Trends
Sci.
2026; 23(8): 13091
Lipid Profile Improvement of Moringa oleifera Leaf Powder from Different Leaf Stalk Colors: In Vitro and In Vivo Studies
Lina Novi Ariani1, Teti Estiasih2,3,*, Wenny Bekti Sunarharum2,
Alfi Khatib4, Sentot Joko Raharjo5 and Ambar Fidyasari3
1Doctoral Program of Food Science, Department of Food Science and Biotechnology,
Faculty of Agricultural Technology, Universitas Brawijaya, Malang 65145, Indonesia
2Department of Food Science and Biotechnology Department, Faculty of Agricultural Technology,
Universitas Brawijaya, Malang 65145, Indonesia
3Center for Local Food Development Studies, Universitas Brawijaya, Malang 65145, Indonesia
4Kulliyyah of Pharmacy, International Islamic University Malaysia, Kuantan, Pahang 25200, Malaysia
5Food and Pharmaceutical Analysist Department, Politeknik Kesehatan Putra Indonesia Malang,
Jawa Timur Malang 65141, Indonesia
(*Corresponding author’s e-mail: [email protected])
Received: 19 December 2025, Revised: 21 January 2026, Accepted: 28 January 2026, Published: 20 March 2026
Abstract
Moringa oleifera (MO) has long been known for traditional diseases preventions and treatments. This tree has wide genetic variabilities such as the distinctive leaf stalk colors. Moringa oleifera leaf powder (MOLP) is potentially used for a functional food ingredient. The high prevalence of hypercholesterolemia requires a search for ingredients for cholesterol-lowering food products. This study aimed to evaluate the ability of MOLP from different leaf stalk colors (red, white, green) in improving lipid profile in vivo. The mechanism of improvement was evaluated by in vitro HMG-CoA and pancreatic lipase inhibitory activities of MOLP extract and in vivo fecal concentration. The results showed that the highest HMG-CoA reductase inhibitory activity was obtained from the MOLP extract with white leaf stalk, followed by red and green ones. The highest inhibition was found in the aqueous extract of MOLP from red leaf stalk with an inhibitory activity of 81.06%, almost comparable to statin of 85.20%. The sequence of the HMG -CoA reductase activity from the highest was the water extract of MOLP from red leaf stalk (81.06%), followed by ethyl acetate extract of MOLP from green leaf stalk (97.30%), water extract of MOLP from white leaf stalk (75.72%). The highest lipase inhibition activity was exhibited by MOLP with red leaf stalk followed by white and green ones. MOLP with different leaf stalk colors reduced LDL-c. TG, and TC, but increased HDL-c significantly, that MOLP from the red laef stalk was the best, followed by white and green ones. The number of identified phytochemicals from ethanol extracts was influenced by leaf stalk color. Flavonoids were the predominant phytochemicals for all MOLP extracts from different leaf stalk colors. MOLP increased fecal cholesterol concentration indicating inhibition of cholesterol absorption. MOLP has more advantages than MO leaf extract in modulating lipid profile due to its composition.
Keywords: Moringa oleifera, Hypercholesterolemia, In vitro, In vivo, HMG-CoA reductase, Pancreatic lipase
List of abbreviations
HDL-c High density lipoprotein-cholesterol
HMG-CoA 3-hydroxy-3-methylglutaryl coenzyme A
LC MS Liquid chromatography – mass spectroscopy
LDL-c Low density lipoprotein-cholesterol
MO Moringa oleifera
MOLP Moringa oleifera Leaf Powder
TC Total cholesterol
TG Triglyceride
SCFA Short chain fatty acid
VLDL-c Very high-density lipoprotein-cholesterol
Introduction
The Moringa genus is a medicinal plant used traditionally for a long time. Moringa species are known for their anti-inflammatory, anticancer, antioxidant and antihyperglycemic activities due to the biological activity of glucosides, flavonoids and glucosinolates [1]. One of the genera of Moringa that is widely consumed in many parts of the world is Moringa oleifera (MO) or drumstick. This species is known as the Miracle Tree [1], because all parts of this plant are used for various purposes and some organs show health benefits. Among MO plant organs, the most widely studied are MO leaves and they showed beneficial effects in several chronic disorders such as hypertension, hypercholesterolemia, insulin resistance, diabetes, cancer, non-alcoholic liver disease, and overall inflammation [2].
MO leaves attract attention due to their pharmacological activity and are widely consumed because of their nutritional content. Study of the benefits of MO leaves for health, especially in the form of extracts such as antidiabetic activities of the aqueous extract by α-glucosidase and α-amylase inhibition [3], protection of pancreatic beta cells, and the induction of glycogen synthesis by water extract [4], methanol and ethanol extracts inhibited α-amylase [5-7] and α-glucosidase [8], participating in the PI3K-Akt signaling pathway of ethanolic extract [9]; reduction lead toxicity [10]; malnutrition therapy of aqueous extract [11] hypolipidemic effect by inhibiting pancreatic lipase by ethanolic extract [8] and hypocholesterolemic effect of polyphenol extract from MO leaves [12].
Hypercholesterolemia is an abnormality of lipid profile in the blood leading to strokes and heart attacks [13] and an important precursor to many cerebrovascular, cardiovascular, and peripheral vascular diseases. Globally the prevalence of hypercholesterolemia is high, as reported by the American Heart Association that the prevalence of hypercholesterolemia to be 11.9 [14]. Finding natural
products for the management of hypercholesterolemia that mimic anti-cholesterol drugs remains a challenge. The increasing prevalence of hypercholesterolemia in the world has prompted the exploration of phytochemical-rich plants for the treatment of hypercholesterolemia. Traditionally, MO leaves have been used for hypercholesterolemia treatments such as by Indians as a hypocholesterolemic agent in their herbal medicine for obese patients [15]. Previous studies reported the pharmacological activities of MO leaf powder (MOLP), including hypoglycemic [16], antioxidant [17], anticancer, anti-inflammation [18], and hypocholesterolemic [19,20] properties.
Mechanisms of hypocholesterolemic effect of MO leaves has been evaluated in several studies. Asrifah et al. [19] indicated that MOLP had the ability to inhibit cholesterol absorption, indicating by high cholesterol concentration in the feces of MOLP fed rats. Besides, MOLP was able to produce short chain fatty acids (SCFAs) in the caecum. The polyphenol extract of MO leaves inhibited HMG-CoA reductase activity as the key enzyme in cholesterol synthesis, and bound to fecal bile acid [12]. It is supposed that HMG-CoA reductase and cholesterol absorption inhibition is related to the bioactive compounds of MO.
Forty-two compounds were identified from the water extract of MO leaves, including phenolic acids and their derivatives, isothiocyanates, flavonoids, alkaloids, nucleosides, and other compounds. Pharmacokinetic study on 5 compounds from water extract of MO leaves showed that all compounds were found in the blood plasma after 5 minutes oral administration indicating their high bioavailability [21]. Fidyasari et al. [22] showed that MOLP from different colors of leaf stalks, i.e. green, red, and white, had different total phenolic and flavonoid contents.
Asrifah et al. [19] examined the cholesterol-decreasing properties of MOLP from white and red leaf stalks. That study compared MOLP with ezetimibe and commercial fermented dairy product containing plant stanol ester as the drug and functional food to inhibit cholesterol absorption, respectively. The result showed that hypocholesterolemic activity of MOLP was similar to commercial fermented dairy products containing plant stanol ester and ezetimibe. MOLP from red leaf stalk exhibited better hypocholesterolemic properties. However, that study did not compare with statin drug as HMG-CoA reductase inhibitor and did not evaluate the ability of the MOLP to inhibit that enzyme.
Redha et al. [24] reviewed the roles of MO leaf extracts as anti-obesity through impairment of the lipid profile (total cholesterol (TC), low density lipoprotein cholesterol (LDL-c), and high density lipoprotein cholesterol (HDL-c)) and body weight. Ezzat et al. [25] revealed that ethanolic extract of MO leaves had anti-obesity activity in high fat diet rats through improving lipid profile. One of anti-obesity mechanism is by inhibiting pancreatic lipase activity thus lowered the TC absorption. Ethanol extract of MO leaves also showed a promising in vitro pancreatic lipase inhibitor (IC50 = 437.1 μg/mL) [26]. Aqueous extract of MO leaves also showed that pancreatic lipase and cholesterol esterase activity inhibition by MO leaves aqueous extract [5].
So far, the comparison of MOLP from different leaf stalk colors has been limitedly explored as a source for its cholesterol-lowering properties. Its anti-cholesterol mechanism has not been fully elucidated. This study aimed to evaluate the effect of MOLP from green, red, and white leaf stalks on lipid profile of hypercholesterolemia rats, in comparison to statin. We supposed that different leaf stalk colors have alight different bioactive compounds in the leaves thus the MOLP from different leaf stalk color might exhibit different hypocholesterolemia properties. The ability of MOLP extracts with different polarity of solvent to inhibit HMG-CoA reductase and lipase was also examined as well as the phytochemicals of the potential extracts. Most studies used fresh leaves for extracting as the part of extract for some purposes such as food supplements. This study used MOLP for extraction to prove that the MOLP phytochemicals with different polarity revealed lipase and HMG-CoA reductase inhibition activity. The ability of MOLP to inhibit cholesterol absorption was examined through fecal cholesterol concentration of the rats.
Materials and methods
Materials
MOLP was obtained from a local farmer (Figure 1), obtained from Bluto District, Sumenep Regency, East Java, Indonesia. The MOLP was taken only once during experiment to ensure the homogeneity of the samples. MOLP was stored in the room temperature in tight jars during the study. For Anticholesterol analysis through HMG-CoA reductase inhibition assay Enzymatic activity was measured using the HMG-CoA Reductase Assay Kit technical bulletin with catalog number CS1090 with reagen Assay Buffer 1X by dissolving 0.2 mL of assay buffer 5x with 0.8 mL of ultrapure water, 1 mL of assay buffer, NADPH, HMG-CoA, HMG-CoA Reductase (HMGR) and Pravastatin. Meanwhile, for anticholesterol analysis through Lipase pancreatic inhibition assay, enzymatic activity was measured using the pancreatic Lipase Assay Kit technical bulletin with catalog number MAK046 with glycerol standards reagent, lipase assay buffer, peroxidase substrate, enzyme mix and lipase substrate. DyaSis CHOD PAP was used for analysis of lipid profile in in vivo experiment.
Methods
MOLP maceration-sonification extraction
The MO leaves with different leaf stalk colors and their corresponding MOLP is shown in Figure 1. MOLP of white, green, and red stalk was extracted using maceration-sonication method with water (aquadest), ethanol, acetone, ethyl acetate and n-hexane, representing different solvent polarities. All organic solvents were pro-analysis grade from Merck (Germany). The polarity of solvent determined the extracted compounds that might affect the extract ability in reducing HMG-CoA reductase and lipase activity. Maceration-sonification extraction techniques following the procedure modified from Ref. 30. The process involved soaking 100 g of MOLP in 800 mL of the respective solvent for 3 h, then subjecting it to ultrasonic extraction for 30 min, set at 55 °C (200 W, 40 kHz). The extract was then filtered, and the residue was re-extracted twice using the same solvent. The combined filtrates were concentrated using a rotary evaporator under vacuum at 45 °C until thick, and the residual solvent was removed by spraying with nitrogen gas. Next, the extract was centrifuged for 10 min to remove solids that might have escaped during filtration. The resulting 40-50 mL filtrate was stored at 4 °C for further analysis. Each extract was analyzed for phytochemical types using LC-MS following the procedure of Teclegeorgish et al. [31].

Figure 1 The leaves of Moringa oleifera with different leaf stalk colors and their corresponding leaf powder; (a) Moleifera leaves with different leaf stalk color: (A) red, (B) green, (C) white, and (b) Moringa oleifera leaves powder with different leaf stalk color: (A) red, (B) green, and (C) white.
In vitro analysis of HMG-CoA reductase inhibition by MOLP extracts
HMG-CoA reductase inhibitory activity was calculated by measuring the absorbance at 340 nm using a BioTek ELX800 microplate reader (BioTek Instruments, US). MOLP extracts were analyzed for their inhibitory activities. HMG-CoA reductase inhibition assay of MOLP extracts were analyzed by following the protocol of HMG-CoA Reductase Assay Kit technical bulletin with catalog number CS1090 (Sigma-Aldrich Co.). As a comparison is statin drug. HMG-CoA reductase inhibition assay of MOLP extracts were analyzed by following the protocol of HMG-CoA Reductase Assay Kit technical bulletin with catalog number CS1090. As a comparison is statin drug.
In vitro analysis of lipase pancreatic inhibition by MOLP extracts
Pancreatic lipase inhibitory activity was calculated by measuring the absorbance at 570 nm using a BioTek ELX800 microplate reader (BioTek Instruments, US). MOLP extracts were analyzed for their inhibitory activities. Pancreatic lipase inhibition assay was measured by following the procedure from Pancreatic Lipase Assay Kit technical bulletin with catalog number MAK046 (Sigma-Aldrich Co.). Orlistat was used as a comparison of drug for lipase inhibition.
Phytochemical analysis of MOLP extracts by LC-MS
The extracts which showed the highest HMG-CoA reductase and pancreatic lipase inhibition activity were analyzed for their phytochemical profiles using LCMS-8040. The LCMS-8040 is the long-proven UFMS providing acquisition speed of 555 MRM/sec and 15 msec polarity switching time. It was equipped with UF-Lens and UF sweeper II collision cell, the hallmark technologies underlying fast ion transmission and high sensitivity.
Chromatographic separation was performed on (Ultra Performance Liquid Chromatography particle technology Bridged Ethylene Hybrid) or Ultra Performance Liquid Chromatography using particle technology bridged ethylene hybrid (UPLCBEH), equipped with a C18 column (100×2.1×1.7 mm3 film thickness, Waters, USA). The column temperature was set at 35 °C and the injection volume was set at 1 µL. Mobile phase A was 0.5 %(v/v) formic acid solution, while mobile phase B was Isocratic. And the flow rate was set at 0.5 mL/min. The change of mobile phase was programmed as follows: The initial mobile phase was 95% mobile phase A and 5% mobile phase B; mobile phase B increased from 5% to 100% of the total mobile phase within 11 min (1.5 min, 20%; 3.5 min, 40%; 5 min, 60%; 7 min, 85%; 8 min, 100%); at 11.5 min, the mobile phase was suddenly changed to 95% mobile phase A and 5% mobile phase B, and then the system was maintained for 1.5 min; then, the next cycle was started. The UPLC-Q-TOF-MS system was configured with a WatersXevo G2Q-TOF mass spectrometer (Milford, MA, USA). Mass spectra were obtained by electrosprayionization (ESI) with Io type [M]+ ion mode and scanning at 0.6 sec/scan (mz: 10 - 1,000). The MSE method was carried out in this study through the ascan model to obtain information on parent and daughter ions in one injection using tandem mass spectrometry, and the MS conditions were as follows: Capillary ionization voltage was 3.0 kV and sampling cone voltage was 23 V; source temperature was 100 °C and solvent temperature was 350 °C; desolvation gas flow rate was 60 mL/h; Collison energy 5.0 V. The scanning range of MS1 and MS2 was between 10 to 1,000 m/z, scanning 0.6 s/scan and collision energy ranged from 25 to 35 V. Data were collected and analyzed with WaterMassLynxv4.1 software.
In vivo experiment of lipid profile improvement by MOLP
The feed used in this study were the AIN-93M standard diet and a high-cholesterol modified AIN-93M standard diet. The formulation of MOLP from different stalk colors was based on the iso-calorie and iso-protein calculation by considering the proximate composition and dietary fiber content [22]. All formulas feed composition and modification of standard and high cholesterol feed according to the AIN-93M are shown in Table 1.
Male rats with 2 - 3 months age and body weight of 150-200 g were grouped into 6: Normal rats fed with standard diet, hypercholesteremia rats fed with standard diet, 3 groups of hypercholesterolemia rats fed with feed formulated by MOLP from white, red, and green leaf stalk colurs. After 3 days adaptation, rats were being hypercholesteromia by feeding hypercholesterolemia diets. After blood TC achieved 200 mg/dL, the treatment was started to employed by feeding according to the treatments. Every 2 weeks, blood were taken orbital plexus, and after centofugation at 3,000 rpm for 15 min, the TC, LDL-c, HDL-c, and triglyceride (TG) were analysis using DiaSys Kit Assay (CHOD PAP method).
Table 1 Composition of high cholesterol feed and modified standard AIN-93M [32] blood serum.
Component |
Composition (g/kg diet) |
||||
Modified Standard AIN-93M* |
Hypercholeste-rolaemic Diet ** |
MOLP from White Leaf Stalk |
MOLP from Red Leaf Stalk |
MOLP from Green Leaf Stalk |
|
MOLP |
- |
- |
603.71 |
495.75 |
474.74 |
Cornstarch |
620. 69 |
620. 69 |
424.64 |
438.05 |
438.66 |
Casein |
140 |
140 |
0 |
0 |
0 |
Sucrose |
100 |
100 |
100 |
100 |
100 |
Soybean Oil |
40 |
40 |
15.79 |
20.32 |
21.91 |
Fiber |
50 |
50 |
0 |
0 |
3.020 |
Mineral mix |
35 |
35 |
10 |
10 |
10 |
Vitamin mix |
10 |
10 |
20 |
20 |
20 |
L-sistin |
1.8 |
1.8 |
- |
- |
- |
Kolin bitartrat |
2.5 |
2.5 |
- |
- |
- |
TBHQ |
0.008 |
- |
- |
- |
- |
Cholic acid |
- |
2 |
- |
- |
- |
Cholesterol |
- |
20 |
- |
- |
- |
Beef tallow |
- |
185 |
- |
- |
- |
Dextrin |
|
|
155 |
155 |
155 |
*The standard feed composition as per the AIN-93M standard, established by Reeves (1993).
** Modification of Standard and High Cholesterol Feed According to AIN-93M".
Fecal cholesterol concentration analysis
At the end of experiment (week 8), TC of feces was measured using enzymatically CHOD-PAP with Dialabs® reagent (Germany), using Microlab® spectrophotometry at a wavelength of 546 nm. Feces were homogenized in 10% Tris HCl buffer solution 10 nM (pH 7.4), and continued with extraction using organic solvents based on the Folch method. The principle of cholesterol extraction in feces according to [33,34] is tissue extraction using organic solvents, namely chloroform: Methanol = 2:1 (v/v), which is continued with washing the resulting filtrate using water to obtain an extract containing cholesterol. The extract was then dried in a vacuum and re-suspended in saline (NaCl solution) containing sodium lauryl sulphate 0.1% in accordance with previous methods [35,36]. Sodium lauryl sulphate 0.1% in saline was used to improve the recovery of cholesterol in the extract of homogenate. This study involve experiments on animals or human subjects.
Ethical Clearance Certificate No. 101-KEP UB-2022 from the Research Ethics Committee (Animal Care and Use Committee) Universitas Brawijaya.
Data analysis
The experimental design to evaluate HMG-CoA reductase and pancreatic lipase activity inhibition was a completely randomized design with 2 factors: The MOLP from different leaf stalk colors (green, red, and white) and the extraction solvents (water, ethanol, acetone, ethyl acetate and n-hexane). All treatments were conducted in triplicate. The in vivo experiment was a nested experimental design with 6 treated groups of rats. Two factors were evaluated: The rat feeding and health status (normal rats fed with standard diet, hypercholesterolemia rats fed with standard diet, 3 groups of hypercholesterolemia rats fed with feed formulated by MOLP from white, red, and green leaf stalk colors), and time of feeding (0, 2, 4, 6 and 8 weeks).
The data were analyzed by Analysis of Variance, followed by the LSD test, to determine whether there was significant differences (α = 0.05) among treatments. LSD Test was conducted to identify the significant factors. The statistical tests were carried out with a 95% confidence level (α = 0.05). The analysis of data used Minitab version 19.
HMG-CoA reductase inhibitory activity of MOLP extracts from different leaf stalk colors
Figure 2(a) shows that MOLP extracts with different solvents have the ability to inhibit HMG-CoA reductase, the rate-limiting enzyme in the hepatic cholesterol synthesis pathway. Statistically (p < 0.05), the type of solvents and the colors of leaf stalk significantly affected the HMG-CoA reductase activity. The lowest activity was found in the water extract of MOLP from the red leaf stalk with the activity of 0.28 0.02 unit/mg P, which was almost similar to statin of 0.25 0.01 unit/mg P. The highest activity was observed in the hexane extract of MOLP from red leaf stalk. Different leaf stalk colors indicated different HMG-CoA reductase activity. The lowest enzyme activity of the extract MOLP from green leaf stalk was from ethyl acetate and the highest activity was from water extract. Meanwhile, the extract form MOLP with white stalk had the lowest activity in water extract and the highest was acetone extract.
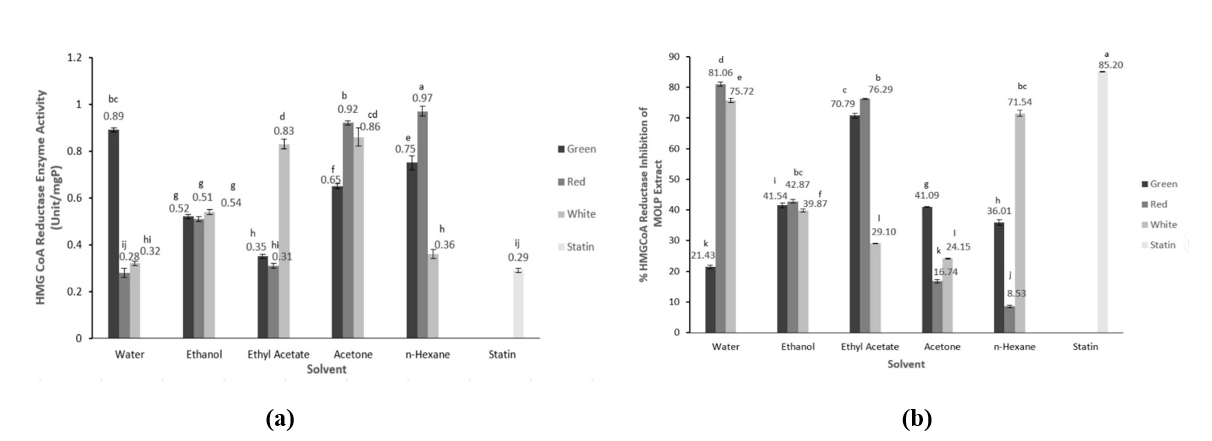
Figure 2 HMG-CoA reductase enzyme activity (a) and % inhibition and (b) of the extracts MOLP (Moringa oleifera leaves powder) with different leaf stalk colors.
Different leaf stalk colors exhibited different HMG-CoA reductase activity significantly (p < 0.05) (Figure 2(b)). The average activity of MOLP from white leaf stalk was 0.58 0.02 unit/mg P, the red was 0.60 0.02 unit/mg P, and the green was 0.64 0.01 unit/mg P. The statin as a control had an enzyme activity of 0.25 0.01 unit/mg P. The extraction solvents affected the HMG-CoA reductase activity. The extracts with different extraction solvents also revealed significantly different HMG-CoA reductase activity (p < 0.05). The lowest activity was found in the extraction solvent of water, and followed in sequence by ethyl acetate, ethanol, hexane, and acetone.
The HMG-CoA reductase activities are inversely proportional their enzyme inhibitory activities. Lower residual enzyme activity means higher inhibitory activity, thus the ability to inhibit cholesterol synthesis or cholesterol reduction is also higher. Regardless of the extraction solvent (average of 5 different extraction solvents), the highest HMG-CoA reductase inhibitory activity was obtained from the MOLP extract with white leaf stalk (48.08 ± 0.01%), followed by red (45.10 ± 0.01%) and green (42.72 ± 0.01%). HMG-CoA reductase inhibition by statins showed an inhibition of 85.20%. The highest inhibition was found in the aqueous extract of MOLP from red leaf stalk with an inhibitory activity of 81.06%, and the lowest inhibitory activity was observed in the hexane extract of MOLP from red leaf stalk at 8.53%. Different extraction solvents with different polarities may have different bioactive compound profiles, resulting in different inhibitory activities. For every MOLP from different leaf stalk colors and extraction solvent, the sequence of the HMG -CoA reductase activity from the highest was the water extract of MOLP from red leaf stalk (81.06%), followed by ethyl acetate extract of MOLP from green leaf stalk (77.30%), water extract of MOLP from white leaf stalk (75.72%). The water extract of MOLP from red stalk exhibited high HMG-CoA reductase activity, which is almost comparable to the statin drug.
A previous in vivo studies [25] showed that ethanol extract of MO significantly suppressed HMG-CoA reductase activity with oral administration of 200 and 400 mg/kg body weight for one month. Jain et al. [37] revealed that methanol extract of MO leaves reduced HMG-CoA reductase activity. Reddy et al. [12] showed that polyphenol extract from MO leaves inhibited HMG-CoA reductase activity. The mechanism of polyphenol inhibition is through binding to the active site of HMG-CoA reductase and blocking the binding of nicotinamide adenine dinucleotide phosphate (NADP+), including kaempferol but with low affinity [38].
Inhibition of pancreatic lipase by MOLP extracts from different leaf stalk colors
MOLP extracts with different extraction solvent polarities showed different abilities in inhibiting pancreatic lipase (Figure 3(b)). Statistically, extraction solvents and MOLP from different leaf stalk colors significantly affected pancreatic lipase activity (p < 0.05). The highest lipase activity inhibition was found in the ethyl acetate extract of MOLP from red leaf stalk (81.01%). The lowest inhibition was the acetone extract of MOLP with white leaf stalk (7.63%). In comparison, orlistat showed an inhibitory activity of 85.21%. These findings indicate that MOLP with different leaf stalk colors have different abilities in inhibiting pancreatic lipase, even though extraction used similar extraction solvents. The highest lipase activity inhibition of MOLP extract with green leaf stalk was the ethyl acetate extract and the n-hexane extract for MOLP with white leaf stalk.
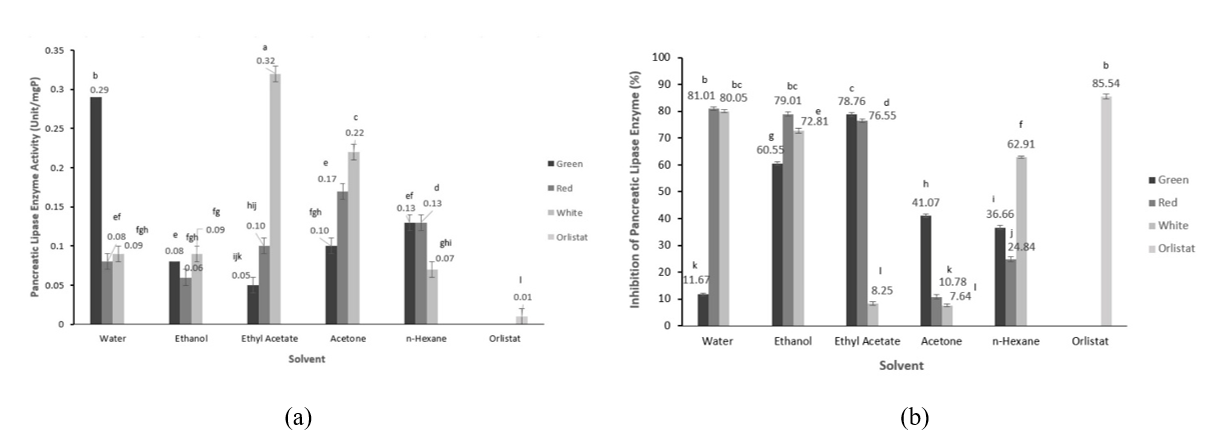
Figure 3 Pancreatic lipase enzyme activity (a) and % inhibition and (b) of the extracts MOLP (Moringa oleifera leaves powder) with different leaf stalk colors.
Despite extraction solvent, the leaf stalk colors affected the lipase inhibitory activity significantly (p < 0.05). The highest inhibition activity was exhibited by MOLP with red leaf stalk (54.44 ± 0.01%), followed by white (46.33 ± 0.01%) and green (45.74 ± 0.01%) leaf stalk. All extracts exhibited lipase inhibitory activity, indicating that multiple compounds in MOLP are responsible for this biological activity. Regardless of leaf stalk color, the highest lipase inhibition was observed in the ethanol extract, followed by the extraction solvents ethanol > ethyl acetate > hexane > acetone. It appears that solvent polarity results in different phytochemical polarities, which affect lipase inhibitory activity. Polar compounds may play a role in lipase inhibition. This finding is almost similar to the inhibition of HMG-CoA reductase activity where polar solvents, meaning polar bioactive compounds, show the highest inhibitory activity.
Some studies reported the lipase inhibition activity of MO leaf extract. Ogundipe et al. [7] reported that MO leaf ethanolic extracts inhibited the α-lipase enzyme in a concentration-dependent manner. The highest inhibitory activity was 93.84% at an extract concentration of 4.68 μg/mL, and the lowest was 79% at a concentration of 300 μg/mL. Pancreatic lipase plays a key role in fat metabolism; thus, inhibition of this enzyme reduces lipid hydrolysis, decreasing lipid absorption and thereby reducing fat accumulation that leads to obesity [38,39]. In this study, lipase inhibition was suggested to increase blood triglyceride levels. The findings in this study also suggest that MOLP has potential as an anti-obesity food ingredient.
Phytochemicals of MOLP from different leaf stalk colors
Phytochemical analysis of the extracts was performed with extracts that had the highest lipase or HMG-CoA reductase inhibitory activity. The extracts used were aqueous extracts from MOLP with red and white leaf stalk because they had the highest lipase and HMG-CoA reductase inhibitory activity. Meanwhile, ethyl acetate extract from MOLP with green leaf stalk showed the highest lipase and HMG-CoA reductase inhibition. Ethanol extracts were used for MOLP extracts from all different leaf stalk colors, because ethanol is usually used as an extraction solvent for bioactive compounds. The phytochemical profiles of the extracts are shown in Table S1.
The metabolite profile of MO leaves depends on genotype, cultivation method [40], and agroclimate [41]. The presence of phytochemical in MO leaves is greatly influenced by geography, variety and environmental conditions as well as season [17]. The metabolite profile of MO leaves is influenced by cultivation methods, harvest time and genotype [40]. In this study, the main factor affecting the different phytochemical profile was the MO genotypes expressed by different leaves stalk colors.
The number of identified phytochemicals from ethanol extracts was influenced by leaf stalk color (Table S1). The MOLP extract from green leaf stalk had 124 phytochemicals, followed by the white ones (100 compounds), and the red ones (95 compounds) (Table S1). The extraction solvent affected the extracted compounds for MOLP from similar leaf stalk colors, where the ethyl acetate extract showed higher identified compounds (135 phytochemicals) than the ethanol extract of MOLP from green leaf stalk. Meanwhile, the water extracts showed fewer compounds from MOLP from red (85 compounds) and white (68 compounds) leaf stalk compared to the corresponding ethanol extracts.
Flavonoids were the predominant phytochemicals for all MOLP extracts from different leaf stalk colors. Ethanol extracts had a higher number of flavonoids than water extracts in MOLP from red and white stalk colors. However, the identified flavonoid compounds of the ethyl acetate extract and ethanol extract were similar in the MOLP from the green leaf stalk. Flavonoids were dominated by quercetin, kaempferol, and their derivatives. This finding is in accordance with our previous study on the ethanolic extract of blanched MOLP [42]. but the extracts from this study showed a greater number of flavonoid compounds. This previous study evaluated the effect of blanching pH [42] which affected the occurrence of phytochemicals. Rutin was also found in the extract at (Table S1). Rutin was reported to strongly bound to HMG-CoA reductase and decreased its activity [43]. whereas quercetin was reported to moderately inhibit HMG-CoA reductase [44]. Kaempferol was reported to inhibit pancreatic lipase and had a synergistic effect with orlistat [45]. In addition, quercetin was also a promising pancreatic lipase inhibitor [46]. Rutin (quercetin-3-O-rutinoside) substantially decreased pancreatic lipase activities with IC50 value of 35 μg/mL [36]. The MOLP extract contained some typical MO leaf phytochemical such as glucosinolates and alkaloid Marumoside A and B (Table S1). These compounds were also found in the ethanolic extract of blanched MOLP [42]. Sheu et al. [47] reported that glucosinolates extract from Brassica juncea reduced HMG-CoA reductase activity. The ethanol and ethyl acetate extracts of MOLP contained sterols, comprising of β-sitosterol and campesterol. MO leaves contained phytosterols [48], which can improve lipid and apo B profiles [40], modulate the expression of transporters, LDL receptors, and inhibit HMG-CoA reductase.
Data in Figure 2 reveal that the highest HMG-CoA reductase inhibitory activity for each MOLP with different leaf stalk colors is the water extract of MOLP from red leaf stalk > the ethyl acetate extract of MOLP from green leaf stalk > the water extract of MOLP from green leaf stalk. Regardless of the lowest number of identified phytochemicals, MOLP from red leaf stalk showed the highest HMG-CoA reductase inhibition. The identified compounds at Table S1without considering their concentrations. A previous study [22]. showed that the total phenolic and flavonoid contents of MOLP from the red leaf stalk are the highest compared to the green and white ones. Meanwhile, the pancreatic lipase inhibitory activities were the water extract of MOLP from red leaf stalk > the water extract of MOLP from green leaf stalk > the ethyl acetate extract of MOLP from green leaf stalk. This data is also in contrary to the number of identified phytochemicals for each extract, meaning that the inhibitory activity was not only determined by the total identified compounds but also by their concentrations.
Lipid profile changes and fecal cholesterol
Figure 4 shows the lipid profile changes during MOLP administration to the hypercholesterolemia rats compared to the normal rats with standard diet and hypocholesterolemia rats without MOLP feeding. Table 2 shows the lipid profile changes of each group after eight-week treatment. MOLP with different leaf stalk colors reduced LDL-c. TG, and TC, but increased HDL-c significantly (p < 0.05). Meanwhile, the positive control of hypocholesterolemia group without treatment remained having high TC, TG, and LDL-c; and low HDL-c. The lipid profile of rats group treated with simvastatin was almost similar with the changes of MOLP treated group.
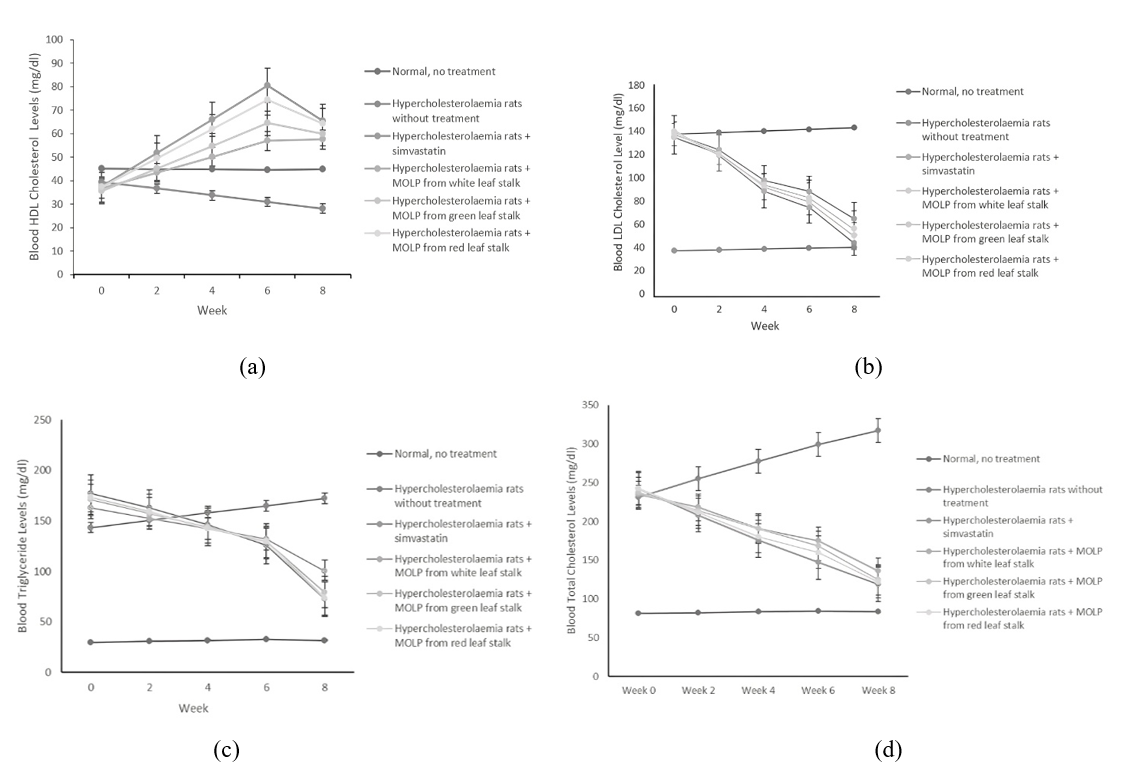
Figure 4 Blood serum lipid profiles changes during eight-week treatment. (a) HDL-c, (b) LDL-c, (c) triglycerides, (d) total cholesterol.
Table 2 Lipid profile and fecal cholesterol changes of groups of hyperglycemia rats treated with Moringa oleifera leaf powder (MOLP) from different leaf stalk colors.
|
Normal Without treatment (negative control) |
Hypercholesterolemia without treatment (positive control) |
Hypercholesterolemia + simvastatin |
Hypercholesterolemia + MOLP from white leaf stalk |
Hypercholesterolemia + MOLP from green leaf stalk |
Hypercholesterolemia + MOLP from red leaf stalk |
Serum Cholesterol Levels (mg/dL) |
|
|
|
|
|
|
Initial - Week 0 |
81.11 ± 0.14 |
231.38 ± 0.11 |
242.45 ± 0.01 |
234.64 ± 0.04 |
237.35 ±0.06 |
242.35 ± 0.12 |
Final - Week 8 |
83.19 ± 0.08 |
317.4 ± 0.05 |
118.67 ± 0.01 |
135.62 ± 0.04 |
124.58 ±0.06 |
121.67 ± 0.10 |
Mean of % Change |
+2.56 f |
+37.18 a |
− 51.05 e |
−42.20 b |
−47.51 c |
−49.8 d |
Fecal Cholesterol Level (mg/dL) (mg/100 g feces) |
|
|
|
|
|
|
Initial - Week 0 |
137.63 ± 0.01 |
79.95 ± 0.02 |
251.51 ± 0.01 |
158.14 ± 0.01 |
166.82 ± 0.02 |
190.98 ± 0.03 |
Final - Week 8 |
135.55 ± 0.05 |
69.2 ± 0.03 |
110.73 ± 0.01 |
78.12 ± 0.03 |
80.15 ± 0.01 |
90.31 ± 0.01 |
Mean of % Change |
−1.55 f |
−13.44 a |
+55.97 e |
+50.6 b |
+51.95 c |
+52.71 d |
Blood Triglyceride Level (mg/dL) |
|
|
|
|
|
|
Initial - Week 0 |
29.63 ± 0.07 |
143.27 ± 0.09 |
177.1 ± 0.07 |
163.13 ± 0.10 |
170.70 ±0.11 |
173.57 ± 0.12 |
Final - Week 8 |
31.72 ± 0.06 |
172.15 ± 0.09 |
73.22 ± 0.08 |
100.42 ± 0.08 |
79.56 ± 0.10 |
73.87 ± 0.08 |
Mean % Change |
+7.05 f |
+20.16 a |
−58.66 c |
−38.44 b |
−53.39 d |
−57.44 e |
Blood HDL Level (mg/dL) |
|
|
|
|
|
|
Initial - Week 0 |
45.13 ± 0.08 |
39.45 ± 0.06 |
37.47 ± 0.09 |
35.56 ± 0.53 |
35.56 ±0.07 |
37.21 ± 0.07 |
Final - Week 8 |
44.72 ± 0.09 |
28.09 ± 0.08 |
65.26 ± 0.05 |
57.44 ± 0.12 |
59.95 ± 0.08 |
64.36 ± 0.06 |
Mean of % Change |
−0.92 e |
−28.80 f |
+74.16 a |
+57.11 d |
+68.59 c |
+72.96 b |
LDL cholesterol (mg/dL) |
|
|
|
|
|
|
Initial - Week 0 |
30.04 ± 0.07 |
135.73 ± 0.06 |
131.82 ± 0.06 |
134.53 ± 0.06 |
138.66 ± 0.08 |
135.93 ± 0.1 |
Final - Week 8 |
32.13 ± 0.08 |
142.9 ± 0.06 |
38.77 ± 0.09 |
58.1 ± 0.05 |
48.72 ± 0.03 |
42.54 ± 0.07 |
Mean of % Change |
+6.95 f |
+5.28 a |
−70.59 e |
−56.81 b |
−64.86 c |
−68.7 d |
Explanation: (−) = decrease (+) = increase.
*Data is the average of % change for each rat. Numbers followed by different letters on the same line indicate there is a significant difference (α = 0.05).
This study strengthens the evidence of the hypolipidemic properties of MOLP. Most studies on this property used MO leaf extract such as reported by Chen et al. [8] who used ethanolic extract and examined the hypocholesterolemic property through pancreatic lipase inhibition assay and cell line for biochemical lipid metabolism analysis. The study by Ezzat et al. [25] revealed the hypocholesterolemic property of ethanolic MO leaf extract in obese rats indicated by a significant reduction of TC and LDL-c. This extract also showed HMG-CoA reductase inhibition. The study by Jain et al. [41] showed that the methanolic MO leaf extract administration along with hyperlipidemic diet for 30 days lowered the blood serum levels of TC, TG, LDL-c, VLDL-c, and atherogenic index. This hypolipidemic properties were attributed to the increase in fecal cholesterol excretion. More specifically, Reddy et al. [12] indicated the hypocholesterolemic activity of polyphenol extract of MO leaves through HMG-CoA reductase inhibition and fecal bile acid-binding.
Limited in vivo studies explored the MOLP administration to improve blood serum lipid profile. Helmy et al. [20] compared the MOLP and MO leaf extract to lower cholesterol in high- fat diet rats. The results showed that feeding MOLP at 0.74% and administration of dry MO leaf extract of 400 mg/kg bw/day inhibited the elevation of TC, TG, LDL-c, and malondialdehyde as the marker of oxidative stress. Jain et al. [37] evaluated the formulated MOLP at 100 and 200 mg/100 g bw doses for 28 days and found a significant TC decrease. Ariestiningsih et al. [49] showed that MOLP feeding at 500 mg/kg bw to prediabetic rats for weeks improved lipid profiles and reduced aortic thickness. Khan et al. [50] showed that MOLP force-fed into rabbits for 45 days improved the lipid profile. No studies reported a comparison of MOLP from different varieties on the lipid profile improvements.
MOLP have more advantages than MO leaf extract in modulating lipid profile because the powder composition might also affect lipid profile improvements, besides bioactive compounds. Fidyasari et al. [22] showed that MOLP from different leaf stalk colors had soluble dietary fiber of 9.54-11.61% and insoluble was 0.23-0.36%. Dietary fiber, mainly soluble fiber, possibly improves lipid profile by inhibiting lipid absorption and short-chain fatty acid production. Yang et al. [51] showed that MO leaf polysaccharide consisted of arabinose, rhamnose, galactose, and galacturonic acid and revealed strong effects on bile acid-binding and cholesterol micelle solubility inhibition thus reducing lipid digestion and absorption.
Fatty acids of MOLP are dominated by unsaturated fatty acids, and the predominant are linoleic and α linolenic acids for all MOLP from different leaf stalk colors. The fat content of MOLP is 3.81-4.04% [22]. Both fatty acids might also improve the lipid profile. A randomized trial by Nuotio et al. [53] showed that diets enriched with α linolenic or linoleic acids decreased serum lipoprotein(a) concentration by 7–10%, with a pronounced effect shown by alpha-linolenic acid. Dietary intervention with α linolenic acid improved lipid profiles by decreasing TC, TG, LDL-c, and VLDL-c levels [52].
Table 2 shows that the MOLP increased fecal cholesterol concentration indicating inhibition of cholesterol absorption by this powder. The fiber in the MOLP might be responsible for this inhibition. The soluble fiber can form a weak gel in the gastrointestinal track and entrap nutrition including fats and cholesterol thus hindering their absorption [53]. Yang et al. [51] postulated the role of MO leaf polysaccharides in the hypolipemic mechanism by binding bile acids through hydrogen bonds and hydrophobic interaction thus regulating cholesterol metabolism. Other hypotheses were bile acid binding or matrix/barrier formation to entrap bile acid micelles, and inhibition of cholesterol solubility in the micelle thus reducing its absorption. Dietary fiber of MO leaves were fermentable in vitro and produced short chain fatty acids, with the predominant propionic acid which was able to inhibit cholesterol synthesis [54].
The leaf stalk colors significantly affected (p<0.05) the changes of TC, TG, LDL-c, and HDL-c levels, in which HDL-c levels increased and TC, TG, and LDL-c decreased. These changes were comparable to a simvastatin-treated group, but the hypocholesterolemic group without MOLP treatment exhibited a contradictory phenomenon. The highest increase in HDL-c level was observed in MOLP with red leaf stalk and sequentially followed by green and white. In accordance, the highest decreases in TG, TC, and LDL-c levels are found in MOLP with red leaf stalk, followed in sequence by green and white.
The extracts of MO leaves have been studied widely and contains bioactive compounds responsible for lowering blood cholesterol levels. MOLP with different leaf stalk colors had a slightly different amount of total phenolic (TPC) and flavonoid content (TFC). The highest TPC and TFC were found in the MOLP from the red leaf stalk, and the lowest were reported in the MOLP from while leaf stalk [22]. Chen et al. [56] reported the bioactive playing the role for hypolipidemic properties were flavonoid glycosides of quercetin and kaempferol mono-glycoside. Kaempferol also has the ability to bind HMG-CoA reductase [57].
The highest dietary fiber was found in the MOLP with red leaf stalk, followed by green and white [22]. The fiber content in the MOLP contributed to the lipid profile improvement thus the best lipid profile repair was found in the rat group fed by MOLP from the red leaf stalk, followed by green and white. This fiber seemed to inhibit cholesterol absorption indicated by fecal cholesterol concentration with the highest in the feces from group of rats fed by MOLP with red leaf stalk, followed by MOLP with green and white leaf stalk.
The occurrence of linoleic and α linolenic acids in the MOLP might be attributed to the lipid profile improvement. The highest linoleic acid was found in the MOLP from red leaf stalk followed by green and white. Meanwhile, the highest α linolenic acid was found in the MOLP with green leaf stalk, followed by white and red. Thie contribution of fatty acids might not as high as the dietary fiber because their content was lower.
MOLP with different colors have different bioactive compound profile. All extracts, except ethanol extract of MOLP from red leaf stalk, contained chlorogenic acid (Table S1). Chlorogenic acid was found to reduce TC and TG levels in plasma of obese rats (rats fed a high-fat diet), thus showing anti-dyslipidemic activity [5]. Phenolic compounds reduce the atherogenic index, blood serum cholesterol levels, LDL-c, triglycerides, and VLDL = c, and increase HDL-c in hyperlipidemic rats [58].
Phytosterols are plant sterols that have a chemical structure similar to cholesterol. They work by competitively inhibiting cholesterol absorption, thereby lowering TC and LDL-c levels [21]. β-Sitosterol inhibits cholesterol absorption. Once phytosterols enter and are carried by enterocytes, phytosterol absorption is inhibited by the activity of efflux transporters, which contain a pair of ATP-binding proteins (ABC) secretes phytosterols and small amounts of cholesterol back into the intestinal lumen [59]. Plant sterols reduces absorption of cholesterol from gut by competing for limited space for cholesterol in mixed micelles and thereby lower serum concentration of cholesterol, despite compensatory increase in cholesterol synthesis in liver and other tissues.
Quercetin, another compound found in the MOLP extracts (Table S1), reduces cholesterol esterase activity [58]. Meanwhile, kaempferol induced the LDLR gene expression thus reducing atherosclerosis risk [43]. Other main typical phytochemicals founds in the extract (Table S1) that responsible for hypolipidemic properties of MO are niazirinin, niaziminin A, and niaziminin B [60]. The MOLP extracts also had glucosinolates, which in the gut were broken down into isothyocianate and this compound reduced LDL-c [61].
Interestingly, the group of rats fed by MOLP with white leaf stalk exhibited lower lipid profile improvement than those from MOLP with red and green leaf stalk. Table 1 shows that the amount of formulated MOLP was the highest quantity in the feed from MOLP with white leaf stalk. The different amounts of MOLP with red, green, and white leaf stalk were due to iso-protein and iso-calorie feed formulation. This fact indicated that not only quantity affected the lipid profile improvement but also the composition of MOLP and their bioactive compounds.
The ability of MOLP with different colors to inhibit HMG-CoA reductase and pancreatic lipase evaluated by their extract showed that the highest HMG-CoA reductase inhibition activity was the extract from MOLP with white leaf stalk, followed by red and green. Meanwhile, the highest lipase inhibition activity was exhibited by MOLP with red leaf stalk, followed by white and green leaf stalk. MOLP with red leaf stalk showed the best lipid profile improvement but did not exhibit the highest HMG-CoA reductase activity. This fact indicates that the mechanism of lipid profile improvement by MOLP comes from many factors, not only enzyme inhibitions, but also the role of dietary fiber, bioactive compounds, and fatty acid composition.
Conclusions
This study proves that the MOLP is more comprehensive in improving blood lipid profile than the extracts due to the synergistic effects of its components such as fiber, fatty acids, and bioactive compounds. Different leaf stalk colors showed different abilities to inhibit HMG-CoA reductase and pancreatic lipase activities. The ability to inhibit pancreatic lipase activity gives an insight for the potential of MOLP as anti-obesity food ingredient. The ability to improve lipid profile in in vivo experiment revealed that the MOLP form the red leaf stalk is the best followed by white and green ones. The variability of phytochemicals in the extract is not well correlated to HMG-CoA and pancreatic lipase inhibitory activities, indicating that the concentrations of phytochemicals have a contribution. MOLP from different leaf stalk colors showed different HMG-CoA reducatse and pancreatic lipase inhibitory activities and lipid profile improvement, meaning that the stalk colors should be considered in preparing MOLP as functional food ingredient especially for cholesterol-lowering food products.
Acknowledgements
The authors are very grateful to Universitas Brawijaya, Indonesia for funding this study through Hibah Guru Besar Grant Batch II Year 2024 Contract No. 02992/UN.F1001/B/KS/2024 and Mr. Hosman for providing the Moringa oleifera leaf powder.
Declaration of Generative AI in Scientific Writing
Generative AI tools, were used solely to enhance the language and readability of this manuscript. All content has been reviewed and verified by the authors to ensure its accuracy and integrity. The use of such AI tools was conducted under human oversight and does not affect the authors' full responsibility for the content. No AI tool was listed or considered as an author or co-author of this work.
CRediT Author Statement
Lina Novi Ariani: Methodology, Formal Analysis, Investigation, Visualisation, Writing - Original Draft. Teti Estiasih: Conceptualization, Data Curation, Methodology, Project Acquisition, Validation, Visualization, Supervision, Writing - original draft, Writing-review and revision. Wenny Bekti Sunarharum: Supervision. Alfi Khatib: Conceptualization, Supervision. Sentot Joko Raharjo: Data Curation, Validation. Ambar Fidyasari: Visualization, Writing-original draft, Writing-review and revision.
[1] NZA Rani, K Husain and E Kumolosasi. Moringa genus: A review of phytochemistry and pharmacology. Frontiers in Pharmacology 2018; 9(2), 108.
[2] P Vijay, M Tamilselvi and R Mohankumar. Isolation, identification and HPLC analysis of a phytochemical from Moringa oleifera leaves. Materials Today: Proceedings 2023; 93(1), 86-90.
[3] W Khan, R Parveen, K Chester, S Parveen and S Ahmad. Hypoglycemic potential of aqueous extract of Moringa oleifera leaf and in vivo GC–MS metabolomics. Frontiers in Pharmacology 2017; 8(9), 77.
[4] EE Amina, JO Adisa, SM Gamde, EB Omoruyi, HM Kwaambwa and LM Mwapagha. Hypoglycemic assessment of aqueous leaf extract of Moringa oleifera on diabetic Wistar rats. Biochemistry Research International 2024; 2024, 9779021.
[5] UF Magaji, O Sacan and R Yanardag. Alpha-amylase, alpha-glucosidase and glycation inhibitory activity of Moringa oleifera extracts. South African Journal of Botany 2020; 128(1), 225-230.
[6] GI Olasehinde, O Ojurongbe, AO Adeyeba, OE Fagade, N Valecha, IO Ayanda, AA Ajayi and LO Egwari. In vitro studies on the sensitivity pattern of Plasmodium falciparum to antimalarial drugs and local herbal extracts. Malaria Journal 2014; 13(2), 63.
[7] A Ogundipe, B Adetuyi, F Iheagwam, K Adefoyeke, J Olugbuyiro and O Ogunlana. In vitro experimental assessment of ethanolic extract of Moringa oleifera leaves as an α- amylase and α-lipase inhibitor. Biochemistry Research International 2022; 2022, 4613109.
[8] GL Chen, YB Xu, JL Wu, N Li and MQ Guo. Hypoglycemic and hypolipidemic effects of Moringa oleifera leaves and their functional chemical constituents. Food Chemistry 2020; 333, 127478.
[9] Z Hong, J Xie, H Hu, Y Bai, X Hu, T Li, J Chen, J Sheng and Y Tian. Hypoglycemic effect of Moringa oleifera leaf extract and its mechanism prediction based on network pharmacology. Journal of Future Foods 2023; 3(4), 383-391.
[10] SJ Melebary and MHR Elnaggar. Impact of Moringa oleifera leaf extract in reducing the effect of lead acetate toxicity in mice. Saudi Journal of Biological Sciences 2022; 30(1), 103507.
[11] GM Kodi, HA Mustafa, A Abd and A Idris. The effects of Moringa oleifera leaves on complete blood count, renal and liver functions as potential therapy for malnutrition. Molecular and Cellular Biomedical Sciences 2022; 6(2), 55-62.
[12] PV Reddy, AU Asna Urooj, SS Sairam, FAF Ahmed and NN Prasad. Hypocholesterolemic effect of Moringa oleifera polyphenols in rats fed high-fat cholesterol diet. Malaysian Journal of Nutrition 2017; 23(2), 473-478.
[13] D Belete, BM Jacobs, C Simonet, JP Bestwick, S Waters, CR Marshall, R Dobson and AJ Noyce. Association between antiepileptic drugs and incident Parkinson disease. JAMA Neurology 2023; 80(2), 183-187.
[14] J Al-Zahrani, MM Shubair, S Al-Ghamdi, AA Alrasheed, AA Alduraywish, FS Alreshidi, SM Alshahrani, M Alsalamah, BF Al-Khateeb, AI Ashathri, A El-Metwally and KK Aldossari. Prevalence of hypercholesterolemia and associated risk factors in Al-Kharj population, Saudi Arabia. BMC Cardiovascular Disorders 2021; 21(1), 22.
[15] S Ghasi, E Nwobodo and JO Ofili. Hypocholesterolemic effects of crude leaf extract of Moringa oleifera in high-fat diet-fed Wistar rats. Journal of Ethnopharmacology 2000; 69(1), 21-25.
[16] L Sissoko, N Diarra, I Nientao, B Stuart, A Togola, D Diallo and ML Willcox. Moringa oleifera leaf powder for type 2 diabetes: A pilot clinical trial. African Journal of Traditional, Complementary and Alternative Medicines 2020; 17(2), 29-36.
[17] ZF Ma, J Ahmad, H Zhang, I Khan and S Muhammad. Evaluation of phytochemical and medicinal properties of Moringa oleifera as a potential functional food. South African Journal of Botany 2020; 129(3), 40-46.
[18] ML Cuellar-Núñez, EG de Mejia and G Loarca-Piña. Moringa oleifera leaves alleviate inflammation through downregulation of IL-2, IL-6 and TNF-α. Food Research International 2021; 144(6), 110318.
[19] I Asrifah, T Estiasih and H Sujuti. Potential hypocholesterolemic activity of flour from Moringa oleifera leaves. Malaysian Journal of Nutrition 2017; 23(3), 449-461.
[20] SA Helmy, NFS Morsy, SM Elaby and MAA Ghaly. Hypolipidemic effect of Moringa oleifera leaf powder and extract in hypercholesterolemic rats. Journal of Medicinal Food 2017; 20(8), 755-762.
[21] J Wang, Y Du, L Jiang, J Li, B Yu, C Ren, T Yan, Y Jia and B He. LC-MS/MS-based chemical profiling of water extracts of Moringa oleifera leaves and pharmacokinetics of their major constituents in rat plasma. Food Chemistry: X 2024; 23(10), 101585.
[22] A Fidyasari, T Estiasih, SN Wulan and A Khatib. The physicochemical, functional, and pasting properties of Moringa oleifera leaf powder from different leaf stalk colors. CyTA-Journal of Food 2024; 22(1), 2402062.
[23] AO Ademosun, G Oboh, S Passamonti, F Tramer, L Ziberna, AA Boligon and ML Athayde. Phenolics from grapefruit peels inhibit HMG-CoA reductase and angiotensin-I converting enzyme and show antioxidative properties in endothelial EA.Hy 926 cells. Food Science and Human Wellness 2015; 4(2), 80-85.
[24] AA Redha, S Perna, A Riva, G Petrangolini, G Peroni, M Nichetti, G Iannello and M Naso. Novel insights on anti-obesity potential of the miracle tree, Moringa oleifera: A systematic review. Journal of Functional Foods 2021; 84(9), 104600.
[25] SM Ezzat, MH El Bishbishy, NM Aborehab, MM Salama, A Hasheesh, AA Motaal, H Rashad and FM Metwally. Upregulation of MC4R and PPAR-α expression mediates the anti-obesity activity of Moringa oleifera Lam. In high-fat diet-induced obesity in rats. Journal of Ethnopharmacology 2020; 251(4), 112541.
[26] GM Swamy and B Meriga. Therapeutic effect of Moringa oleifera leaf extracts on oxidative stress and key metabolic enzymes related to obesity. International Journal of Pharmaceutical Sciences and Research 2020; 11(4), 1949-1957.
[27] SJ Stohs and MJ Hartman. Review of the safety and efficacy of Moringa oleifera. Phytotherapy Research 2015; 29(6), 796-804.
[28] P Maitreesophone, HEE Khine, JQL Nealiga, V Kongkatitham, P Panuthai, C Chaotham, K Likhitwitayawuid and B Sritularak. α-Glucosidase and pancreatic lipase inhibitory effects and anti-adipogenic activity of endrofalconerol B, a bisbibenzyl from Dendrobium harveyanum. South African Journal of Botany 2022; 146(5), 187-195.
[29] A Grover, HS Rehan, LK Gupta and M Yadav. Correlation of compliance to statin therapy with lipid profile and serum HMG-CoA reductase levels in dyslipidemic patients. Indian Heart Journal 2017; 69(1), 6-10.
[30] X Yong-Bing, C Gui-Lin and G Ming-Quan. Antioxidant and anti-inflammatory activities of crude extracts of Moringa oleifera from Kenya and their correlations with flavonoids. Antioxidants 2019; 8(8), 296.
[31] ZW Teclegeorgish, YM Aphane, NS Mokgalaka, P Steenkamp and VJ Tembu. Nutrients, secondary metabolites and antioxidant activity of Moringa oleifera leaves and Moringa-based commercial products. South African Journal of Botany 2021; 142(11), 409-420.
[32] PG Reeves, FH Nielsen and GC Fahey. AIN-93 purified diets for laboratory rodents: Final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. Journal of Nutrition 1993; 123(11), 1939-1951.
[33] P Hasimun, EY Sukandar, IK Adnyana and DH Tjahjono. A simple method for screening antihyperlipidemic agents. International Journal of Pharmacology 2011; 7(1), 74-78.
[34] TTC Yang and MWL Koo. Hypocholesterolemic effects of Chinese tea. Pharmacological Research 1997; 35(6), 505-512.
[35] V Rodríguez-Sureda and J Peinado-Onsurbe. A procedure for measuring triacylglyceride and cholesterol content using a small amount of tissue. Analytical Biochemistry 2005; 343(2), 277-282.
[36] MS Ganjayi, RS Karunakaran, S Gandham and B Meriga. Quercetin-3-O-rutinoside from Moringa oleifera downregulates adipogenesis and lipid accumulation and improves glucose uptake via activation of AMPK/GLUT-4 in 3T3-L1 cells. Revista Brasileira de Farmacognosia 2023; 33(2), 334-344.
[37] PG Jain and RC Patel. Hypolipidemic activity of Moringa oleifera Lam. (Moringaceae) on high-fat diet-induced hyperlipidemia in albino rats. Revista Brasileira de Farmacognosia 2010; 20(6), 969-973.
[38] H Hasim, F Tunnisa, DN Faridah, S Saraswati and F Slameut. Antioxidant and lipase inhibitory potential and UHPLC profile of different fractions of Moringa oleifera leaves. Tropical Journal of Natural Product Research 2025; 9(1), 31-36.
[39] F Braham, DO Carvalho, CMR Almeida, F Zaidi, JMCS Magalhães, LF Guido and MP Gonçalves. Online HPLC-DPPH screening method for evaluation of radical scavenging phenols extracted from Moringa oleifera leaves. South African Journal of Botany 2020; 129(3), 146-154.
[40] LR Managa, ESD Toit and G Prinsloo. Variations in the leaf metabolite profile between hydroponic and field grown Moringa oleifera Lam. genotypes. Biochemical Systematics and Ecology 2021; 97, 104302.
[41] DK Mahato, A Kumar, M Devi, R Pandhi, B Sharma, S Pandey, H Pathak, K Ahmed, S Sharma and J Das. Ethnopharmacological properties and nutraceutical potential of Moringa oleifera. Phytomedicine Plus 2022; 2(1), 100168.
[42] T Estiasih, R Hidayat, RN Putri, K Ahmadi, DY Ali and RA Laeliocattleya. Moringa oleifera leaf powder colour improvement and lipoxygenase inactivation by different pHs and water-blanching times. CyTA-Journal of Food 2025; 23(1), 2586311.
[43] N Hanis, NA Ismail and EZ Ali. Systematic review on effectiveness of flavonoids against hypercholesterolemia: Insights from in-silico, in-vitro, and in-vivo studies. Food Chemistry Advances 2025; 7, 100981.
[44] K Pathak, J Das, A Das, R Saikia, JJ Sahariah, D Deb, P Das, S Gogoi, MP Pathak and H Pathak. Exploring the therapeutic potential of Garcinia cowa Roxb. for type 2 diabetes mellitus management: A network pharmacology approach. In Silico Research in Biomedicine 2025; 1, 100103.
[45] S Li, J Pan, X Hu, Y Zhang, D Gong and G Zhang. Kaempferol inhibits the activity of pancreatic lipase and its synergistic effect with orlistat. Journal of Functional Foods 2020; 72, 104041.
[46] JF Zhou, WJ Wang, ZP Yin, GD Zheng, JG Chen, JE Li, LL Chen and QF Zhang. Quercetin is a promising pancreatic lipase inhibitor in reducing fat absorption in vivo. Food Bioscience 2021; 43, 101248.
[47] M Sheu, MC Yeh, MC Tsai, CC Wang, YL Chang, CJ Wang and HP Huang. Glucosinolates extracts from Brassica juncea ameliorate HFD-induced non-alcoholic steatohepatitis. Nutrients 2023; 15(16), 3497.
[48] TT Liu, XT Liu, QX Chen and Y Shi. Lipase inhibitors for obesity: A review. Biomedicine & Pharmacotherapy 2020; 128(8), 110314.
[49] AD Ariestiningsih, AR Cempaka, I Kusumastuty, A Cahyaningrum, S Setiawati, D Arintya, U Azmi, P Salwa, H Malahayati, RA Wibowo and D Handayani. Moringa leaf powder improves lipid profiles and aortic thickness in Wistar rat model of prediabetes mellitus. Amerta Nutrition 2024; 8(2), 278-289.
[50] K Khan, GM Dahri, Y Shaikh, N Irum, ZY Memon, R Malik and MS Rajput. Effects of Moringa oleifera leaf powder on lipid parameters of adult male rabbits. Professional Medical Journal 2024; 31(6), 925-931.
[51] Y Yang, L Lin, M Zhao and X Yang. Hypoglycemic and hypolipemic potentials of Moringa oleifera leaf polysaccharide and polysaccharide-flavonoid complex. International Journal of Biological Macromolecules 2022; 210(6), 518-529.
[52] P Nuotio, MA Lankinen, T Meuronen, VD de Mello, T Sallinen, KA Virtanen, J Pihlajamäki, M Laakso and U Schwab. Dietary n-3 alpha-linolenic and n-6 linoleic acids modestly lower serum lipoprotein(a) concentration but differentially influence other atherogenic lipoprotein traits: A randomized trial. Atherosclerosis 2024; 395(8), 117562.
[53] K Khan, GM Dahri, Y Shaikh, N Irum, ZY Memon, R Malik and MS Rajput. Effects of Moringa oleifera leaf powder on lipid parameters of adult male rabbits. The Professional Medical Journal 2024; 31(6), 925-931.
[54] H Yue, B Qiu, M Jia, W Liu, X Guo, N Li, Z Xu, F Du, T Xu and D Li. Effects of α-linolenic acid intake on blood lipid profiles: A systematic review and meta-analysis of randomized controlled trials. Critical Reviews in Food Science and Nutrition 2020; 61(17), 2894-2910.
[55] LA Alahmari. Dietary fiber influence on overall health, with an emphasis on cardiovascular disease, diabetes, obesity, colon cancer and inflammation. Frontiers in Nutrition 2024; 11, 1510564.
[56] S Chen, X Wang, Y Cheng, H Gao and X Chen. Classification, biosynthesis, biological activities and application potential of flavonoids. Molecules 2023; 28(13), 4982.
[57] Z Wang, KL Lam, J Hu, S Ge, A Zhou, B Zheng, S Zeng and S Lin. Chlorogenic acid alleviates obesity and modulates gut microbiota in high-fat-fed mice. Food Science & Nutrition 2019; 7(2), 579-588.
[58] TFSN Soares, AVC da Silva and EN Muniz. Moringa leaf extract: A cost-effective and sustainable product to improve plant growth. South African Journal of Botany 2021; 141, 171-176.
[59] RK Saini, I Sivanesan and YS Keum. Phytochemicals of Moringa oleifera: A review of their nutritional, therapeutic and industrial significance. 3 Biotech 2016; 6(2), 203.
[60] E Camilleri and R Blundell. A comprehensive review of the phytochemicals, health benefits, pharmacological safety and medicinal prospects of Moringa oleifera. Heliyon 2024; 10(6), e27807.
[61] C Armah, C Derdemezis, MH Traka, JR Dainty, JF Doleman, S Saha, W Leung, JF Potter, JA Lovegrove and RF Mithen. Diet rich in high glucoraphanin broccoli reduces plasma LDL cholesterol: Evidence from randomised controlled trials. Molecular Nutrition & Food Research 2015; 59(5), 918-926.