Trends
Sci.
2026;
23(8):
12658
Effect of Acute Toxicity of 2,4-D Dimethylammonium on Rice Field Crab (Esanthelphusa dugasti): Toxicity Testing, Hemocyte Abnormality, Neurotoxicity, and Alteration of Tissue
Chutima Thanomsit1, Sugunya Kumla2, Sawipa Ruttanakorn3, Witchuda Prasatkaew4, Panomsak Meemon5,6, Jakkaphun Nanuam7 and Phochit Nanthanawat7,*
1Department of Fisheries, Faculty of Agriculture and Technology,
Rajamangala University of Technology Isan Surin Campus, Surin 32000, Thailand
2Department of Fisheries, Faculty of Natural Resources,
Rajamangala University of Technology Isan Sakon Nakhon Campus, Sakon Nakhon 47160, Thailand
3Department of Fisheries Technology, Faculty of Agricultural Technology, Kalasin University,
Kalasin 46000, Thailand
4Faculty of Science and Technology, Dhonburi Rajabhat University, Samutprakan 10540, Thailand
5School of Physics, Institute of Science, Suranaree University of Technology, Nakhon Ratchasima 30000, Thailand
6Center of Excellence in Advanced Functional Materials, Suranaree University of Technology,
Nakhon Ratchasima 30000, Thailand
7Office of Educational Affairs, Faculty of Science, Burapha University, Chonburi 20131, Thailand
(*Corresponding author’s e-mail: [email protected])
Received: 14 November 2025, Revised: 10 January 2026, Accepted: 17 January 2026, Published: 15 March 2026
Abstract
This study investigated the acute and sub-lethal toxicity of 2,4-D dimethylammonium on the rice field crab (Esanthelphusa dugasti), an ecologically important crustacean in Southeast Asian rice-field ecosystems. Crabs were exposed to 0 - 3 mg/L of 2,4-D dimethylammonium for up to 96 h, and toxicity was assessed using mortality, hematological parameters, cholinesterase activity, protein expression, histopathology, immunohistochemistry, and Optical Coherence Tomography (OCT). Mortality increased in a concentration- and time-dependent manner, with the median lethal concentration (LC50) values decreasing from 4.67 mg/L at 24 h to 2.98 mg/L at 96 h. Significant reductions in total hemocyte counts and marked hemocyte deformities were observed at multiple exposure levels (p < 0.05). Neurotoxicity was evidenced by progressive inhibition of acetylcholinesterase (AChE) and butyrylcholinesterase (BChE), with AChE activity declining from 149.45 ± 0.95 nmol/min/mg protein in controls to 119.66 ± 2.02 nmol/min/mg protein at 3 mg/L after 96 h, accompanied by reduced AChE expression. Histopathological analysis revealed concentration-dependent damage in gill and hepatopancreas tissues, including vacuolation, lumen distension, lamellar collapse, and hemocyte infiltration (p < 0.0001). Notably, OCT enabled non-destructive, depth-resolved visualization of internal tissue disruption, corroborating histological findings and providing three-dimensional insight into organ-level pathology. Unlike previous studies on 2,4-D dimethylammonium toxicity in crabs that relied on conventional endpoints, this study demonstrates the novel application of OCT as a complementary imaging tool linking molecular neurotoxicity with structural tissue damage. These findings highlight the ecological risk of 2,4-D dimethylammonium to rice-field crabs and support the use of OCT in advanced aquatic ecotoxicological assessment.
Keywords: Acetylcholinesterase, Aquatic toxicology, Hemocyte morphology, Optical coherence tomography, Rice field crab, 2,4-D dimethylammonium
Introduction
2,4-Dichlorophenoxyacetic acid is a phenoxy acetic acid herbicide widely used for the selective control of broadleaf weeds [1]. Among its various formulations, the dimethylammonium salt is particularly prevalent in agriculture due to its high solubility and effectiveness. This brown liquid formulation is commonly applied post-emergence in crops such as rice, wheat, and maize. Its extensive use, especially in rice paddies, has led to frequent detection in water and soil, raising concerns over its ecological and toxicological impacts on non-target organisms.
The environmental behavior of 2,4-D dimethylammonium is shaped by its physicochemical properties, namely, its high-water solubility and ability to persist under certain conditions. While generally biodegradable, 2,4-D dimethylammonium has a half-life of approximately 10 days in soils and even shorter in water, though residues may persist longer under dry or cold conditions [2]. In aquatic environments, it exists predominantly as a free anion, especially from amino salt formulations, and undergoes rapid hydrolysis. However, this process slows at higher pH levels, increasing its environmental persistence and mobility. Contamination of ponds and streams can occur through direct application, surface runoff, or atmospheric deposition [2,3]. Surveys in Malaysia and Thailand have confirmed measurable levels of 2,4-D dimethylammonium in paddy soils and water bodies, demonstrating its widespread environmental presence [4].
Toxicologically, 2,4-D dimethylammonium functions as a synthetic auxin, mimicking plant growth hormones and disrupting cellular regulation. While this mechanism targets plants, emerging evidence suggests it may also exert endocrine-disrupting effects in non-target animals [5]. In crustaceans, endocrine functions such as molting and reproduction are regulated by hormonal pathways like ecdysteroids and crustacean hyperglycemic hormone (CHH). Although these systems differ from those in plants, concerns about indirect or cross-reactive toxicity have prompted broader evaluations of 2,4-D dimethylammonium’s impacts beyond acute toxicity, particularly in aquatic invertebrates.
Research on the impacts of 2,4-D dimethylammonium on aquatic species has expanded in recent years. Khanchanasal et al. [5] reported that exposure in adult Nile tilapia resulted in increased mortality, behavioral abnormalities, external morphological changes, impaired acetylcholinesterase (AChE) activity, and tissue damage in the liver, gills, stomach, and intestines. Several studies have demonstrated that herbicides inhibit AChE, particularly under prolonged exposure, whereas short-term exposure may stimulate compensatory increases in AChE synthesis, supporting its use as a short-term bioindicator [6]. Thanomsit et al. [6] also documented sub-lethal effects of 2,4-D dimethylammonium on AChE expression across multiple tissues including the brain, gills, muscles, and plasma along with associated histological and morphological alterations.
For crustacean-related studies reported in Thailand, Kumla et al. [7] indicated that the widespread use of herbicides such as 2,4-D dimethylammonium poses significant risks to aquatic ecosystems. Their study examined the toxicological effects of 2,4-D dimethylammonium on juvenile giant freshwater prawns (Macrobrachium rosenbergii), focusing on mortality, immune responses, antioxidant enzyme activities, neurotoxicity, and histological alterations. Acute toxicity tests demonstrated dose- and time-dependent mortality, with LC₅₀ values decreasing from 24 to 96 h. Sub-lethal exposure resulted in significant reductions in total hemocyte count (THC) and induced morphological abnormalities in hemocytes. Moreover, optical coherence tomography (OCT) provided detailed visualization of structural changes in the head, compound eyes, and abdomen.
In addition, adverse effects of 2,4-D dimethylammonium on rice field crabs (Somanniathelphusa bangkokensis) have also been documented, including morphological, biochemical, and histological alterations [8]. Despite these advances, existing studies on 2,4-D dimethylammonium toxicity in crustaceans have largely relied on endpoint-specific or destructive analyses and have focused primarily on a limited number of species. Consequently, there remains a critical gap in understanding how neurotoxicity and immunotoxicity induced by 2,4-D dimethylammonium translate into depth-resolved, three-dimensional structural damage in non-target rice field crabs. In particular, no previous study has integrated hematological, cholinesterase-based biomarkers with OCT to directly link molecular and cellular dysfunction with organ-level pathology in Esanthelphusa dugasti. Addressing this gap is essential for improving mechanistic interpretation and ecological relevance of herbicide toxicity assessments in rice-field ecosystems.
The common rice field crab, E. dugasti [9] is a semi-terrestrial crustacean endemic to Southeast Asia, especially Thailand. It inhabits rice paddies, adjacent fields, and burrows, playing a vital role in ecosystem nutrient cycling and serving as a low-cost protein source for local communities [10]. Its close association with agricultural landscapes, however, renders it vulnerable to pesticide exposure. With its dual aquatic-terrestrial lifestyle and omnivorous feeding behavior, E. dugasti faces multiple exposure routes, including ingestion of contaminated food or sediment, dermal absorption from treated soils, and direct contact with pesticide-laden water.
Additionally, the species is occasionally considered a pest by farmers, which may result in intentional chemical exposure. These socio-ecological dynamics heighten the likelihood of contamination and pose food safety risks to human consumers. Despite 2,4-D dimethylammonium’s widespread use and its relatively low bioaccumulation potential, little is known about its sub-lethal or chronic effects on E. dugasti or the associated public health implications [9].
Based on existing evidence of herbicide-induced neurotoxicity and immunotoxicity in aquatic invertebrates, this study was designed to test the hypothesis that increasing concentrations and exposure duration of 2,4-D dimethylammonium would result in progressively greater toxic effects in E. dugasti. Specifically, we hypothesized that exposure would (i) produce concentration- and time-dependent mortality, (ii) suppress acetylcholinesterase and butyrylcholinesterase activity and expression as indicators of neurotoxicity, (iii) induce alterations in total hemocyte count and hemocyte morphology reflecting immunotoxic effects, and (iv) cause structural damage in gill and hepatopancreas tissues that can be visualized using OCT. By explicitly testing these hypotheses, the present study aims to link molecular, cellular, and tissue-level toxicity within an integrated mechanistic framework and to demonstrate the potential of OCT as a complementary tool for aquatic toxicological assessment.
Materials and methods
Ethics statement
All animal experiments were conducted in accordance with the Ethical Principles and Guidelines for the Use of Animals in Scientific Research, as established by the National Research Council of Thailand. This study was approved under animal use license number Ul-03405-2559. All procedures involving animals were performed following the regulations of the Faculty of Agriculture and Technology and were approved by the Committee for Animal Biological Experimentation at Rajamangala University of Technology, Isan.
Preparation of test animals
A total of 200 adult male rice field crabs (E. dugasti) were collected from a private farm in Mueang District, Sakon Nakhon Province, Thailand. The crabs used in the experiment had an average weight range of 17.2 ± 1.5 g. Prior to experimentation, the animals were acclimated for 30 days in a concrete tank (60×60×100 cm3) under continuous aeration. Water quality was monitored daily throughout the acclimation and experimental periods to ensure stable environmental conditions. Key parameters included temperature (24 - 28 °C), pH (8.6 ± 0.8), dissolved oxygen (6.5 ± 0.7 mg/L), and total ammonia concentration (<0.2 mg/L).
During the acclimation period, the crabs were fed a commercial pellet diet formulated for freshwater prawns (CP 9092 L; CPF Global Food Solution Public Company Ltd., Thailand), containing 36% crude protein. Feeding was conducted twice daily (morning and evening) at a ration of approximately 3% of body weight per day. Feeding was suspended 24 h prior to the start of the exposure experiments to minimize potential effects of feeding on water quality and physiological variability.
Toxicity testing
To assess the toxicity of 2,4-D dimethylammonium, rice field crabs were initially exposed to a broad concentration range of 0, 1, 10, 100, and 1000 mg/L. Each treatment was conducted in triplicate, with 20 crabs per replicate. Based on the preliminary results, a narrower concentration range (0, 0.75, 1.5, 2.25, and 3 mg/L) was subsequently tested over exposure periods of 24, 48, 72, and 96 h, also in triplicate with 20 crabs per group. The median lethal concentration (LC₅₀) was calculated using probit analysis based on cumulative mortality rates, in accordance with OECD guidelines [11].
The selected concentration ranges were designed to reflect environmentally relevant exposure scenarios in rice-field ecosystems. Although typical background concentrations of 2,4-D dimethylammonium in surface waters are generally lower, short-term pulse exposures may occur in shallow rice paddies following herbicide application, rainfall-driven runoff, irrigation discharge, or rapid water exchange events. Therefore, the preliminary wide-range exposure was used for range finding, while the narrower concentration range (0 - 3 mg/L, expressed as active ingredient) was selected to establish the No Observed Effect Concentration (NOEC) and Lowest Observed Effect Concentration (LOEC), and LC₅₀ values under acute exposure conditions and to represent realistic worst-case pulse scenarios relevant to rice-field crabs.
Hemolymph collection and THC
The base of the fourth and fifth walking legs was cleaned with 70% ethanol prior to hemolymph extraction using a 21-gauge needle. Hemolymph was collected into tubes containing 10% trisodium citrate (1:1 ratio) and centrifuged at 3,000×g for 10 - 15 min. The supernatant was used to assess total hemocyte counts using a hemocytometer. Samples were stained with Wright-Giemsa stain (15 - 20 min), rinsed with distilled water, and observed under a light microscope at 40x magnification. Differential hemocyte counts were divided into different subtypes based on morphology, especially size and nuclear-to-cytoplasmic ratio performed using differential interference contrast microscopy, classifying subtypes as: (1) large granule hemocytes (granulocytes) with high-density granules, (2) small granule hemocytes (semi-granulocytes) with low-density granules, and (3) small hyalinocytes (Type 1 - 4) with no cytoplasmic granules and irregular change. Further subclassification followed [12].
Protein extraction and quantification
Gill tissues from rice field crabs one individual per treatment, with three replicates for each experimental group were homogenized in 0.02 M Tris-HCl buffer (pH 7.2) containing phenylmethylsulfonyl fluoride (PMSF), at a ratio of 1 g of tissue to 1 mL of buffer. The homogenates were centrifuged at 4 °C at 4,500 rpm for 60 min, and the resulting supernatant was used for protein analysis via SDS-PAGE and for AChE expression analysis using Western blot.
Protein concentrations in the gill tissue samples from both the control and 2,4-D dimethylammonium-exposed groups were measured using the Bradford method. Protein levels were compared to a bovine serum albumin (BSA) standard solution. Absorbance was measured at 595 nm, and total protein concentrations were calculated by comparing the absorbance readings to those of the BSA standard, following [13].
AChE and butyrylcholinesterase (BChE) activity
Following a modified protocol based on [14], the activities of AChE and BChE were evaluated in gill tissues of rice field crabs exposed to 2,4-D dimethylammonium at concentrations of 0, 0.75, 1.5, 2.25, and 3 mg/L for 24, 48, 72, and 96 h (modified from [15]. Tissues from three replicates in each treatment, including both control and exposed groups, were homogenized on ice in 0.02 M phosphate buffer (pH 7.4), and the homogenates were centrifuged at 5,000×g for 60 min at 4 °C. The resulting supernatant was stored at –4°C for subsequent analysis. A colorimetric assay was used to determine enzyme activity at room temperature. AChE activity was measured based on the reaction between acetylthiocholine and dithiobisnitrobenzoate, as described by Ellman et al. [16], with absorbance recorded at 405 nm over a 5-min period using a microplate reader. BChE activity was assessed using butyrylthiocholine as the substrate, following the method of Escartín and Porte [17]. Enzyme activities were expressed as nanomoles per minute per milligram of protein (nmol/min/mg protein).
SDS-PAGE and western blot for AChE expression
The supernatant from rice field crab gill tissue was mixed with 2×sample buffer (50% glycerol, 250 mM Tris-HCl buffer pH 6.8, 10% SDS, 0.5% bromophenol blue, and 0.5% mercaptoethanol) at a 1:1 ratio, then heated at 95 °C for 10 min and subsequently cooled on ice. Samples were loaded into SDS-PAGE gels (10% acrylamide for separating, 4% acrylamide for stacking) and electrophoresed at 120 V for 70 min. After electrophoresis, gels were stained with Coomassie Brilliant Blue for 2 h, destained with a methanol-acetic acid solution, and analyzed. For Western blotting, proteins were transferred onto a nitrocellulose membrane and incubated overnight with PAb-AChE (1:50). The membrane was then washed with 0.5% Blotto solution and incubated with GAR-HRP for 3 h followed by DAB staining (0.03% DAB, 0.006% H₂O₂, 0.05% CoCl₂) for 3 min. The blots were visualized under a microscope for color development [16].
Histological and immunohistochemical analysis
Rice field crabs from both the control and treated groups were randomly selected for tissue analysis, with one crab taken from each replicate, resulting in three crabs per treatment. The selected crabs were sacrificed on ice, and the hepatopancreas and gill tissues were fixed in 10% phosphate-buffered formalin at 4 °C. The samples were processed, trimmed, dehydrated, and embedded in paraffin. Paraffin blocks were sectioned at 6 µm using a microtome, mounted on glass slides, and baked at 45 °C to ensure proper adherence. The slides were stained with Hematoxylin and Eosin (H&E) for morphological analysis in both control and treated groups [8].
Specimens were examined under a light microscope, and a comprehensive histological assessment was performed following the slightly modified method of Bernet et al. [18] for evaluating aquatic pollution. For each crab, histological alterations in the hepatopancreas and gills were recorded, if present, across ten non-overlapping high-power fields (HPF) at 40x magnification. The frequency of each alteration was calculated using the following Eq. (1)
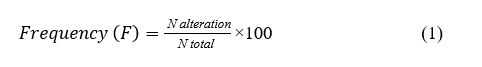
where N alteration is the number of occurrences of a specific alteration, and N total is the total number of 40x microscopic fields examined within the group (100).
The observed alterations were grouped into five reaction patterns (rp), each comprising several related alterations. Each alteration (alt) was scored (a) on a scale from 0 (absence) to 6 (diffuse) to reflect the extent of distribution and was assigned an importance factor (w) ranging from 1 (mild) to 3 (severe), indicating the level of severity. Organ indices (I org) for the hepatopancreas and gills were calculated using the following formula:
![]()
A higher organ index indicates a more severe pathological condition. The total index for each group of crabs was then calculated as:

The higher the total index, the more severe the pathological condition in the examined organs of that group.
For immunohistochemistry, slides were deparaffinized and rehydrated, then incubated with PAb-AChE for 3 h. After washing with PBS, the slides were incubated with GAR-HRP (1:1,000) for 3 h followed by staining with DAB substrate for 5 min. Finally, the slides were rinsed with distilled water and evaluated under a 40x magnification microscope (OLYMPUS CX31, Japan).
OCT analysis
OCT system used in this study was a custom-built, spectrometer-based frequency-domain OCT (FD-OCT) system, adapted from the designs of Thanomsit et al. [19] and Saetiew et al. [20]. For this study, the system was configured to acquire data at a rate of 20,000 depth scans (A-scans) per second, enabling real-time processing and display at 20 frames per sec. Each frame comprised 1,000 A-scans. A complete 3D dataset consisting of 1,000 cross-sectional (B-scan) images was acquired in approximately one minute, including data storage time. The system provided axial and transverse resolutions of approximately 10 µm and 15 µm, respectively, with a maximum imaging depth of 1 mm, limited by the inherent opacity of the sample.
Due to the size and opacity of the rice field crabs, in vivo OCT imaging was not feasible. Therefore, ex vivo imaging was performed. After exposure to 2,4-D dimethylammonium, the crabs were sacrificed, and the compound eyes, hepatopancreas, and gills were carefully dissected and placed in Petri dishes for imaging. Although in vivo imaging was not possible, OCT provided a key advantage in enabling non-destructive analysis of intact tissue without requiring sectioning or staining. This capability allowed real-time visualization of depth-resolved morphological changes, significantly reducing both the time and costs typically associated with traditional histological preparation.
Each 3D OCT dataset covered a region of interest (ROI) of up to 10×10 mm2 in the lateral plane and 1 - 2 mm in depth, depending on the specific tissue (hepatopancreas or gill). Data processing was performed using LabVIEW software (National Instruments, USA), and the resulting images were exported in JPEG format. Morphological changes were assessed using en-face image reconstruction in ImageJ (NIH, USA), and 3D volumetric rendering was conducted to enable detailed structural analysis of the affected tissues.
Statistical analysis
Data for total hemocyte count, percentage of abnormal hemocytes, protein concentration, and AChE and BChE activities were analyzed using two-way analysis of variance (ANOVA) to evaluate the effects of concentration and exposure time. Prior to ANOVA, data normality was assessed using the Shapiro-Wilk test, and homogeneity of variances was evaluated using Levene’s test. When necessary, data were log-transformed to meet ANOVA assumptions. Statistical significance was accepted at p < 0.05. When significant differences were detected, Duncan’s New Multiple Range Test was applied for post hoc comparison of treatment means. All statistical analyses were performed using the PROC ANOVA procedure in SAS® OnDemand for Academics.
Results and discussion
Cumulative mortality percentage and median lethal concentration (LC50)
The cumulative mortality of rice field crabs (E. dugasti) exposed to 2,4-D dimethylammonium at concentrations of 0, 0.75, 1.5, 2.25, and 3 mg/L for 24, 48, 72, and 96 h increased in a clear dose- and time-dependent manner. At 24 h, mortality rates were 0.00%, 8.33 ± 2.89%, 13.33 ± 2.89%, 18.33 ± 2.89%, and 21.67 ± 5.77%, respectively, and continued to rise at 48 h to 0.00%, 15.00 ± 0.00%, 16.67 ± 2.89%, 23.33 ± 2.89%, and 26.67 ± 5.77%. At 72 h, mortality rates were 0.00%, 18.33 ± 2.89%, 21.67 ± 2.89%, 25.00 ± 0.00%, and 28.33 ± 2.89%, and by 96 h, they reached 0.00%, 25.00 ± 0.00%, 33.33 ± 2.89%, 40.00 ± 5.00%, and 50.00 ± 5.00%, respectively. Statistical analysis confirmed significant differences between the control and exposure groups, particularly at concentrations ≥1.5 mg/L at each time point (p < 0.05), as illustrated in the radar plots (Figure 1). The LC₅₀ values calculated using probit analysis decreased with longer exposure, measuring 4.67, 4.34, 4.20, and 2.98 mg/L at 24, 48, 72, and 96 h, respectively, indicating increasing toxicity over time.
THC and abnormal hemocytes
THC in rice field crabs (E. dugasti) exposed to 2,4-D dimethylammonium at concentrations of 0.75, 1.5, 2.25, and 3 mg/L for 24, 48, 72, and 96 h exhibited a clear concentration-dependent decline. Statistically significant differences (p < 0.05) were observed between the control and treatment groups at multiple time points. At 24 h, THC was significantly reduced in the 1.5, 2.25, and 3 mg/L groups compared to the control. This trend continued at 48 and 72 h, with progressively lower hemocyte counts at higher concentrations. By 96 h, the effect was most pronounced, with the 3 mg/L group showing the lowest THC (1.34 ± 0.02×10⁶ cells/mL) relative to the control group (1.58 ± 0.02×10⁶ cells/mL), indicating cumulative immunotoxicity with prolonged exposure (Figure 2).
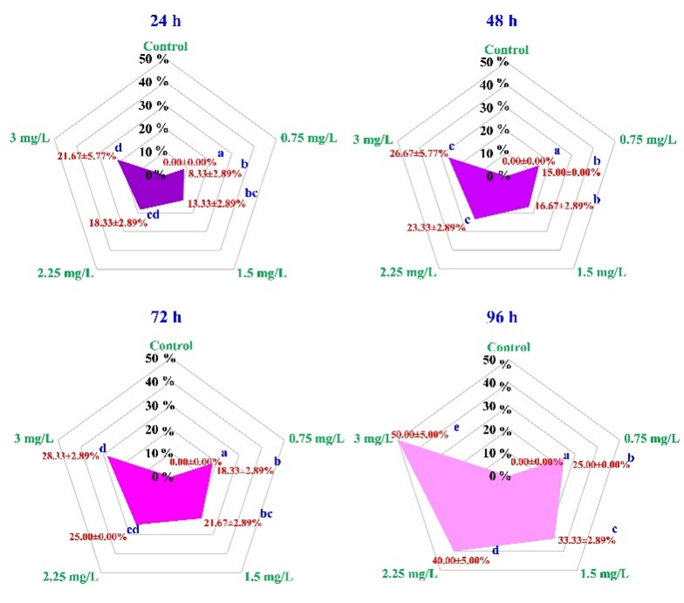
Figure 1 Cumulative mortality percentages of rice field crabs exposed to 2,4-D dimethylammonium at concentrations of 0 (control), 0.75, 1.5, 2.25, and 3 mg/L over 24, 48, 72, and 96 h. Mortality increased in a dose- and time-dependent manner. Different lowercase letters indicate significant differences between treatment groups at each time point (p < 0.05).
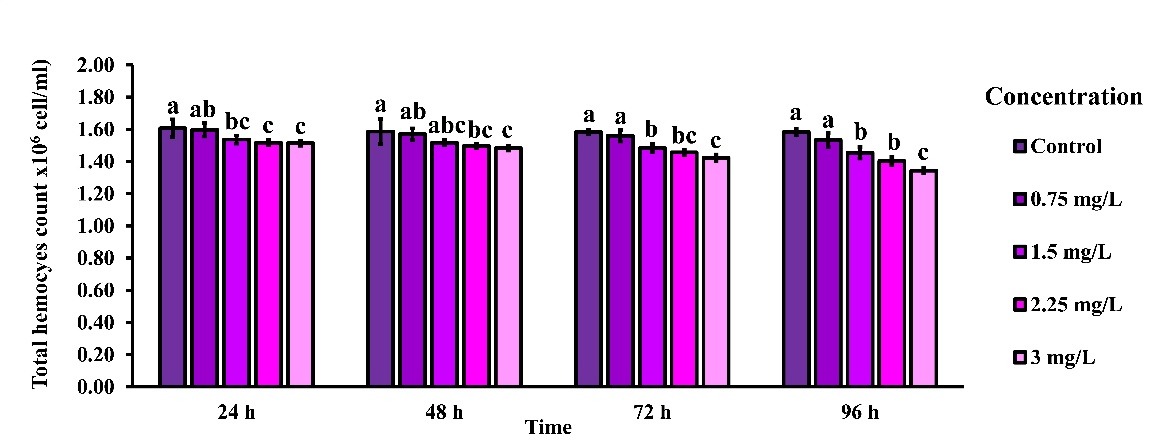
Figure 2 THC in rice field crabs exposed to 2,4-D dimethylammonium at concentrations of 0 (control), 0.75, 1.5, 2.25, and 3 mg/L for 24, 48, 72, and 96 h. THC decreased in a concentration- and time-dependent manner. Different lowercase letters indicate statistically significant differences between groups at each time point (p < 0.05).
Figure 3 presents representative images of hemocyte abnormalities in rice field crabs exposed to 2,4-D dimethylammonium at concentrations of 0, 0.75, 1.5, 2.25, and 3 mg/L. Six hemocyte types were examined: large granule hemocytes (LGH), small granule hemocytes (SGH), and hyalinocytes types 1 to 4. Exposure to increasing concentrations of the herbicide resulted in distinct morphological alterations. Abnormalities involving both the granular cells (red arrows) and the cytoplasm (blue arrows) were observed in LGHs, SGHs, and hyalinocyte type 2 (Figure. 3, columns 1 - 3). In contrast, hyalinocytes types 1 and 3 exhibited predominantly cytoplasmic abnormalities (Figure 3, columns 3 and 5, respectively), while hyalinocyte type 4 showed alterations primarily restricted to granular components (Figure 3, column 6).
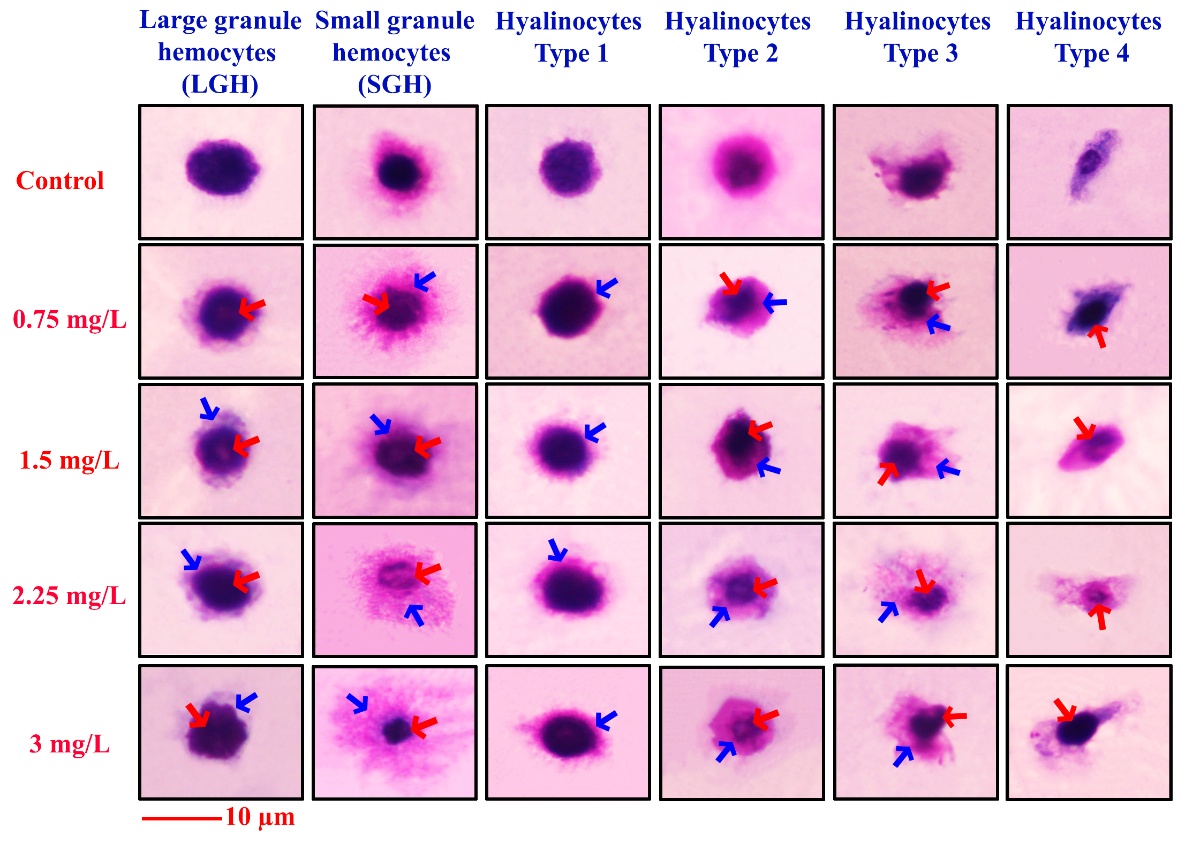
Figure 3 Morphology of six hemocyte types in rice field crabs exposed to 2,4-D dimethylammonium (0 - 3 mg/L). Dose-dependent abnormalities were observed, including cytoplasmic shrinkage (blue arrows) and granule disintegration (red arrows). Scale bar = 10 µm.
Hemocyte abnormalities in rice field crabs
Hemocyte abnormalities were assessed in rice field crabs exposed to 2,4-D dimethylammonium at various concentrations and exposure durations. At 24 h, significant abnormalities in large granule hemocytes were observed only at 3 mg/L (2.50 ± 0.71%, p < 0.05). For small granule hemocytes, significant abnormalities were detected at concentrations of 1.5, 2.25, and 3 mg/L, with the highest percentage (2.50 ± 0.71%) at 3 mg/L. Hyalinocytes types 1 to 4 also showed concentration-dependent abnormalities, with the highest level (3.00 ± 0.00%) recorded across all types at 3 mg/L.
At 48 h, all tested concentrations caused statistically significant abnormalities in all hemocyte types compared to the control group (p < 0.05). This trend continued at 72 h, where exposure to all concentration levels led to significant abnormalities. At the highest concentration (3 mg/L), the percentages of abnormalities were 6.50 ± 0.71% for large granule hemocytes, 6.00 ± 0.00% for small granule hemocytes, 7.00 ± 0.00% for hyalinocyte type 1, 6.00 ± 0.00% for type 2, and 7.00 ± 0.00% for types 3 and 4.
At the final time point of 96 h, significant abnormalities were observed at all concentrations and in all hemocyte types compared to the control (p < 0.05). The most pronounced abnormalities occurred at 3 mg/L, with values of 7.50 ± 0.71% for large granule hemocytes, 6.50 ± 0.71% for small granule hemocytes, 6.50 ± 0.71% for hyalinocyte type 1, 7.50 ± 0.71% for type 2, 7.50 ± 0.71% for type 3, and 9.00 ± 0.00% for type 4 (Figure 4).
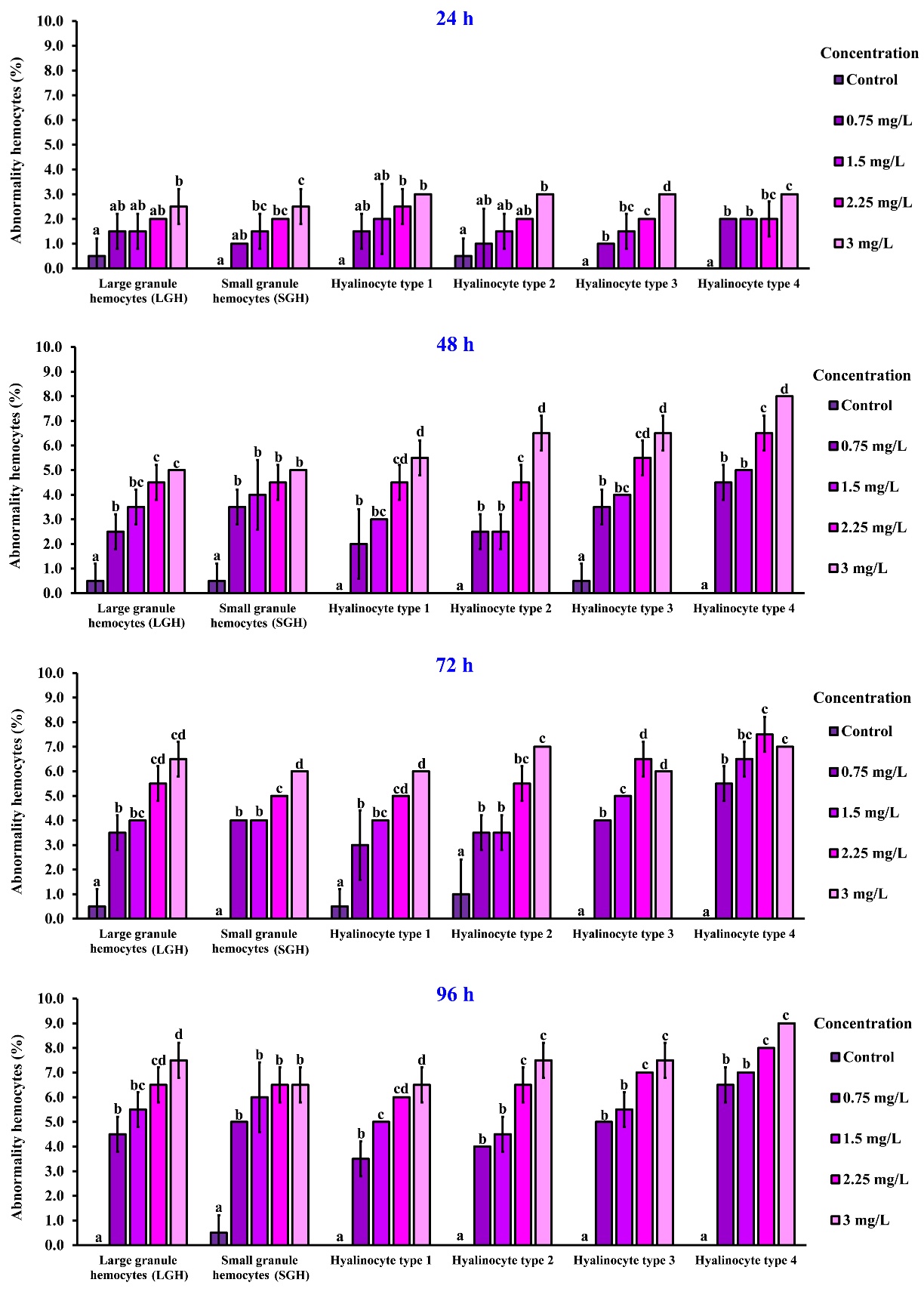 Figure
4
Hemocyte abnormality percentages in rice field crabs exposed to
2,4-D dimethylammonium (0 - 3 mg/L) for 24, 48, 72, and 96 h.
Abnormalities increased with both concentration and exposure time
across all hemocyte types. Different lowercase letters indicate
significant differences between groups (p
< 0.05).
Figure
4
Hemocyte abnormality percentages in rice field crabs exposed to
2,4-D dimethylammonium (0 - 3 mg/L) for 24, 48, 72, and 96 h.
Abnormalities increased with both concentration and exposure time
across all hemocyte types. Different lowercase letters indicate
significant differences between groups (p
< 0.05).
AChE and BChE activity in muscle
The activities of AChE and BChE in the muscle tissue of rice field crabs exposed to 2,4-D dimethylammonium at concentrations of 0.75, 1.5, 2.25, and 3 mg/L for 24, 48, 72, and 96 h, relative to the control group are shown in Figure 5 (top and bottom panels, respectively). AChE activity (Figure 5, top panel) declined progressively with increasing concentrations at all time points. At 24 h, AChE activity decreased from 149.45 ± 0.95 nmol/min/mg protein in the control group to 136.18 ± 1.49 at 3 mg/L, with further reductions observed at 48 h (149.29 ± 2.45 to 132.18 ± 1.64), 72 h (148.63 ± 1.97 to 126.67 ± 2.21), and 96 h (148.67 ± 0.64 to 119.66 ± 2.02). BChE activity (Figure 5, bottom panel) showed a similar concentration-dependent decrease, dropping from 44.68 ± 0.78 to 36.12 ± 0.16 at 24 h, 44.61 ± 0.53 to 32.55 ± 1.12 at 48 h, 44.45 ± 0.81 to 29.02 ± 1.41 at 72 h, and 44.90 ± 0.01 to 25.40 ± 1.18 at 96 h. These results indicate that both AChE and BChE are significantly inhibited by 2,4-D dimethylammonium in a dose- and time-dependent manner, suggesting neurotoxic effects on the exposed crabs.
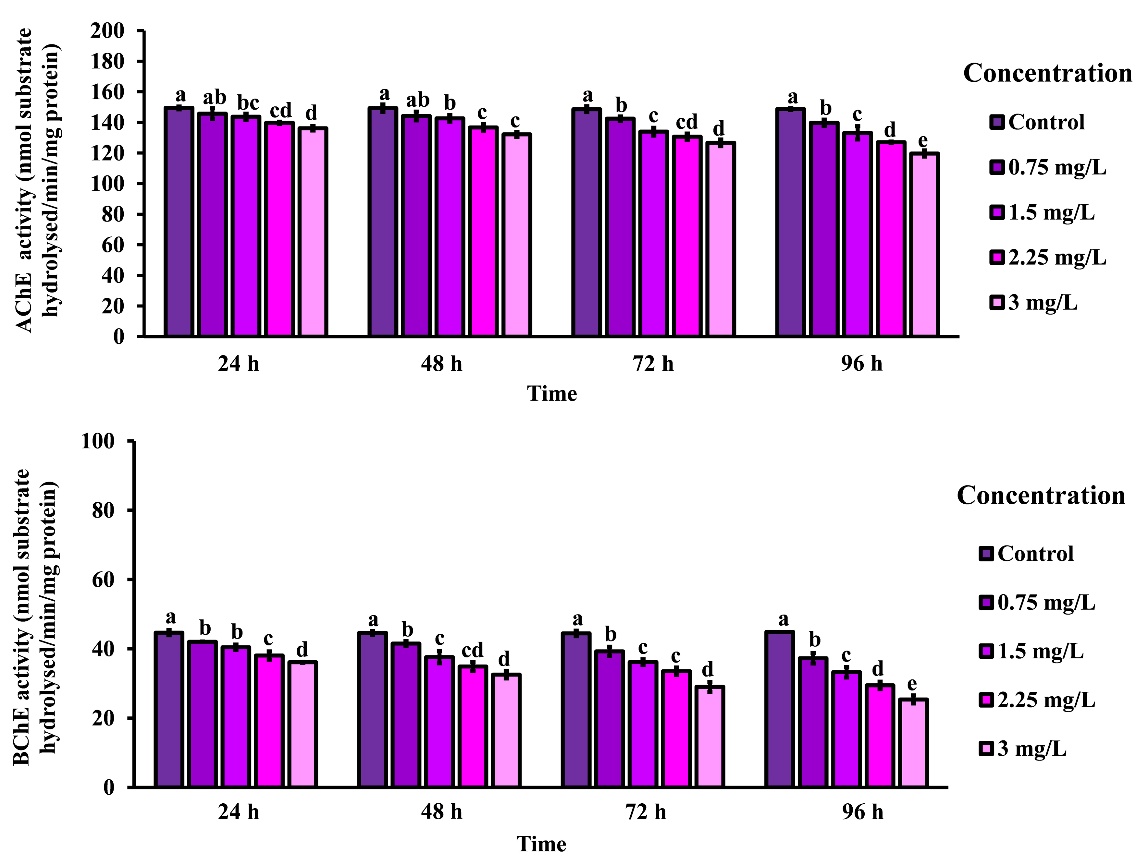
Figure 5 AChE and BChE activities in the muscle of rice field crabs exposed to 2,4-D dimethylammonium (0 - 3 mg/L) for 24, 48, 72, and 96 h. Both enzymes showed a significant, dose- and time-dependent decrease in activity. Different lowercase letters indicate significant differences between groups (p < 0.05).
Protein analysis and AChE expression
Figure 6 presents the protein profiles of gill tissues from rice field crabs exposed to 2,4-D dimethylammonium at concentrations of 0, 0.75, 1.5, 2.25, and 3 mg/L for 24, 48, 72, and 96 h, as analyzed by SDS-PAGE. Two prominent protein bands, corresponding to molecular weights of 73 and 71 kDa, were consistently observed. However, exposure to higher concentrations and longer durations resulted in a reduction in both band size and intensity, indicating potential protein degradation or expression suppression.
Western blot analysis revealed that AChE was expressed as a protein band with a molecular weight of 71 kDa. The intensity of the AChE band decreased progressively with increasing herbicide concentration and exposure time. Notably, AChE expression was undetectable at 72 h in the 3 mg/L group, and at 96 h, no AChE expression was observed in the 1.5, 2.25, and 3 mg/L groups.
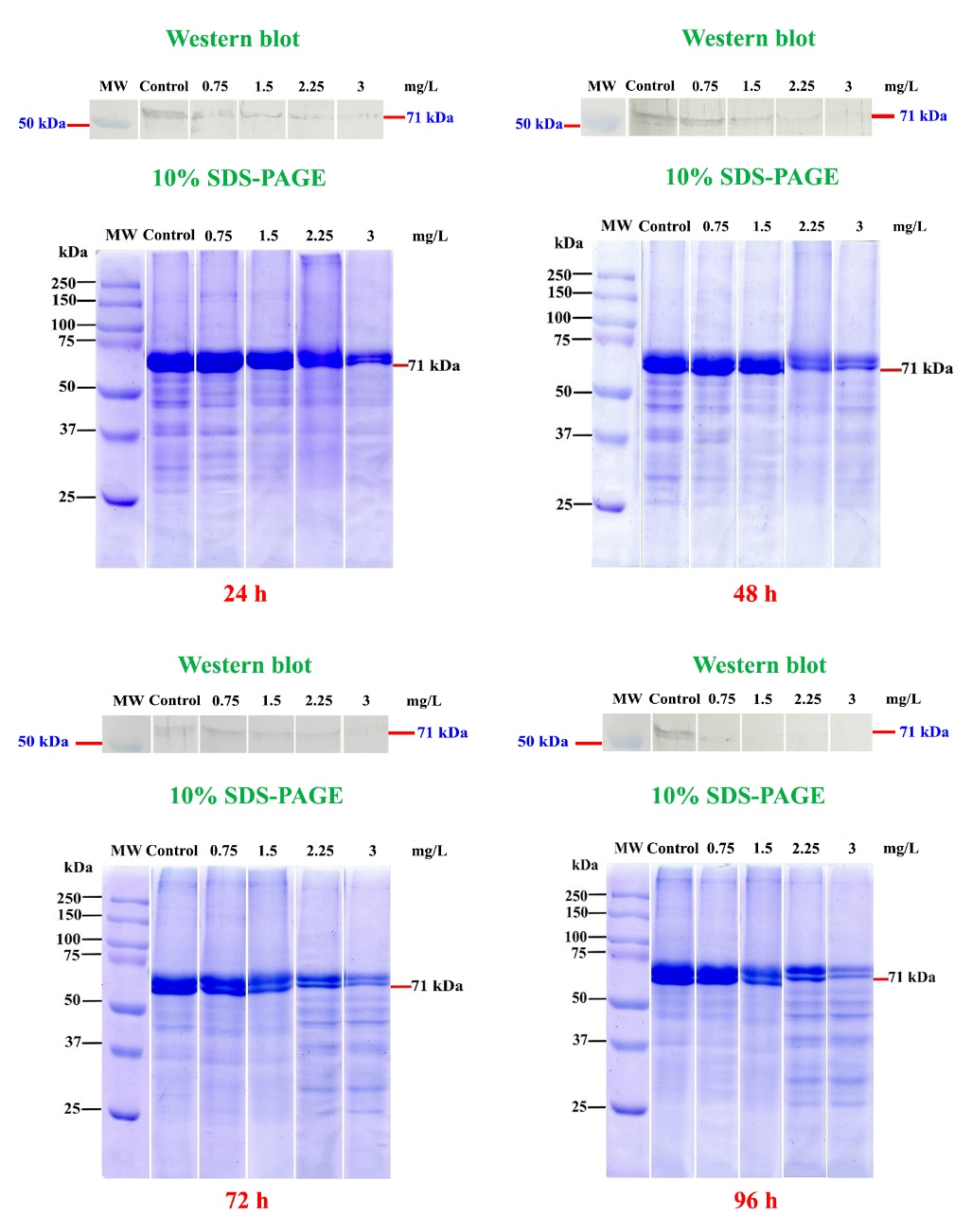
Figure 6 SDS-PAGE and Western blot analysis of gill tissue from rice field crabs exposed to 2,4-D dimethylammonium (0 - 3 mg/L) for 24 - 96 h. Protein bands near 71 kDa decreased with higher concentrations and longer exposure. AChE expression (71 kDa) was progressively reduced and absent at ≥1.5 mg/L after 96 h.
Histological alterations
Figure 7 illustrates representative histopathological changes in the hepatopancreas (upper panels) and gill (lower panels) of rice field crabs (Esanthelphusa dugasti) exposed to 2,4-D dimethylammonium at concentrations of 0, 0.75, 1.5, 2.25, and 3 mg/L for 24, 48, 72, and 96 h. Lesions became progressively more pronounced with increasing concentration and exposure duration. In the hepatopancreas, prominent alterations included lumen distension (DL) and vacuolation (V), particularly at concentrations ≥1.5 mg/L after 48 - 96 h. In gill tissues, structural damage such as lamellar collapse (LC), cuticle rupture (CR), tissue swelling (TS), and hemocyte infiltration (IH) intensified at higher concentrations and longer exposure times, indicating progressive tissue degeneration, especially at 2.25 - 3 mg/L after 72 - 96 h (Figure 7, gill panels).
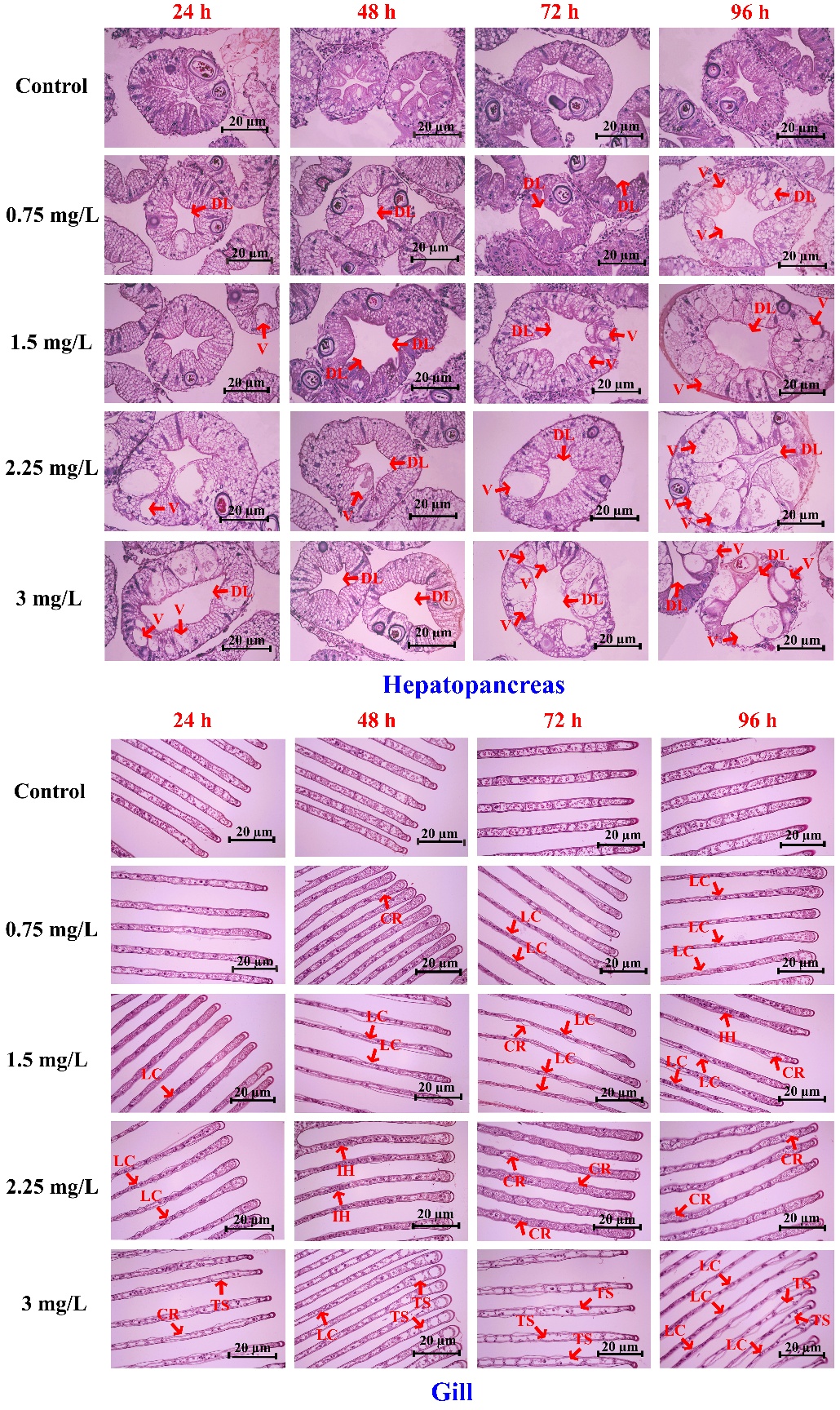
Figure 7 Histopathological alterations in the hepatopancreas (upper panels) and gill (lower panels) of Esanthelphusa dugasti following exposure to 2,4-D dimethylammonium at different concentrations (0 - 3 mg/L) and exposure durations (24 - 96 h). Rows represent herbicide concentration, and columns represent exposure time. Arrows indicate major pathological features, including lumen distension (DL), vacuolation (V), epithelial degeneration (DE), lamellar collapse (LC), and hemocyte infiltration (H). Scale bar = 20 µm.
Table 1 summarizes the frequency and severity of these alterations using organ index values derived from lesion frequency, extent, and severity. In the hepatopancreas, both lumen distension and vacuolation increased significantly at 2.25 and 3 mg/L, particularly from 48 h onward (p < 0.0001). Similarly, gill lesions, including lamellar collapse, swelling, and cuticle rupture, intensified across all time points at higher concentrations (p < 0.0001). The most severe gill damage was observed at 3 mg/L after 96 h, with peak index values of 21.33 ± 4.62 for swelling and 16.00 ± 0.00 for cuticle rupture. Hemocyte infiltration also rose significantly at ≥2.25 mg/L, particularly after 72 and 96 h (p < 0.05). These quantitative data support the progressive tissue deterioration seen in Figure 7 and confirm that 2,4-D dimethylammonium induces significant histological damage in both digestive and respiratory tissues of E. dugasti.
Table 1 Frequency and severity index of histological alterations in the hepatopancreas and gill tissues of E. dugasti exposed to 2,4-D dimethylammonium at concentrations of 0, 0.75, 1.5, 2.25, and 3 mg/L for 24, 48, 72, and 96 h. Values are expressed as mean ± SD. Different letters indicate statistically significant differences (p < 0.05).
Concentrations
|
24 h |
48 h |
72 h |
96 h |
|||||
F |
Index |
F |
Index |
F |
Index |
F |
Index |
||
Hepatopancreas |
|
||||||||
Distension of lumen |
|||||||||
Control |
0.00 |
0.00 ± 0.00a |
0.00 |
0.00 ± 0.00a |
0.00 |
0.00 ± 0.00a |
0.00 |
0.00 ± 0.00a |
|
0.75 mg/L |
1.11 |
0.67 ± 1.15a |
2.22 |
0.67 ± 1.15a |
3.33 |
2.00 ± 0.00a |
5.56 |
2.00 ± 0.00ab |
|
1.5 mg/L |
3.33 |
2.00 ± 0.00b |
4.44 |
2.00 ± 0.00a |
4.44 |
2.00 ± 0.00a |
7.78 |
4.67 ± 2.31bc |
|
2.25 mg/L |
4.44 |
2.00 ± 0.00b |
5.56 |
2.00 ± 0.00a |
6.67 |
2.00 ± 0.00a |
8.89 |
6.67 ± 1.15cd |
|
3 mg/L |
6.67 |
2.00 ± 11.55b |
7.78 |
6.00 ± 3.46b |
8.89 |
6.67 ± 2.31b |
11.11 |
8.00 ± 0.00d |
|
p - value |
|
0.070 |
|
<.0001 |
|
<.0001 |
|
<.0001 |
|
Vacuolation |
|
|
|
|
|
|
|
|
|
Control |
0.00 |
0.00 ± 0.00a |
0.00 |
0.00 ± 0.00a |
0.00 |
0.00 ± 0.00a |
0.00 |
0.00 ± 0.00a |
|
0.75 mg/L |
1.11 |
0.67 ± 1.15ab |
2.22 |
1.33 ± 1.15a |
3.33 |
2.00 ± 0.00a |
4.44 |
2.00 ± 0.00ab |
|
1.5 mg/L |
2.22 |
1.33 ± 1.15ab |
4.44 |
2.67 ± 1.15a |
5.56 |
2.67 ± 1.15a |
6.67 |
2.00 ± 0.00ab |
|
2.25 mg/L |
3.33 |
1.33 ± 1.15ab |
5.56 |
2.00 ± 0.00a |
6.67 |
2.00 ± 0.00a |
7.78 |
4.67 ± 3.06b |
|
3 mg/L |
4.44 |
2.00 ± 00b |
7.78 |
4.00 ± 3.46b |
10.00 |
6.00 ± 3.46b |
12.22 |
9.33 ± 2.31c |
|
p - value |
|
0.080 |
|
<.0001 |
|
<.0001 |
|
<.0001 |
|
Gill |
|
||||||||
Lamellar collapse |
|||||||||
Control |
0.00 |
0.00 ± 0.00a |
0.00 |
0.00 ± 0.00a |
0.00 |
0.00 ± 0.00a |
0.00 |
0.00 ± 0.00a |
|
0.75 mg/L |
1.11 |
1.33 ± 2.31ab |
2.22 |
2.67 ± 2.31ab |
3.33 |
5.33 ± 2.31ab |
4.44 |
5.33 ± 2.31ab |
|
1.5 mg/L |
2.22 |
2.67 ± 2.31ab |
3.33 |
5.33 ± 2.31ab |
5.56 |
5.33 ± 2.31ab |
6.67 |
5.33 ± 2.31ab |
|
2.25 mg/L |
3.33 |
2.67 ± 2.31ab |
4.44 |
5.33 ± 2.31ab |
6.67 |
5.33 ± 2.31ab |
7.78 |
10.67 ± 6.93bc |
|
3 mg/L |
4.44 |
5.33 ± 2.31b |
5.56 |
8.00 ± 6.93b |
7.78 |
8.00 ± 6.93b |
10.00 |
13.33 ± 6.11c |
|
p - value |
|
0.045 |
|
<.0001 |
|
<.0001 |
|
<.0001 |
|
Tissue swelling |
|
|
|
|
|
|
|
|
|
Control |
0.00 |
0.00 ± 0.00a |
0.00 |
0.00 ± 0.00a |
0.00 |
0.00 ± 0.00a |
0.00 |
0.00 ± 0.00a |
|
0.75 mg/L |
0.00 |
0.00 ± 0.00a |
1.11 |
1.33 ± 2.31a |
3.00 |
4.00 ± 0.00ab |
4.44 |
4.00 ± 0.00a |
|
1.5 mg/L |
2.22 |
2.67 ± 2.31b |
4.44 |
4.00 ± 0.00ab |
5.56 |
4.00 ± 0.00ab |
6.67 |
4.00 ± 0.00a |
|
2.25 mg/L |
3.33 |
4.00 ± 0.00b |
5.56 |
4.00 ± 0.00ab |
7.78 |
8.00 ± 6.93b |
8.89 |
10.67 ± 4.62b |
|
3 mg/L |
6.67 |
4.00 ± 0.00b |
7.78 |
9.33 ± 6.11b |
12.22 |
16.00 ± 0.00c |
14.44 |
21.33 ± 4.62c |
|
p - value |
|
<.0001 |
|
<.0001 |
|
<.0001 |
|
<.0001 |
|
Cuticle rupture |
|||||||||
Control |
0.00 |
0.00 ± 0.00a |
0.00 |
0.00 ± 0.00a |
0.00 |
0.00 ± 0.00a |
0.00 |
0.00 ± 0.00a |
|
0.75 mg/L |
0.00 |
0.00 ± 0.00a |
1.11 |
0.00 ± 2.31a |
2.22 |
2.00 ± 2.31a |
3.33 |
4.00 ± 0.00ab |
|
1.5 mg/L |
2.22 |
2.67 ± 2.31ab |
3.00 |
4.00 ± 0.00b |
4.44 |
4.00 ± 0.00ab |
5.56 |
4.00 ± 0.00ab |
|
2.25 mg/L |
4.44 |
4.00 ± 0.00b |
5.56 |
4.00 ± 0.00b |
6.67 |
4.00 ± 0.00ab |
7.78 |
8.00 ± 6.93b |
|
3 mg/L |
5.56 |
5.33 ± 2.31b |
7.78 |
5.33 ± 2.31b |
10.00 |
9.33 ± 6.11b |
12.22 |
16.00 ± 0.00c |
|
p - value |
|
<.0001 |
|
<.0001 |
|
<.0001 |
|
<.0001 |
|
Infiltration of hemocytes |
|||||||||
Control |
0.00 |
0.00 ± 0.00a |
0.00 |
0.00 ± 0.00a |
0.00 |
0.00 ± 0.00a |
0.00 |
0.00 ± 0.00a |
|
0.75 mg/L |
0.00 |
0.00 ± 0.00a |
0.00 |
0.00 ± 0.00a |
1.11 |
1.33 ± 2.31a |
2.22 |
2.67 ± 2.31ab |
|
1.5 mg/L |
0.00 |
0.00 ± 0.00a |
1.11 |
1.33 ± 2.31ab |
1.11 |
1.33 ± 2.31a |
3.33 |
2.67 ± 2.31ab |
|
2.25 mg/L |
2.22 |
1.67 ± 2.08ab |
3.33 |
3.00 ± 1.73b |
4.44 |
3.00 ± 1.73a |
4.44 |
3.00 ± 1.73ab |
|
3 mg/L |
4.44 |
3.00 ± 1.73b |
5.56 |
3.00 ± 1.73b |
6.67 |
3.00 ± 1.73a |
7.78 |
5.33 ± 2.31b |
|
p - value |
|
0.000 |
|
0.000 |
|
0.045 |
|
0.017 |
|
Immunohistochemistry analysis
Figure 8 shows the immunohistochemical localization of AChE in the hepatopancreas and gill tissues of rice field crabs exposed to 2,4-D dimethylammonium at concentrations of 0 (control), 0.75, 1.5, 2.25, and 3 mg/L for 24, 48, 72, and 96 h. Positive AChE expression was indicated by dark brown to black staining. In the hepatopancreas, staining was primarily observed in the resorptive (R) cells and fibrillar (F) cells. In the gill tissue, strong AChE localization was noted in the epithelial cells of the lamellae and in circulating hemocytes. Staining intensity decreased progressively with increasing concentration and exposure time, reflecting suppressed AChE expression in both tissues under herbicide stress.
OCT analysis
OCT has been used to study the effects of the herbicide 2,4-D dimethylammonium on the hepatopancreas and gill of the rice field crab. The study found that changes in the hepatopancreas occur after 48 h of exposure to the chemical (data not shown), but the most significant changes occur after 96 h of exposure (Figure 9). In the control group, the hepatopancreas cells appear normal with a smooth surface, whereas in the exposed group, the structure of the hepatopancreas is damaged, with spaces inside the cells and a rough surface (indicated by the white arrows). In the gill, changes first occur after 24 h of exposure, but only slight changes are observed (data not shown). The most significant changes are seen after 96 h of exposure, where swelling in the gill raker cells and damage to the gill lamellae are observed (Figure 10).
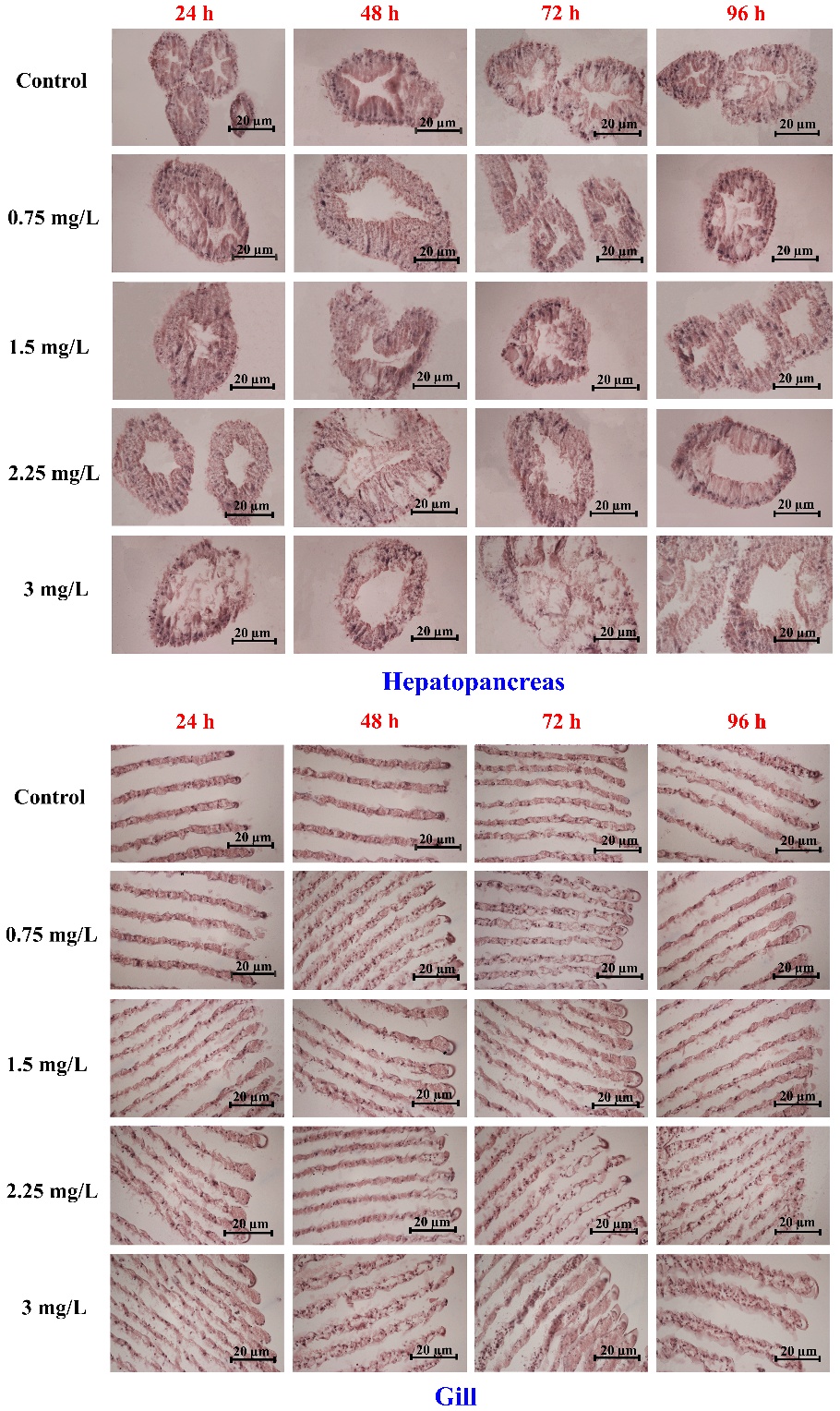
Figure 8 Immunohistochemical localization of AChE in the hepatopancreas and gill tissues of rice field crabs (E. dugasti) exposed to 2,4-D dimethylammonium (0 - 3 mg/L) for 24 - 96 h. Scale bars = 20 µm.
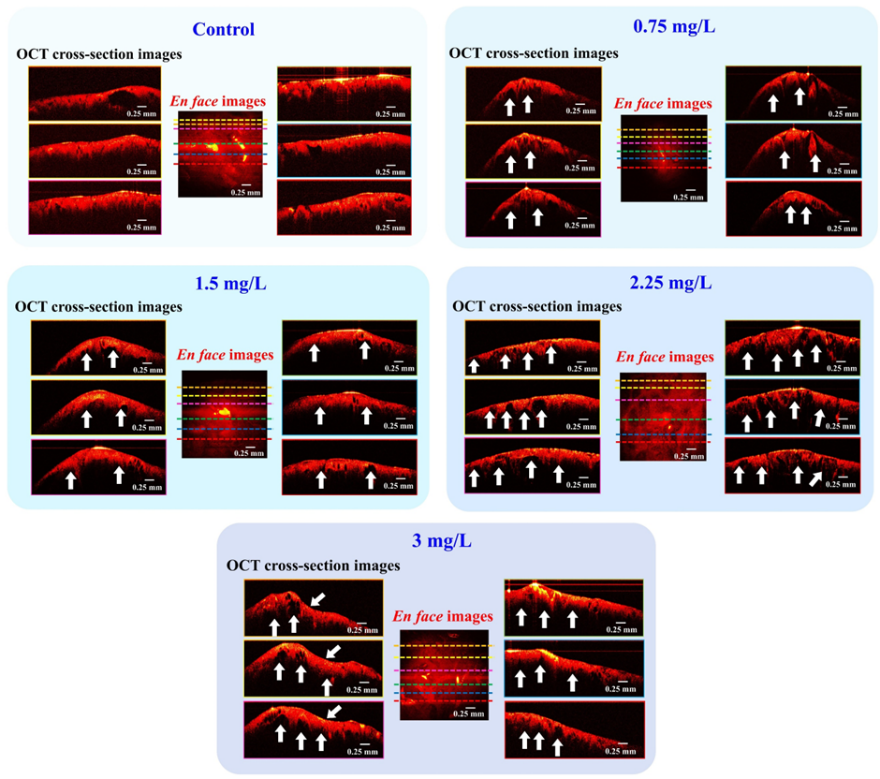
Figure 9 Representative OCT cross-sectional and en face images of hepatopancreas tissues from rice field crabs exposed to 2,4-D dimethylammonium at concentrations of 0 (control), 0.75, 1.5, 2.25, and 3 mg/L for 96 h. Note: White arrows indicate areas of altered tissue.

Figure 10 Representative OCT cross-sectional and en face images of gill tissues from rice field crabs exposed to 2,4-D dimethylammonium at concentrations of 0 (control), 0.75, 1.5, 2.25, and 3 mg/L for 96 h. Note: White arrows indicate areas of altered tissue.
Discussion
Aquatic organisms are routinely exposed to environmental contaminants through gill absorption, integument contact, and ingestion [21]. Such exposures can impair growth, alter behavior, reduce fertility, and increase mortality. In this study, E. dugasti exhibited dose- and time-dependent mortality when exposed to 2,4-D dimethylammonium, with LC₅₀ values decreasing over prolonged exposure durations—consistent with findings in S. bangkokensis [18].
Deyashi and Chakraborty [10] reported that crustaceans are often exposed to a variety of environmental toxicants, pollutants, and pathogenic organisms in their aquatic environments. Like other invertebrates, they lack adaptive immunity and therefore rely heavily on their innate immune system, which serves as the primary defense mechanism against foreign invaders and pathogens. Circulating hemocytes play a central role in immune defense, stress regulation, and tissue homeostasis. Crustaceans rely primarily on innate immunity, mediated by circulating hemocytes, due to the absence of adaptive immune systems. Our results showed that 2,4-D dimethylammonium significantly reduced total hemocyte counts and induced morphological abnormalities across hemocyte types large granule, small granule, and hyalinocytes (types 1 - 4). These abnormalities, especially in cytoplasmic and granular structures, increased with both concentration and exposure time, echoing observations by Kumla et al. [8] and Gupta et al. [22]. Abnormalities identified were primarily in the granular cells and cytoplasm, causing morphological alterations when compared to the control group. This finding aligns with a similar study by Kumla et al. [8], which investigated the effects of 2,4-D dimethylammonium on S. bangkokensis. The study found that the abnormalities in hemocytes were influenced by both exposure time and concentration, particularly in the cytoplasm and granular cells. However, the degree of abnormality also depended on the type of hemocyte. Furthermore, this study is consistent with the findings of Gupta et al. [22], which reported that exposure to 2,4-D dimethylammonium led to abnormal shapes of hemocytes and damage to the granules.
Additionally, the activity of AChE and BChE decreased significantly with prolonged exposure and higher herbicide concentrations, indicating neurotoxicity. These findings align with prior studies in S. bangkokensis and Barytelphusa guerini [8,14], where inhibition of cholinesterase activity persisted even after removal from the contaminated environment.
Protein analysis further supported these biochemical findings. SDS-PAGE and Western blot revealed reduced AChE expression, with a consistent molecular weight of ~71 kDa, particularly at higher concentrations and longer exposures. These results are corroborated by similar reports in Nile tilapia, hybrid catfish, and prawn [7,23,24] The observed downregulation may reflect herbicide-induced cytotoxicity in AChE-producing cells.
As biomarkers, biochemical and protein responses offer a powerful tool for pollutant monitoring [25]. In this study, hemolymph, selected for its high enzymatic activity [26], revealed two AChE-related protein bands (73 and 71 kDa), which both diminished in size and intensity with exposure. Western blot confirmed that AChE expression decreased proportionally to concentration and exposure duration. These findings reinforce those of Kumla et al. [8] and Khanchanasal et al. [5], suggesting that AChE expression is a reliable biomarker of pesticide exposure across species.
Histological analysis revealed concentration-dependent structural damage in both the gills and hepatopancreas. Gill lesions included lamellar collapse, cuticle rupture, tissue swelling, and hemocyte infiltration, consistent with impaired respiratory function [27]. The hepatopancreas exhibited lumen distension and vacuolation, reflecting disruptions in digestion and detoxification pathways [28]. Immunohistochemistry confirmed reduced AChE localization in these tissues, validating its biomarker potential and paralleling results observed in S. bangkokensis [8].
To complement the histological findings, we employed OCT, a non-destructive technique that enables real-time, high-resolution imaging. OCT analysis revealed early and progressive morphological changes in both gills and hepatopancreas. In gills, lamellar disorganization and swelling were evident by 96 h. In hepatopancreas tissue, surface roughening and internal voids emerged with increasing severity. These findings correspond with prior OCT studies in prawn and Nile tilapia [15,29] reinforcing OCT’s value in ecotoxicological assessment. Kumla et al. [8] similarly demonstrated OCT’s sensitivity in detecting herbicide-induced morphological disruptions in S. bangkokensis, noting severe gill and hepatopancreas damage even at low concentrations after 48 h.
Collectively, these findings indicate that enzymatic suppression, particularly of cholinesterases, acts in concert with immunotoxic effects to destabilize tissue structure and function, ultimately contributing to respiratory and digestive failure and increased mortality. The integration of biochemical, histological, and OCT-based evidence provides a cohesive mechanistic framework for understanding 2,4-D dimethylammonium toxicity in E. dugasti.
The toxicity of 2,4-D dimethylammonium observed in this study is likely mediated by multiple interacting cellular mechanisms. 2,4-D dimethylammonium has been reported to induce oxidative stress and mitochondrial dysfunction, leading to impaired energy metabolism and cellular homeostasis. In addition, its amphiphilic properties may destabilize cellular membranes, increasing permeability and promoting vacuolation and tissue disorganization. Although classical endocrine disruption is not well defined in crustaceans, interference with neuroendocrine-like and cholinergic signaling may further impair neuromuscular control, immune function, and tissue integrity. These mechanisms provide a plausible explanation for the observed cholinesterase inhibition, hemocyte dysfunction, and progressive gill and hepatopancreas damage detected by histology and OCT.
While this study provides a comprehensive acute toxicity assessment of 2,4-D dimethylammonium in Esanthelphusa dugasti, several limitations should be acknowledged. The experiments focused on short-term exposure and therefore do not capture chronic, recovery, or multigenerational effects that may occur under environmentally realistic conditions. In addition, adult rice field crabs possess a hard exoskeleton that strongly limits OCT penetration depth; therefore, OCT imaging was performed ex vivo, which may not fully represent dynamic physiological processes occurring in vivo. The absence of direct oxidative stress and mitochondrial biomarkers also limits confirmation of specific molecular pathways inferred from the observed biochemical and histological changes. Furthermore, controlled laboratory conditions cannot fully replicate the complexity of natural rice-field ecosystems, where multiple stressors and fluctuating environmental parameters may influence toxicity. Future studies should incorporate chronic and recovery-based assays, oxidative stress biomarkers, gene expression and metabolomic profiling, and field or mesocosm validation to better resolve toxicity mechanisms and ecological relevance.
Although the LC₅₀ and LOEC/NOEC values reported in this study were derived under controlled laboratory conditions and should not be interpreted as direct field application thresholds, they provide important insight into potential risk scenarios in rice-field ecosystems. The clear concentration- and time-dependent toxic responses observed across mortality, neurotoxicity, immunotoxicity, and tissue damage suggest that Esanthelphusa dugasti may be particularly vulnerable to short-term pulse exposures of 2,4-D dimethylammonium. Such pulse events may occur following herbicide application combined with rainfall, irrigation discharge, or rapid water exchange in rice fields. Therefore, the present findings do not necessarily indicate an immediate need for application dose modification, but rather highlight the importance of application timing and water management practices, including avoiding application prior to heavy rainfall, implementing controlled drainage, and maintaining buffer zones. These measures may substantially reduce acute exposure risk to non-target crustaceans while preserving herbicide efficacy. Further field-based and chronic exposure studies are required to refine long-term risk management strategies.
Conclusions
This study provides an integrated assessment of 2,4-D dimethylammonium toxicity in the rice field crab E. dugasti, demonstrating that acute exposure elicits coordinated neurotoxic, immunotoxic, and structural tissue responses. By combining conventional biochemical and histological analyses with OCT, this work advances beyond endpoint-specific toxicity evaluation and highlights the value of depth-resolved, non-destructive imaging for linking molecular dysfunction to organ-level pathology in aquatic invertebrates.
While the findings clearly indicate susceptibility of E. dugasti to short-term herbicide exposure, the present study was limited to acute toxicity scenarios and does not address potential sub-chronic, chronic, or recovery effects that are critical for comprehensive ecological risk assessment. In addition, OCT imaging was performed ex vivo, and laboratory conditions cannot fully capture the complexity of natural rice-field ecosystems. These limitations underscore the need for future studies incorporating long-term exposure designs, and molecular biomarkers, and field- or mesocosm-based validation.
Overall, this study establishes OCT as a powerful complementary tool for mechanistic ecotoxicology and provides a framework for future investigations aimed at improving environmental monitoring, informing herbicide management strategies, and supporting conservation of non-target organisms in agricultural ecosystems.
Acknowledgements
This research was supported by Suranaree University of Technology (SUT), Nakhon Ratchasima 30000, Thailand Science Research and Innovation (TSRI), and the National Science, Research, and Innovation Fund (NSRF) (NRIIS number 204303).
Declaration of Generative AI in Scientific Writing
The author utilized a generative AI tool (e.g., ChatGPT) to assist in refining the language, improving the eloquence, and ensuring grammatical accuracy in portions of this manuscript. However, the author retains full responsibility for the accuracy of all content, ideas, and conclusions.
CRediT Author Statement
Chutima Thanomsit: Conceptualization; Methodology; Original draft preparation. Sugunya Kumla: Investigation; Methodology, Sawipa Ruttanakorn: Methodology; Investigation, Resource. Witchuda Prasatkaew: Methodology; Investigation. Panomsak Meemon: Visualization; Investigation, Jakkaphun Nanuam: Review& Editing/Conclusion; Data analysis; and Phochit Nanthanawat: Supervision; Methodology;
References
[1] AF Vigário and SMT Sabóia-Morais. Effects of the 2,4-D herbicide on gills epithelia and liver of the fish Poecilia vivipara. Pesquisa Veterinaria Brasileira 2014; 34(6), 523-528.
[2] CA Jote. A review of 2,4-D environmental fate, persistence and toxicity effects on living organisms. Organic & Medicinal Chemistry International Journal 2019; 9(1), 555755.
[3] MT Halter. 2,4-D in the Aquatic Environment. Municipality of Metropolitan Seattle, Washington, 1980.
[4] M Casimero, MJ Abit, AH Ramirez, NG Dimaano and J Mendoza. Herbicide use history and weed management in Southeast Asia. Advances in Weed Science 2022; 40(1), 020220054.
[5] P Khanchanasal, C Nuankaew, S Saowakoon, J Nanuam, P Nanthanawat and C Thanomsit. Toxicity level of 2, 4-D dimethylammonium in Nile tilapia and Acetylcholinesterase (AChE) Expression (Biomarker) to identify exposure in sub-lethal concentration. Burapha Science Journal 2022; 27(2), 1278-1299.
[6] C Thanomsit, P Khanchanasal, W Prasatkaew, J Nanuam, P Meemon, A Wattanakornsiri and P Nanthanawat. Adverse effects of 2, 4-D dimethylammonium based-herbicide on Acetylcholinesterase expression in Nile tilapia (Oreochromis niloticus). Environmental Toxicology and Pharmacology 2024; 106, 104383.
[7] S Kumla, B Chartchumni, P Nanthanawat, J Nanuam, S Ruttanakorn, P Meemon and C Thanomsit. Morphological, biochemical, and histological alterations in juvenile giant freshwater prawns (Macrobrachium rosenbergii) exposed to sub-lethal concentrations of 2,4-D dimethylammonium. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology 2024; 295, 110212.
[8] S Kumla, N Senasri, P Nanthanawat, J Nanuam, P Meemon, S Udduang and C Thanomsit. Comprehensive evaluation of 2,4-D dimethylammonium toxicity in rice field crabs (Somanniathelphusa bangkokensis). Environmental Toxicology and Pharmacology 2025; 117, 104725.
[9] K Thaiso and W Wongmaneeprateep. Effects of different dietary protein sources on growth and survival rates of rice field crab (Esanthelphusa dugasti) Nursing. Khon Kaen Agriculture Journal 2012; 40, 123-128.
[10] C Thanomsit, S Saowakoon, A Wattanakornsiri, J Nanuam, W Prasatkaew, P Nanthanawat, P Mongkolvai and W Chalorcharoenying. Glyphosate (Roundup): Fate in aquatic environment, adverse effect and toxicity assessment in aquatic organisms. Naresuan University Journal: Science and Technology 2020; 28(1), 65-81.
[11] OECD. Test guideline No.203 Fish, acute toxicity testing. OECD, France, 2025.
[12] M Deyashi and SB Chakraborty. Cytomorphological characterization, classification and counting of haemocytes in freshwater crab, Varuna litterata (Crustacea: Decapoda). Invertebrate Zoology 2022; 19(2), 120-134.
[13] MM Bradford. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry 1976; 72(1-2), 248-254.
[14] MR Narra. Effects of chlorpyrifos insecticide, on cholinesterase activity and its depuration in the crab Barytelphusa guerini. International Journal of Environmental Technology and Management 2015; 18(1), 1-8.
[15] C Thanomsit, S Kumla, J Saetiew, J Saenjae, P Nanthanawat, J Nanuam and P Meemon. Application of Optical Coherence Tomography (OCT) to evaluate the adverse effects of 2,4-D dimethylammonium on morphological changes in Riceland Prawn (Macrobrachium lanchesteri). Environmental Toxicology and Pharmacology 2025; 113, 104608.
[16] GL Ellman, KD Courtney, VA Jr and RM Featherstone. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochemical pharmacology 1961; 7(2), 91-95.
[17] E Escartín and C Porte. The use of cholinesterase and carboxylesterase activities from Mytilus galloprovincialis in pollution monitoring. Environmental Toxicology and Chemistry 1997; 16(10), 2090-2095.
[18] D Bernet, H Schmidt, W Meier, P Burkhardt‐Holm and T Wahli. Histopathology in fish: Proposal for a protocol to assess aquatic pollution. Journal of Fish Diseases 2001; 22(1), 25-34.
[19] C Thanomsit, J Saetiew and P Meemon. Optical coherence tomography as an alternative tool for evaluating the effects of glyphosate on hybrid catfish (Clarias gariepinus × Clarias macrocephalus). Toxicology Reports 2022; 9, 181-190.
[20] J Saetiew, J Saenjae,C Thanomsit, P Nanthanawat, A Wattanakornsiri and P Meemon. Development of a field-ready optical coherence tomography system for assessing 2,4-D dimethylammonium-based herbicide exposure in African catfish. Optical Engineering 2024; 63(2), 024102.
[21] N Krishnan. Aquatic Toxicity Tests. Wikibooks, England, 2019.
[22] RK Gupta, J Sharma and A Vohra. Identification of different types of haemocytes in freshwater crab Paratelphusa masoniana (Henderson). International Journal of Aquaculture and Fishery Sciences 2013; 3(1), 7-12.
[23] C Nualkaw, A Wattanakornsiri, J Nanuam, P, Nanthanawat, W Prasatkaew, C Meeprom, S Saowakoon and C Thanomsit. Effects of paraquat-based herbicide on acetylcholinesterase (AChE), antioxidant status and histological alterations in freshwater fish: Nile tilapia (Oreochromis niloticus). Aquaculture, Aquarium, Conservation & Legislation 2022; 15(6), 3197-3211.
[24] C Thanomsit, P Kiatprasert, W Prasatkaew, N Khongchareonporn and P Nanthanawat. Acetylcholinesterase (AChE) monoclonal antibody generation and validation for use as a biomarker of glyphosate-based herbicide exposure in commercial freshwater fish. Comparative Biochemistry and Physiology Part - C: Toxicology and Pharmacology 2021; 241, 108956.
[25] CH Walker, RM Sibly, SP Hopkin and DB Peakall. Principles of ecotoxicology. Taylor & Francis, Oxfordshire, 2012.
[26] NN Kovalev, EV Mikheev and AI ChepkasovaI. Cholinesterase activity in tissues of some crab species from the Japan Sea. Journal of Evolutionary Biochemistry and Physiology 2012, 48(4), 409-413.
[27] CL Negro. Histopathological effects of endosulfan to hepatopancreas, gills and ovary of the freshwater crab Zilchiopsis collastinensis (Decapoda: Trichodactylidae). Ecotoxicology and Environmental Safety 2015; 113, 87-94.
[28] A Maharajan, Y Narayanasamy, V Ganapiriya and K Shanmugavel. Histological alterations of a combination of Chlorpyrifos and Cypermethrin (Nurocombi) insecticide in the fresh water crab, Paratelphusa jacquemontii (Rathbun). The Journal of Basic & Applied Zoology 2015; 72, 104-112.
[29] C Thanomsit, S Saowakoon, W Prasatkaew, S Ruttanakorn, J Nanuam, C Kangpanich, K Sreeputhorn and P Meemon. Optical Coherence Tomography (OCT) alterations choice for aquatic organism anatomy and morphology analysis: A case study of A Giant River Prawn (Macrobrachium Rosenbergii). Chiang Mai Journal of Science 2024; 51(1), 186669.