations) |
|
(µg/mL) |
TNF-α |
PGE2 |
|
ZTNH |
25 |
38.34 ± 1.11 |
8.60 ± 0.28 |
|
|
(80.09 ± 2.83) |
(77.84 ± 1.98) |
ZTSH |
25 |
32.56 ± 1.17 |
6.02 ± 0.72 |
|
|
(76.65 ± 1.81) |
(78.96 ± 2.91) |
ZTNE |
50 |
33.86 ± 0.88 |
12.32 ± 0.90 |
|
|
(85.81 ± 1.63) |
(83.48 ± 2.39) |
ZTSE |
50 |
27.53 ± 1.23 |
0.79 ± 0.08 |
|
|
(94.02 ± 1.84) |
(90.05 ± 2.73) |
ZTNW |
50 |
5.22 ± 0.48 |
0.46 ± 0.07 |
|
|
(93.15 ± 1.82) |
(90.47 ± 1.96) |
ZTSW |
50 |
0.62 ± 0.29 |
0.27 ± 0.04 |
|
|
(86.47 ± 2.41) |
(89.38 ± 1.37) |
ZTEO |
25 |
24.65 ± 0.68 |
13.09 ± 1.54 |
|
|
(84.30 ± 2.02) |
(82.27 ± 2.47) |
Zerumbone |
6.25 |
10.69 ± 0.90 |
5.93 ± 0.08 |
|
|
(82.40 ± 2.23) |
(84.59 ± 2.37) |
Diclofenac |
50 |
15.62 ± 0.35 |
IC50 = 2.05 ± 0.00 ng/mL |
|
|
(78.56 ± 1.89) |
|
The inhibitory effect on TNF-α and PGE2 production was tested at the maximum concentration, which showed non-toxic on RAW264.7 cells (Cell viability > 70%.). Data are expressed as Mean ± SD (n = 3 independent experiments).
These findings highlight the potent anti-inflammatory properties of ZT rhizome extracts and essential oil, primarily through the inhibition of key pro-inflammatory mediators, NO and IL-6. The most notable effect was the strong suppression of NO production. Among the tested samples, the essential oil (ZTEO) was the most effective, followed by the non-steamed hexane (ZTNH) and ethanol (ZTNE) extracts. The observed suppression of NO production may be attributed to the downregulation of inducible nitric oxide synthase (iNOS) expression, a crucial step in the inflammatory cascade [24].
Previous studies have highlighted the possible anti-inflammatory potential of ZT rhizomes. In Wistar rats, ZT essential oil significantly reduced hind paw swelling and inflammation severity when applied prior to carrageenan-induced inflammation [6]. This effect is hypothesized to be associated with the presence of sesquiterpene and monoterpene compounds, which are abundant in Zingiber species. In another study, crude protein isolated from ZT rhizomes was found to strongly inhibited NO production in RAW 264.7 cells co-stimulated with LPS and interferon-gamma (IFN-γ) and also reduced the expression levels of iNOS, IL-6, and TNF-α [9].
Likewise, zerumbone, one of the main phytochemicals in the ZT rhizomes, demonstrated potent inhibition of NO and IL-6 production, with an efficacy at least 57-fold greater than the standard anti-inflammatory drug diclofenac. These results suggests that zerumbone may contribute to the anti-inflammatory activity observed in ZT extracts. Such a possibility is consistent with literatures identifying zerumbone as a potential inhibitor of the nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) pathway, which regulates the transcription of iNOS and IL-6 [25,26].
Despite their strong inhibitory effects on NO and IL-6, zerumbone and all ZT extracts exhibited limited activity against TNF-α and PGE2 at their maximum non-toxic concentrations tested. The negligible effect on PGE2 production suggests that they may not significantly interfere with the cyclooxygenase (COX) pathway, the primary molecular target of the positive control, diclofenac. However, it should be noted that the choice to limit tested concentrations in these experiments was dictated by the observed cytotoxicity in RAW 264.7 cells; higher concentrations were not pursued, as they compromised cell viability, which would have led to unreliable inhibitory data.
A notable observation from our results is that the steaming process was associated with a reduction in the anti-inflammatory activity of ZT rhizome, as the ZTSH and ZTSE extracts exhibited diminished inhibitory effects on NO and IL-6 compared to their non-steamed counterparts. This reduction may be attributed to the potential heat sensitivity of the anti-inflammatory constituents, particularly zerumbone. Thermal processing can either enhance or reduce the biological activity of plants and their active compounds through degradation or structural transformation, with the extent of the effect depending on the processing method, duration, temperature, plant species and phytochemical composition [27,28].
For example, thermal treatment via roasting significantly decreased the antibacterial activity of turmeric rhizomes compared to fresh samples [29]. By contrast, the anti-inflammatory activity of steamed ginger evaluated in Helicobacter pylori-infected gastric epithelial cells revealed a significant reduction in pro-inflammatory cytokines, including IL-8, TNF-α, IL-6, iNOS, and IFN-γ, which was more potent than that of non-steamed ginger extract [30]. These varying outcomes highlight the necessity of optimizing extraction or processing methods to preserve the bioactive constituents and maintain therapeutic efficacy.
Chemical profiling of ZT rhizome extracts and essential oil by GC/MS analysis
GC/MS analysis of volatile components revealed notable differences in the chemical profile of ZT rhizome extracts and essential oil depending on thermal pretreatment (steamed vs. non-steamed) and extraction method (hexane, ethanol, and hydrodistillation). as shown in Table 4 and Figure 2.
Across all extracts, zerumbone and α-humulene were among the main monocyclic sesquiterpene constituents, although their relative peak areas varied significantly depending on preparation conditions, whereas, β-elemene was a minor monocyclic sesquiterpene. Bicyclic sesquiterpene (β-caryophyllene), oxygenated sesquiterpenes (humulene epoxide I and II), sesquiterpene alcohols (γ-eudesmol and β-eudesmol), and a monoterpene alcohol (trans-verbenol) were also detected in all extracts.
In the hexane extracts, ZTNH contained similar chemical compounds to ZTSH but in different proportions. The 6 major shared constituents were zerumbone (42.56% and 48.78%), α-humulene (26.53% and 32.54%), (E)-labda-8(17),12-diene-15,16-dial (12.30% and 3.94%), β-eudesmol (1.75% and 2.03%), cryptomeridiol (1.59% and 1.45%), and β-caryophyllene (1.56% and 2.05%). Notably, terpinen-4-ol, which was present in ZTNH (5.45%), was absent in ZTSH following the steaming of ZT rhizomes.
In the case of ethanol extracts, a slightly different pattern was observed. In both ZTNE and ZTSE, the 6 major constituents were zerumbone (52.99% and 65.27%), α-humulene (30.02% and 19.98%), cryptomeridiol (2.13% and 2.91%), β-eudesmol (1.76% and 3.50%), β-caryophyllene (1.34% and 1.82%), and humulene epoxide I (1.20% and 1.06%), Following steam pretreatment, terpinen-4-ol, which was present in ZTNE (4.88%), was absent in ZTSE. Additionally, compared with the hexane extracts, certain sesquiterpenes (α-gurjunene and caryophyllene oxide) and the diterpene (E)-labda-8(17),12-diene-15,16-dial were not detected in either ethanol extract.
For the essential oil, ZTEO was dominated by monoterpenes, with sabinene (19.63%), terpinen-4-ol (16.76%), β-pinene (10.14%), and α-humulene (7.80%) as the major components. The sesquiterpene zerumbone (15.68%) was also present in a high proportion, while other monoterpenes such as γ-terpinene, α-pinene, 4-carene, 1,8-cineole, α-terpineol, and D-limonene were detected only in low amounts.
Table 4 Chemical compositions identified in ZT extracts and essential oil using GC/MS analysis.
No. |
RT (min) |
Compound a |
Relative peak area (%) |
||||
ZTNH |
ZTSH |
ZTNE |
ZTSE |
ZTEO |
|||
1 |
6.577 |
α-Thujene |
ND |
ND |
ND |
ND |
0.53 |
2 |
6.869 |
α-Pinene |
ND |
ND |
ND |
ND |
2.25 |
3 |
8.697 |
Sabinene |
ND |
ND |
ND |
ND |
19.63 |
4 |
8.908 |
β-Pinene |
ND |
ND |
ND |
ND |
10.14 |
5 |
9.570 |
β-Myrcene |
ND |
ND |
ND |
ND |
0.98 |
6 |
10.480 |
α-Phellandrene |
ND |
ND |
ND |
ND |
0.50 |
7 |
11.152 |
4-Carene |
ND |
ND |
ND |
ND |
2.79 |
8 |
11.629 |
o-Cymene |
ND |
ND |
ND |
ND |
1.09 |
9 |
11.942 |
d-Limonene |
ND |
ND |
ND |
ND |
1.34 |
10 |
12.121 |
1,8-Cineole |
ND |
ND |
ND |
ND |
2.44 |
11 |
13.254 |
β-Ocimene |
ND |
ND |
ND |
ND |
0.52 |
12 |
14.036 |
γ-Terpinene |
ND |
ND |
ND |
ND |
4.70 |
13 |
14.889 |
4-Thujanol |
ND |
ND |
ND |
ND |
0.63 |
14 |
16.140 |
Terpinolene |
ND |
ND |
ND |
ND |
0.99 |
15 |
19.452 |
trans-p-Menth-2-enol |
ND |
ND |
ND |
ND |
0.75 |
16 |
24.705 |
Terpinen-4-ol |
5.45 |
ND |
4.88 |
ND |
16.76 |
17 |
26.013 |
α-Terpineol |
ND |
ND |
ND |
ND |
1.35 |
18 |
38.633 |
α-Gurjunene |
0.16 |
0.25 |
ND |
ND |
ND |
19 |
39.288 |
β-Caryophyllene |
1.56 |
2.05 |
1.34 |
1.82 |
0.37 |
20 |
40.865 |
α-Humulene |
26.53 |
32.54 |
30.02 |
19.98 |
7.80 |
21 |
44.353 |
β-Elemene |
0.60 |
0.73 |
0.39 |
0.41 |
1.42 |
22 |
45.426 |
Caryophyllene oxide |
0.32 |
0.42 |
ND |
ND |
0.21 |
23 |
45.999 |
Humulene epoxide I |
1.1 |
1.33 |
1.16 |
1.03 |
0.88 |
24 |
46.345 |
Humulene epoxide II |
1.26 |
1.4 |
0.96 |
0.69 |
0.49 |
25 |
47.082 |
γ-Eudesmol |
0.65 |
0.89 |
0.75 |
0.87 |
0.79 |
26 |
47.792 |
β-Eudesmol |
1.75 |
2.03 |
1.76 |
3.5 |
1.59 |
27 |
48.535 |
trans-Verbenol |
1.32 |
1.15 |
1.16 |
1.04 |
0.81 |
28 |
50.128 |
Zerumbone |
42.56 |
48.78 |
52.99 |
65.27 |
15.68 |
29 |
52.397 |
Cryptomeridiol |
1.59 |
1.45 |
2.13 |
2.91 |
ND |
30 |
64.286 |
(E)-Labda-8(17),12-diene-15,16-dial |
12.30 |
3.94 |
ND |
ND |
ND |
a All compounds shown in the table have a similarity of more than 80%, which were identified by comparing the MS spectra of each peak with a standard library (W11N17).
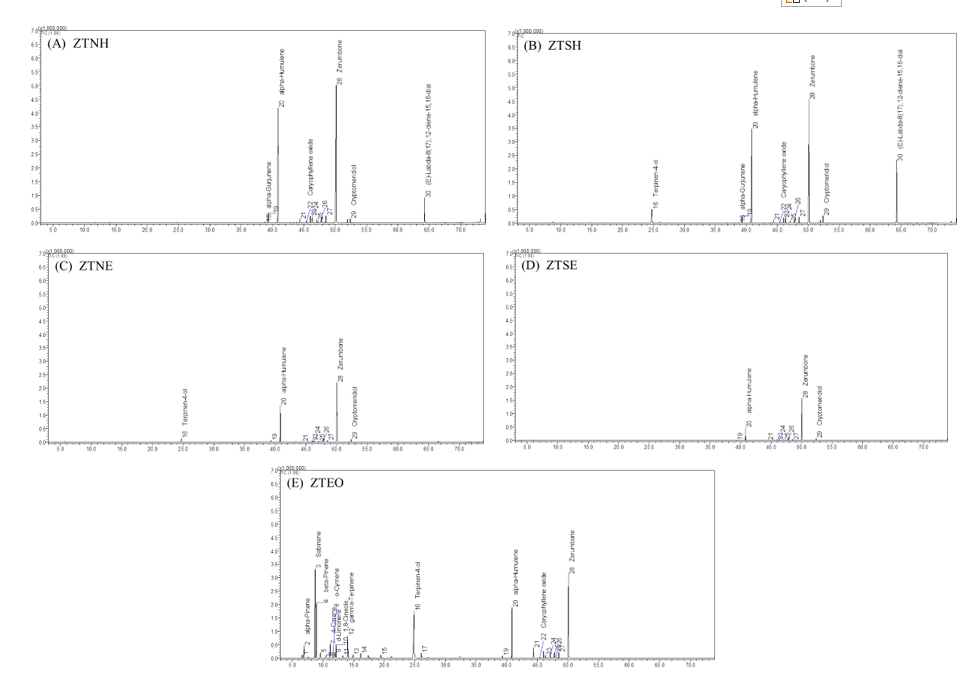
Figure 2 GC/MS chromatograms of some chemical compositions identified in (A) ZTNH, (B) ZTSH, (C) ZTNE, (D) ZTSE and (E) ZTEO.
The chemical profile of ZTEO in our study aligns with previous findings [5,14], revealing an abundance of monoterpenes, including sabinene, β-pinene, and terpinen-4-ol, as well as sesquiterpenes, particularly zerumbone and α-humulene. In contrast, the solvent extracts (ZTNH, ZTSH, ZTNE, and ZTSE) were predominantly composed of sesquiterpenes, primarily zerumbone and α-humulene, suggesting that these compounds are more selectively extracted by organic solvents. Furthermore, the consistent presence of zerumbone across all tested ZT extracts and essential oil supports its identification as a major bioactive marker of the ZT rhizome, consistent with the observed anti-inflammatory activity against NO and IL-6 production.
Steaming pretreatment, further altered the chemical profile of the ZT rhizome. Notably, the monoterpene alcohol, terpinen-4-ol, present in both non-steamed hexane and ethanol extracts, was entirely absent in their steamed counterparts, likely due to its thermal instability or high volatility. Differences were also observed based on solvent polarity, wherein the non-polar hexane extracts contained (E)-labda-8(17),12-diene-15,16-dial. This diterpene, known for its anti-inflammatory properties [31], was undetected in the more polar ethanol extracts and the essential oil, suggesting that the compound possesses notable lipophilic character and requires a non-polar solvent for effective extraction. Moreover, the presence of this diterpene may contribute to the superior inhibitory activity observed in the hexane extracts compared to the ethanol extracts.
Quantitative analysis of zerumbone in ZT rhizome extracts and its essential oil
The HPLC chromatograms revealed that zerumbone, was the major constituent in the hexane and ethanol extracts, as well as in the essential oil, eluting at a retention time of 33.3 min, as shown in Figure 3. Among all samples, ZTEO contained the highest amount of zerumbone (24.02 ± 0.22% w/w of extract), as presented in Table 5. Moreover, the zerumbone content in both the n-hexane and ethanol extracts of non-steamed ZT rhizome (22.18 ± 0.44% w/w of extract for ZTNH and 9.27 ± 0.34% w/w of extract for ZTNE) was significantly higher compared to their steamed counterparts (19.65 ± 0.27% w/w of extract for ZTSH and 7.34 ± 0.06% w/w of extract for ZTSE). This finding suggests that the steaming process likely induced thermal degradation, or volatilization of zerumbone [32]. Additionally, the n-hexane extracts contained a higher amount of zerumbone than the ethanol extracts, an outcome consistent with the lipophilic nature of zerumbone, which likely accounts for its absence in the water extracts.
Our findings further highlight a clear correlation between zerumbone content and the anti-inflammatory potency of the ZT extracts and essential oil, specifically regarding the inhibition of NO and IL-6 production. ZTEO, which possessed the highest percentage of zerumbone, exhibited the most potent inhibitory effect among all tested extracts. Conversely, the absence of zerumbone in the water extracts resulted in a total lack of inhibitory activity, suggesting that zerumbone is a primary bioactive constituent contributing to these anti-inflammatory effects. Furthermore, the steaming process led to a noticeable decrease in zerumbone content in ZTSH and ZTSE extracts compared to their non-steamed counterparts (ZTNH and ZTNE), This reduction corresponded with an increase in the IC50 values for both NO and IL-6 inhibition, thereby supporting the observation that steaming negatively impacts the bioactive content and efficacy of the extracts.
Interestingly, this outcome contrasts with traditional Thai practices, which typically involve steaming the rhizomes of Zingiberaceae plants prior to medicinal preparations. While a previous study reported that the ethanol extract of steamed ginger contained higher levels of 6-gingerol compared to the non-steamed extract [16], our current results suggest that zerumbone content in ZT rhizomes is particularly sensitive to thermal processing. Consequently, to preserve this bioactive constituent, minimal or no heat treatment may be preferable.
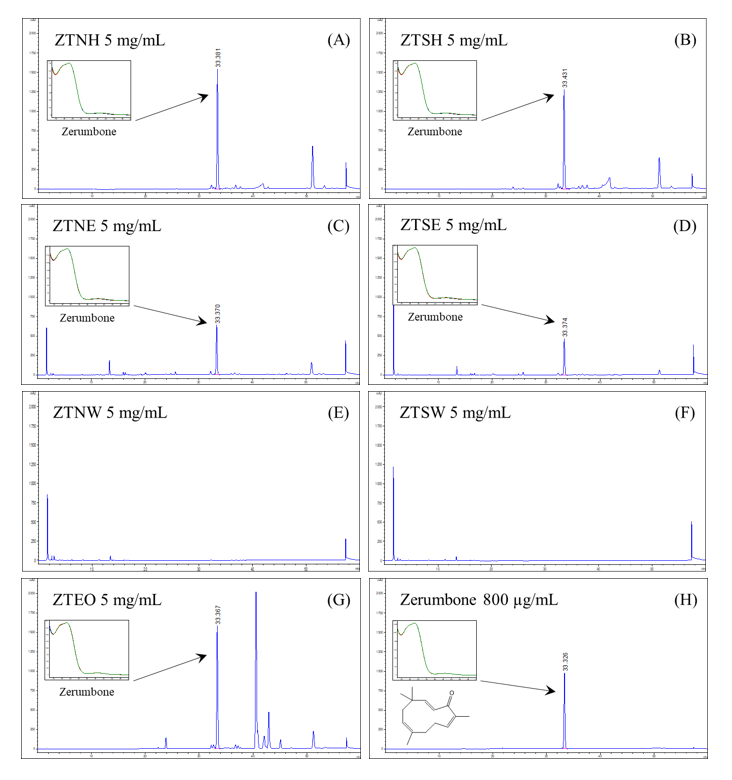
Figure 3 HPLC chromatograms of (A) ZTNH, (B) ZTSH, (C) ZTNE, (D) ZTSE, (E) ZTNW, (F) ZTSW, (G) ZTEO, and (H) standard zerumbone, at a specific UV wavelength of 215 nm.
Table 5 The amount of zerumbone in ZT extracts and essential oil.
Sample |
Zerumbone content (%w/w of extract) |
|||
N1 |
N2 |
N3 |
Mean ± SD |
|
ZTNH |
22.66 |
21.78 |
22.10 |
22.18 ± 0.44 a,* |
ZTSH |
19.93 |
19.65 |
19.38 |
19.65 ± 0.27 b,* |
ZTNE |
9.25 |
8.93 |
9.61 |
9.27 ± 0.34 c,* |
ZTSE |
7.28 |
7.39 |
7.35 |
7.34 ± 0.06 d,* |
ZTNW |
ND |
ND |
ND |
ND |
ZTSW |
ND |
ND |
ND |
ND |
ZTEO |
23.77 |
24.19 |
24.11 |
24.02 ± 0.22 |
Data are expressed as Mean ± SD (n = 3 independent experiments). Values followed by different superscript letters (a-d) indicate significant differences (p < 0.05) based on 2-Way ANOVA (solvent × steaming interaction) followed by Tukey’s post-hoc test. Independent samples t-tests were used to compare all individual ZT extracts with ZTEO (*, p < 0.05).
Conclusions
This study provides a comprehensive evaluation of the impact of steaming on the anti-inflammatory activity and chemical composition of ZT rhizome extracts and essential oil. The results demonstrate that non-steamed samples and essential oil possess superior anti-inflammatory properties, particularly through the inhibition of NO and IL-6 production, with zerumbone identified as a primary bioactive constituent. Steaming was found to reduce both the biological activity and zerumbone content, which may be attributed to thermal degradation or volatilization. GC-MS and HPLC analyses revealed significant differences in chemical compositions between steamed and non-steamed samples, highlighting the influence of steaming pretreatment. Collectively, these findings indicate that non-steamed ZT rhizomes are preferable for preserving anti-inflammatory potency and key phytochemical constituents.
Acknowledgements
This study was supported by Thammasat University Research Unit in Applied Thai Traditional Medicine and Herbal Innovation and a research grant from the Faculty of Medicine, Thammasat University, Thailand Contract No. TP. 2-07/2567. The authors also would like to acknowledge Michael Jan Everts, from the Clinical Research Center, Faculty of Medicine, Thammasat University, Thailand for English editorial assistance.
Declaration of Generative AI in Scientific Writing
The authors acknowledge the use of generative AI tools (e.g., QuillBot and Grammarly) in the preparation of this manuscript, specifically for language editing and grammar correction. No content generation or data interpretation was performed by AI. The authors take full responsibility for the content and conclusions of this work.
CRediT Author Statement
Patcharaporn Muanrit: Conceptualization, Methodology, Data curation, Formal analysis, Investigation, Validation, and Writing – original draft. Sumalee Panthong: Methodology, Resources, Data curation, Formal analysis, Investigation, Validation, Visualization and Supervision. Saovapak Poomirat: Resources, Data curation, Formal analysis, Investigation, Validation, Visualization and Supervision. Ubonwan Saesiw: Resources, Data curation, Formal analysis, Investigation, Validation, and Visualization. Intouch Sakpakdeejaroen: Conceptualization, Methodology, Data curation, Formal analysis, Investigation, Validation, Visualization, Project administration, Resources, Supervision, and Writing – review & editing.
References
VP Chavda, J Feehan and V Apostolopoulos. Inflammation: The cause of all diseases. Cells 2024; 13(22), 1906.
CLR Soares, P Wilairatana, LR Silva, PS Moreira, NMMV Barbosa, PR da Silva, HDM Coutinho, IRA de Meneze and CFB Felipe. Biochemical aspects of the inflammatory process: A narrative review. Biomedicine & Pharmacotherapy 2023; 168, 115764.
JL Wautier and MP Wautier. Pro- and anti-inflammatory prostaglandins and cytokines in humans: A mini review. International Journal of Molecular Sciences 2023; 24(11), 9647.
R Sohail, M Mathew, KK Patel, SA Reddy, Z Haider, M Naria, A Habib, ZU Abdin, WR Chaudhry and A Akbar. Effects of non-steroidal anti-inflammatory drugs (NSAIDs) and gastroprotective NSAIDs on the gastrointestinal tract: A narrative review. Curēus 2023; 15(4), e37080.
A Wongpia, P Kongsuwan and T Luangsuphabool. Chemical profiling of essential oils of Zingiber ottensii Valeton collected from Yala province, southern Thailand. AgriScience and Society Journal 2025; 1(1), 40-49.
W Thitinarongwate, W Nimlamool, P Khonsung, R Mektrirat and P Kunanusorn. Anti-inflammatory activity of essential oil from Zingiber ottensii Valeton in animal models. Molecules 2022; 27(13), 4260.
P Panyajai, F Chueahongthong, N Viriyaadhammaa, W Nirachonkul, S Tima, S Chiampanichayakul, S Anuchapreeda and S Okonogi. Anticancer activity of Zingiber ottensii essential oil and its nanoformulations. PLoS One 2022; 17(1), e0262335.
TV Chen, T Boonma and NTT Hien. An overview of the chemical compositions and biological activities of essential oils from selected Zingiber species (Zingiberaceae). Natural Product Communications 2025; 20(3), 1934578X251329422.
C Chantaranothai, T Palaga, A Karnchanatat and P Sangvanich. Inhibition of nitric oxide production in the macrophage-like RAW 264.7 cell line by protein from the rhizomes of Zingiberaceae plants. Preparative Biochemistry & Biotechnology 2013; 43(1), 60-78.
A Karnchanatat, N Tiengburanatam, A Boonmee, S Puthong and P Sangvanich. Zingipain, A cysteine protease from Zingiber ottensii Valeton rhizomes with antiproliferative activities against fungi and human malignant cell lines. Preparative Biochemistry & Biotechnology 2011; 41(2), 138-153.
J Ruttanapattanakul, N Wikan, K Chinda, T Jearanaikulvanich, N Krisanuruks, M Muangcha, S Okonogi, S Potikanond and W Nimlamool. Essential oil from Zingiber ottensii induces human cervical cancer cell apoptosis and inhibits MAPK and PI3K/AKT signaling cascades. Plants 2021; 10(7), 1419.
F Tunnisa, D Nur Faridah, A Afriyanti, D Rosalina, MA Syabana, N Darmawan and ND Yuliana. Antioxidant and antidiabetic compounds identification in several Indonesian underutilized Zingiberaceae spices using SPME-GC/MS-based volatilomics and in silico methods. Food Chemistry: X 2022; 14, 100285.
K Akiyama, H Kikuzaki, T Aoki, A Okuda, NH Lajis and N Nakatani. Terpenoids and a diarylheptanoid from Zingiber ottensii. Journal of Natural Products 2006; 69(11), 1637-1640.
W Thitinarongwate, R Mektrirat, W Nimlamool, P Khonsung, S Pikulkaew, S Okonogi and P Kunanusorn. Phytochemical and safety evaluations of Zingiber ottensii Valeton essential oil in zebrafish embryos and rats. Toxics 2021; 9(5), 102.
N Pimrat, S Panthong and A Itharat. Effect of steaming process on chemical content and biological activity of Curcuma zedoaria extract. Asian Medical Journal and Alternative Medicine 2021; 21(1), 45-54.
N Roekruangrit, N Jaiarree, A Itharat, A Wanichsetakul, P Wanichsetakul and W Pipatrattanaseree. Comparative study on antioxidant activities and phytochemical components of steamed and non-steamed ginger extracts. Asian Medical Journal and Alternative Medicine 2018; 18(4), 528-536.
N Roekruangrit, N Jaiarree, A Itharat, P Wanichsetakul, S Panthong, S Makchuchit and S Poomirat. Comparative study on biological activities of steamed and non - steamed ginger extracts. Science & Technology Asia 2019; 24(4), 94-101.
P Muanrit, S Poomirat and I Sakpakdeejaroen. Validation of HPLC method for quantitative determination of zerumbone in the rhizome of Zingiber ottensii Valeton. Talanta Open 2025; 11, 100405.
N Jaiarree, N Roekruangrit, A Itharat, P Wanichsetakul and S Poomirat. The efficacy of steamed ginger extract versus placebo for pain relief at the perineum and uterus in first normal postpartum women. Journal of the Medical Association of Thailand 2023; 106(4), 334-340
R Sriyom, A Itharat, O Prajuabjinda, P Thongdeeying, S Ruangnoo, S Makchuchit, P Kuropakornpong, K Namphonsaen, P Monkanna and NM Davies. Anti-inflammatory and cytotoxic effects of Jatropha podagrica extracts on skin cancer. Research in Pharmaceutical Sciences 2025; 20(4), 498-510.
T Kitnithiprapha, S Panthong, I Sakpakdeejaroen and S Kondo. Anti-Inflammatory and antimicrobial effects of herbal formulation called Apo-Taat using extended-spectrum ß-lactamase-producing Escherichia coli isolates. The Scientific World Journal 2025; 2025(1), 6151640.
RR Pratama, RA Sari, I Sholikhah, H Mansor, HI Chang, Sukardiman and R Widyowati. Inhibition of nitric oxide production in RAW 264.7 cells and cytokines IL-1β in osteoarthritis rat models of 70% ethanol extract of Arcangelisia flava (L.) merr stems. Heliyon 2024; 10(15), e35730.
K Sukkasem, A Itharat, K Thisayakorn, N Tangsuphoom, S Panthong, S Makchuchit, J Inprasit, N Prommee, W Khoenok, K Sriyam, D Pahusee, A Tasanarong, B Ooraikul and NM Davies. Exploring in vitro and in vivo anti-inflammatory activities of the Thai traditional remedy Kheaw-Hom and its bioactive compound, ethyl p-methoxycinnamate, and ethnopharmacological analysis. Journal of Ethnopharmacology 2024; 319, 117131.
ME Kim and JS Lee. Advances in the regulation of inflammatory mediators in nitric oxide synthase: Implications for disease modulation and therapeutic approaches. International Journal of Molecular Sciences 2025; 26(3), 1204.
HM AbuZahra, P Rajendran and MB Ismail. Zerumbone exhibit protective effect against zearalenone induced toxicity via ameliorating inflammation and oxidative stress induced apoptosis. Antioxidants 2021; 10(10), 1593.
CC Su, SC Wang, IC Chen, FY Chiu, PL Liu, CH Huang, KH Huang, SH Fang, WC Cheng, SP Huang, HC Yeh, CC Liu, PY Lee, MY Huang and CY Li. Zerumbone suppresses the LPS-induced inflammatory response and represses activation of the NLRP3 inflammasome in macrophages. Frontiers in Pharmacology 2021; 12, 652860.
J Zagórska, L Czernicka-Boś, W Kukula-Koch, R Szalak and W Koch. Impact of thermal processing on the composition of secondary metabolites of ginger rhizome-a review. Foods 2022; 11(21), 3484.
J Zagórska, L Czernicka-Boś, W Kukula-Koch, K Iłowiecka and W Koch. Impact of thermal processing on the selected biological activities of ginger rhizome-a review. Molecules 2023; 28(1), 412.
HT Nguyen, S Wu, T Ootawa, HC Nguyen, HT Tran, P Pothinuch, HTT Pham, ATH Do, HT Hoang, MZ Islam, A Miyamoto and HTT Nguyen. Effects of roasting conditions on antibacterial properties of Vietnamese turmeric (Curcuma longa) rhizomes. Molecules 2023; 28(21),7242.
MY Song, DY Lee, SY Park, SA Seo, JS Hwang, SH Heo and EH Kim. Steamed ginger extract exerts anti-inflammatory effects in Helicobacter pylori-infected gastric epithelial cells through inhibition of NF-κB. Journal of Cancer Prevention 2021; 26(4), 289-297.
NT Hien, DT Cuc, NTT Thuy, H Hiep, VT Huyen, DTT Ai and NX Nhiem. Labdane-type diterpenoids and sesquiterpenes from Curcuma aromatica and their nitric oxide inhibitory activity in lipopolysaccharide-stimulated RAW264.7 macrophages. Journal of Asian Natural Products Research 2024; 26(3), 387-393.
MD Ibáñez, NM Sánchez-Ballester and MA Blázquez. Healthy zerumbone: From natural sources to strategies to improve its bioavailability and oral administration. Plants 2022; 12(1), 5.