Trends
Sci.
2026;
23(8):
12785
Protective Effects of Salacca zalacca Peel Extract on Oxidative and Inflammatory Liver Injury Biomarkers in CCl4-Induced Rats
Danis
Pertiwi1,*,
Maritsatun Nisa2,
![]() and Laili Fitria Zulfa3,
and Laili Fitria Zulfa3,
![]()
1Department of Clinical Pathology, Faculty of Medicine, Universitas Islam Sultan Agung, Semarang 50112, Indonesia
2Faculty of Medicine, Universitas Islam Sultan Agung, Semarang 50112, Indonesia
3Faculty of Biology, Universitas Gadjah Mada, Yogyakarta 55281, Indonesia
(*Corresponding author’s e-mail: [email protected])
Received: 24 November 2025, Revised: 15 December 2025, Accepted: 22 December 2025, Published: 10 March 2026
Salacca zalacca peel extract (SPE) contains phenolic and flavonoid compounds with reported antioxidant and anti-inflammatory properties, but in vivo evidence from liver injury models remains limited. This study evaluated the effects of SPE on biomarkers in CCl₄-induced liver injury in Wistar rats by measuring serum alanine aminotransferase (SGPT), TNF-α, and superoxide dismutase (SOD). Thirty rats were divided into 5 groups: Healthy controls, 2% CCl₄-treated controls (C-), 3.6 mg/kg NAC-treated controls (C+), and CCl₄-induced rats treated with low (400 mg/kg BW/day) and high (600 mg/kg BW/day) doses of SPE for 14 days. SPE was administered orally following CCl₄ induction. SGPT levels were measured by spectrophotometry, and TNF-α and SOD levels were analyzed using ELISA. Compared to CCl₄ controls, SPE groups showed significantly reduced SGPT and TNF-α levels and elevated SOD activity (p < 0.005), indicating improved markers of hepatocellular injury, inflammation, and antioxidant status. The high-dose SPE group exhibited the greatest effect. In conclusion, SPE administration demonstrated antioxidant and anti-inflammatory benefits by lowering SGPT and TNF-α while increasing SOD levels.
Keywords: Salacca zalacca peel extract, CCl₄, SGPT, TNF-α, SOD, Hepatoprotection
Introduction
The liver is essential for metabolism and detoxification, with high susceptibility to oxidative stress and inflammation caused by toxins and drugs [1]. Previous studies have shown that various liver injuries are often due to infection, alcohol, or chemicals, serving as major health challenges. Among experimental models, carbon tetrachloride (CCl₄) is widely used to investigate acute liver injury [2,3]. Metabolism of CCl₄ by CYP2E1 produces free radicals and ROS, causing oxidative stress, cellular damage, and activating NF-κB along with high TNF-α levels [4,5]. It also raises serum alanine aminotransferase (SGPT), TNF-α, and lowers superoxide dismutase (SOD) activity, serving as a useful tool for testing hepatoprotective agents [6,7]. Although N-acetylcysteine (NAC) has been widely used as a synthetic hepatoprotective to replenish glutathione and scavenge ROS, the application is associated with
inconsistent results, low bioavailability, a short half-life, and side effects [8]. These challenges have increased research interest in plant-derived antioxidants, including flavonoids, which have been widely reported to attenuate oxidative processes in various experimental settings [3,9,10].
Snake fruit (Salacca zalacca) peel, a by-product rich in flavonoids and phenolic acids [11,12], contains compounds such as quercetin [13], ferulic acid [14], chlorogenic acid [15], rutin [16], and catechins [17]. These have shown antioxidant and anti-inflammatory properties in previous research, protecting cellular and animal models of oxidative stress. Specific studies highlight roles for quercetin and chlorogenic acid in modulating oxidative and inflammatory responses. Ethanolic extracts of the peel exhibit strong antioxidant activity, and both in vivo and in vitro studies report its protective effects in neurotoxicity and metabolic stress, including reduced oxidative markers and inflammatory mediators. Thus, the phytochemical profile of S. zalacca peel warrants further study in oxidative and inflammatory conditions, such as CCl₄-induced liver injury [12,18,19]; Gallic acid Peel [20], Caffeic acid, Ferulic acid, Chlorogenic acid, Quercetin, Rosmarinic acid, Gallic acid Peel [20], Caffeic acid, Ferulic acid, Chlorogenic acid, Quercetin, Rosmarinic acid.
Despite the high content of flavonoids and phenolic acids, in vivo studies on the effects of S. zalacca peel extract, particularly in toxin-induced liver injury models, remain limited. Most available studies have focused on its antioxidant capacity or evaluated isolated compounds rather than whole-extract effects in CCl₄-induced injury [21-23]. There are limited data on the effect on biomarkers such as SGPT, SOD, and TNF-α [15]. Therefore, this study aimed to evaluate the effects of SPE extract on these biomarkers in Wistar rats with CCl₄-induced acute liver injury, providing in vivo evidence for its hepatoprotective potential [24,25].
Ethical statement
All animal procedures complied with relevant ethical guidelines and were approved by the Bioethics Commission of Universitas Sultan Agung Semarang (Ethical Approval No. 153/III/2025). Animals were euthanized by CO₂ inhalation after blood collection [26-28]. A separate control group was used, as recovery from CCl₄-induced injury is unreliable within the study timeframe, rendering a within-subject design unsuitable [29].
Animal handling and experiment design
For
the animal model, male Wistar rats (Rattus
norvegicus),
aged 2 to 3 months and weighing between 180 and 200 g,
were obtained from the Universitas Gadjah Mada breeding unit.
Animals were acclimatized for 7 days under standard housing
conditions with ad libitum access to food and water. Thirty rats
were randomly assigned to 5 groups (n = 6):
P1 (Healthy
control): standard diet and water; P2 (negative
control): 2%
CCl₄ at 0.2 mL/10 g BW orally for 14 days; P3 (positive
control): CCl₄
followed 3 h later by oral NAC (3.6 mg/kg BW/day); P4: CCl₄
followed 3 h later by SPE 400 mg/kg BW/day; P5: CCl₄ followed
3 h later by SPE 600 mg/kg BW/day. The 3-hour interval was selected
based on previous protocols, allowing sufficient time for CCl₄
absorption while enabling evaluation of post-insult intervention
[30,31].
Analytical-grade NAC, CCl₄, and ethanol (96%) were obtained from Merck (Germany). ELISA kits for TNF-α, SOD, and SGPT were obtained from MyBioSource® (Wuhan, China). High-concentration ethanol (96%) was used as an extraction solvent because previous phytochemical studies report its suitability for efficiently extracting phenolic and flavonoid constituents from tropical plant materials such as S. zalacca peel [32,33].
Preparation of Salacca zalacca peel extract
Fresh S. zalacca var. pondoh peels were collected from Nglumut, Magelang, and authenticated at Universitas Islam Sultan Agung. Peels were sorted, cut, and shade-dried in a temperature-regulated drying cabinet (Memmert, Germany) at 50 - 60 °C under indirect heat to prevent degradation of thermo-sensitive compounds [34,35]. The temperature was controlled with a digital thermostat throughout the process. Dried peels were ground into powder, and 600 g was macerated in 96% ethanol (1:10 w/v) for 4 days at room temperature with intermittent agitation. The mixture was filtered and re-macerated twice under identical conditions to maximize extraction [36,37]. Combined filtrates were concentrated at 45 °C under reduced pressure using a rotary evaporator and stored at 2 - 8 °C. Extraction yield (%) was calculated as:
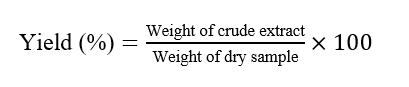
Dosage determination
The dosage of SPE (400 and 600 mg/kg BW/day) was selected based on previous in vivo studies employing similar dose ranges in antioxidant investigations. A 3.6 mg/kg BW/day NAC dose (200×0.018) was derived from the standard 200 mg human dose using body surface area conversion, to reflect a clinically relevant dose rather than high experimental dosing typically used in hepatoprotection models [38,39]. The CCl₄ dose (2% solution, 0.2 mL/10 g BW) was chosen based on established protocols that reliably induce acute liver injury in rats [9].
Preparation of serum from collected blood
Retro-orbital blood was collected from rats using a modified method. On day 15, approximately 3 mL of blood was collected from the retro-orbital plexus under chloroform inhalation anesthesia by a trained laboratory technician. This method was selected to obtain sufficient serum volume for all biochemical assays [39-41]. Blood samples were centrifuged at 2,000 rpm for 10 min, and the separated serum was aliquoted and stored at −80 °C until analysis.
Activity of serum alanine aminotransferase (ALT/SGPT)
Serum ALT/SGPT levels were measured using the spectrophotometric method [42]. The samples obtained were brought to room temperature and mixed for homogeneity. Reagent tablets were dissolved in buffer (1:10), then 50 µL serum was combined with 500 µL reagent, and the sample was incubated for 1 min. Absorbance at 340 nm was measured at 1-minute intervals for a total of 3 min using a Thermo Fisher spectrophotometer. The measurement of enzyme activity (U/L) was performed by observing the reduction in absorbance.
Activity of tumor necrosis factor-alpha (TNF-α)
The activity of TNF-α was assessed using an ELISA assay [43]. Samples were brought to room temperature and gently mixed to ensure uniformity before analysis. Subsequently, TNF-α was measured with MyBioSource® ELISA kits (MBS2600843) per the manufacturer’s guidelines. Samples were analyzed at 450 nm, and the results are in pg/mL (ratio scale).
Activity of superoxide dismutase (SOD)
The activity of SOD was assessed using an ELISA assay [44]. Samples were brought to room temperature, resuspended, and analyzed using MyBioSource® ELISA kits (MBS2600843, Wuhan, China) per manufacturer instructions. Samples were analyzed at 450 nm, and the results were expressed as percent inhibition.
Statistical Analysis
Data were expressed as mean ± standard deviation (SD). Normality and homogeneity were assessed using the Shapiro-Wilk and Levene’s tests, respectively. One-way ANOVA with LSD post hoc was used for homogeneous data, while Tamhane’s was used for non-homogeneous data [44,45]. Statistical significance was set at p < 0.05. Analyses were performed using SPSS version 26.0.
Results and Discussion
Body weight changes during SPE treatment in CCl₄-induced rats
Body weight was monitored weekly (W0 - W3) (Table 1). All groups began with similar baseline weights (175 - 178.6 g). Over 3 weeks, every group gained weight, but to varying degrees: healthy controls (P1) gained the most (+24.2 g), while the CCl₄-treated groups (P2) showed the smallest increase (+14.1 g). NAC-treated groups (P3) and both SPE groups (P4: +20.0 g, P5: +24.1 g) showed intermediate gains. Notably, the highest SPE dose (P5) had a final weight trajectory comparable to healthy controls.
Table 1 Weekly body weight (g) of rats across experimental groups from week 0 to week 3.
Group (n = 6) |
Mean ± SD (g) |
W0 (g) |
W1 (g) |
W2 (g) |
W3 (g) |
P1 |
190.53 ± 8.95 |
178.6 |
186.6 |
194.1 |
202.8 |
P2 |
183.80 ± 5.25 |
175.5 |
183.5 |
186.6 |
189.6 |
P3 |
185.15 ± 7.36 |
175 |
182.5 |
188 |
195.1 |
P4 |
188.53 ± 7.33 |
178.3 |
186 |
191.5 |
198.3 |
P5 |
188.30 ± 8.92 |
176.5 |
184.3 |
191.8 |
200.6 |
Note: P1: Healthy control; P2: CCl4-treated control (C-); P3: NAC-treated control (C+); P4: CCl4-induced+SZE 400 mg/kg BW/day); P5: CCl4-induced+SZE 600 mg/kg BW/day). W: Weeks; W0 = Week 0 (baseline); W1 = Week 1; W2 = Week 2; W3 = Week 3.
Body weight is a key indicator of overall health and systemic effects in hepatotoxicity studies. CCl₄ exposure typically suppresses weight gain by impairing appetite, nutrient absorption, and metabolism through oxidative stress and liver injury. In this study, the CCl₄-treated groups (P2) showed the lowest weight gain, confirming previous findings [46]. NAC-treated groups (P3) and both SPE groups (P4, P5) experienced improved weight gain compared to P2. By week 3, P5 nearly matched the healthy control (P1), indicating that SPE may stabilize physiological function during CCl₄ exposure [47]. Benefits similar to those observed here have been reported for other polyphenol-rich extracts, such as pomegranate peel, grape seed [48], and Citrus flavonoids [49]. The outcomes shown in Figure 1 are attributed to improved biochemical health and reduced stress, although this study did not investigate metabolic mechanisms. Overall, S. zalacca peel extract reduced CCl₄-induced weight loss, supporting its hepatoprotective potential. Additional research on food intake, metabolism, and tissue health is recommended for a fuller understanding.
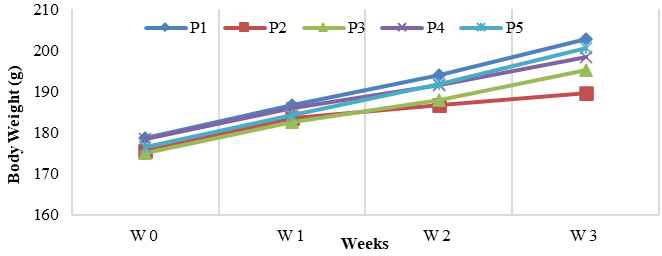
Figure 1 Body Weight Trends Demonstrating Physiological Recovery in SPE-Treated Rats Compared with CCl₄ Controls. P1: Healthy control; P2: CCl4-treated control (C-); P3: NAC-treated control (C+); P4: CCl4-induced + SZE 400 mg/kg BW/day); P5: CCl4-induced + SZE 600 mg/kg BW/day). W: Weeks; W0 = Week 0 (baseline); W1 = Week 1; W2 = Week 2; W3 = Week 3.
Effect of SPE on SGPT levels in CCl₄-induced wistar rats
The CCl₄-treated groups (P2) had the highest mean SGPT, indicating liver injury, while the healthy control (P1) had the lowest. SPE at 400 mg/kg (P4) and 600 mg/kg (P5) reduced SGPT significantly vs. the negative control (p < 0.001). Since SGPT data were normally distributed but not homogeneous, a one-way ANOVA (Table 2) and Tamhane post hoc test (Table 3) were performed. The CCl₄-treated groups (P2) had a significantly higher mean SGPT (43.37 ± 0.5902) than other groups (p < 0.001), while P3, P4, and P5 had lower values (29.37 ± 1.98; 29.05 ± 0.56; 23.46 ± 0.79). Significant differences were found among most groups: P2 > P1 (p < 0.001); P4 differed from P1, P2, and P5 (all p < 0.001) but not P3; and P5 differed from all but P3. P4 and P3 did not differ, and both SPE doses improved SGPT versus controls. The results are displayed in Figure 2.
SPE produced a clear reduction in SGPT levels in CCl₄-induced rats compared to untreated controls, indicating improved biochemical markers linked to hepatocellular stress [50]. Although no histological analysis was performed, the decrease in SGPT suggests a lessened biochemical disturbance after extract treatment. This effect is consistent with reports showing that polyphenol-rich plant materials can modulate oxidative processes in liver injury models [42,43]. The largest reduction was seen at 600 mg/kg, likely indicating a dose-related effect and justifying further study. Similar dose-dependent reductions in SGPT and SGOT have been observed with apple, potato, and pomegranate peel extracts in CCl₄-treated rats [51], where polyphenols enhance cellular stability and antioxidant capacity [52-54]. Improvements in other oxidative markers, such as SOD, CAT, and GSH, are also reported in similar models [55]. Elevated SGPT levels in the CCl₄ group match established patterns of acute hepatic stress. CCl₄ undergoes CYP2E1-dependent metabolism, producing reactive intermediates that trigger lipid peroxidation and membrane damage [44,56], processes associated with increased circulating transaminases in liver injury models, though these were not directly measured in this study. SGPT is widely acknowledged as a sensitive marker for hepatocellular leakage under oxidative stress, and its elevation in the untreated group is consistent with expectations. Both SPE doses lowered SGPT values relative to the CCl₄-only group.
Table 2 Results of descriptive analysis, normality test, homogeneity test, and 1-way ANOVA for the SGPT variable.
Group (n = 6) |
Mean ± SD (U/L) |
Saphiro-Wilk test (p) |
Levene’s test (p) |
1-way ANOVA test (p) |
P1 |
19.09 ± 0.50 |
0.46a |
< 0.001* |
< 0.001* |
P2 |
43.37 ± 0.59 |
0.41a |
|
|
P3 |
29.37 ± 1.98 |
0.26a |
|
|
P4 |
29.05 ± 0.56 |
0.42a |
|
|
P5 |
23.46 ± 0.79 |
0.50a |
|
|
Note:
P1: Healthy control; P2: CCl4-treated
control (C-); P3: NAC-treated
control (C+);
P4: CCl4-induced
+ SZE 400 mg/kg BW/day); P5: CCl4-induced
+ SZE 600 mg/kg BW/day). SGPT: Serum Glutamic Pyruvate Transaminase;
a =
p >
0.05; *
= p <
0.05.
Table 3 Results of Tamhane’s post hoc test for SGPT variables.
Group (n = 6) |
P2 |
P3 |
P4 |
P5 |
P1 |
< 0.001* |
< 0.001* |
< 0.001* |
< 0.001* |
P2 |
|
< 0.001* |
< 0.001* |
< 0.001* |
P3 |
< 0.001* |
|
1.000a |
0.003* |
P4 |
< 0.001* |
1.000a |
|
Note:
P1: healthy control; P2: CCl4-treated
control (C-); P3: NAC-treated
control (C+);
P4: CCl4-induced
+ SZE 400 mg/kg BW/day); P5: CCl4-induced
+ SZE 600 mg/kg BW/day). SGPT = serum glutamic pyruvate
transaminase;
a
= p
> 0.05; *
= p <
0.05.
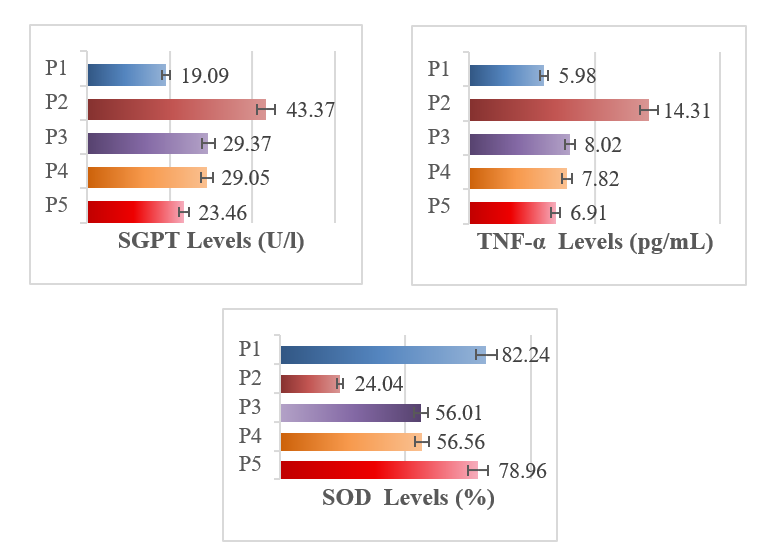
Figure 2 Graph of mean SGPT, TNF-α, and SOD levels among groups: Healthy control; P2: CCl4-treated control (C-); P3: NAC-treated control (C+); P4: CCl4-induced + SZE 400 mg/kg B W/day); P5: CCl4-induced + SZE 600 mg/kg BW/day). SGPT: Serum glutamic pyruvate transaminase; TNF-α = tumor necrosis factor-alpha; SOD = superoxide dismutase.
With the 600 mg/kg dose matching the effect of NAC [57,58]. While mechanisms behind these improvements remain to be clarified, the biochemical trends align with existing evidence that phytochemical-rich extracts affect oxidative responses. The included schematic summarizes in Figure 3, the most commonly cited mechanisms related to context [59,60].
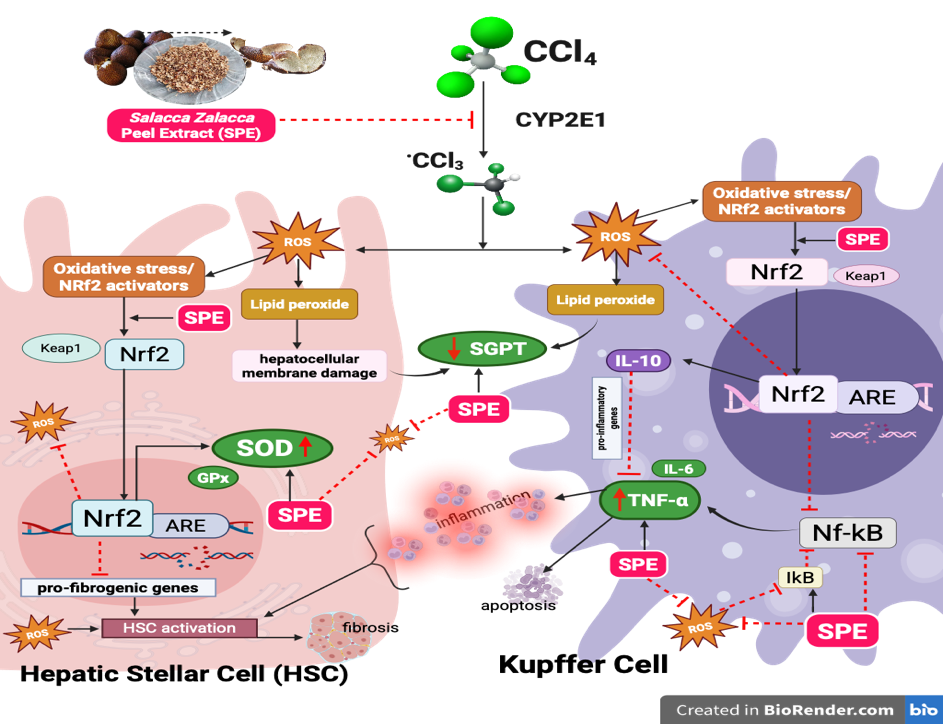
Figure 3 Hepatoprotective mechanism of SPE: NRF2-mediated regulation of kupffer cells and hepatic stellate cells in CCl₄-induced acute liver injury [59,60].
Effect of SPE on TNF-α levels in CCl₄-induced wistar rats
TNF-α levels were significantly higher in the negative control group exposed to CCl₄ (P2; 14.31 ± 0.23) than in the healthy group (P1; 5.98 ± 0.15). SPE treatment at 400 and 600 mg/kg reduced TNF-α to P4 (7.81 ± 0.36) and P5 (6.90 ± 0.14), respectively (p < 0.001). One-Way ANOVA (Table 4) and LSD Post Hoc tests (Table 5) showed significant group differences, with P5 showing the greatest reduction. P2 was higher than all other groups, and both P4 and P5 showed improvements over controls, especially at 600 mg/kg. Results are shown in Figure 2.
SPE treatment significantly reduced TNF-α levels in CCl₄-induced rats, reflecting improved biochemical markers of inflammatory stress. While this study did not assess the underlying inflammatory pathways or cellular mechanisms, the observed reduction mirrors findings from many studies where polyphenol-rich plant extracts modulate cytokine levels in oxidative liver injury models. Extracts with nootkatone [29], cyanidin [9], kaempferol [3], and various fruit peels (apple [51], blueberry, cranberry [47], spirulina [61]) have consistently demonstrated dose-dependent decreases in TNF-α, IL-6, and IL-1β in CCl₄-induced hepatotoxicity, often alongside improved liver histology and reduced inflammatory cell infiltration. The elevated TNF-α seen in the untreated CCl₄ group aligns with established patterns of hepatocellular stress and inflammation. Although this study did not explore mechanisms like NF-κB activity, IκB turnover, Kupffer cell activation, or fibrotic signaling, the reduction in TNF-α after SPE administration suggests a beneficial modulation of this inflammatory biomarker [56]. Literature often attributes such reductions to effects on NF-κB or Nrf2/HO-1 pathways by polyphenolics (Figure 3) [60,62]. In particular, both doses of SPE, especially 600 mg/kg, significantly decreased TNF-α relative to CCl₄ controls. While several bioactive compounds in SPE may contribute to these effects, specific mechanisms remain to be elucidated. Thus, the TNF-α reduction noted here should be seen as consistent with polyphenol-rich extracts in oxidative injury settings [63,64]. Further research using molecular and histological analysis is needed to clarify the responsible pathways.
Table 4 Results of descriptive analysis, normality test, homogeneity, and 1-way ANOVA for the TNF-α variable.
Group (n = 6) |
Mean ± SD (pg/mL) |
Saphiro-Wilk test (p) |
Levene’s test (p) |
1-way ANOVA test (p) |
P1 |
5.98 ± 0.15 |
0.83a |
0.018* |
< 0.001* |
P2 |
14.31 ± 0.23 |
0.78a |
|
|
P3 |
8.02 ± 0.22 |
0.96a |
|
|
P4 |
7.81 ± 0.36 |
0.36a |
|
|
P5 |
6.90 ± 0.14 |
0.97a |
|
|
Note:
P1: healthy control; P2: CCl4-treated
control (C-); P3: NAC-treated
control (C+);
P4: CCl4-induced
+ SZE 400 mg/kg BW/day); P5: CCl4-induced
+
SZE 600 mg/kg BW/day).
TNF-α = tumor necrosis factor-alpha; a
= p
>
0.05;
*
= p
<
0.05.
Table 5 Results of the LSD post hoc test for the TNF-α variable.
Group (n = 6) |
P2 |
P3 |
P4 |
P5 |
P1 |
< 0.001* |
< 0.001* |
< 0.001* |
0.027* |
P2 |
|
< 0.001* |
< 0.001* |
< 0.001* |
P3 |
< 0.001* |
|
0.156a |
< 0.001* |
P4 |
< 0.001* |
0.156a |
|
< 0.001* |
P5 |
< 0.001* |
< 0.001* |
< 0.001* |
|
Note:
P1: healthy control; P2: CCl4-treated
control (C-); P3: NAC-treated
control (C+);
P4: CCl4-induced
+
SZE 400 mg/kg BW/day);
P5: CCl4-induced
+ SZE 600 mg/kg BW/day). TNF-α = tumor necrosis factor-alpha; a
= p
>
0.05;
*
= p
<
0.05.
Effect of SPE on SOD levels in CCl₄-induced Wistar rats
The negative control group (P2; 24.04 ± 3.54) showed a significant reduction in SOD levels compared to the healthy control (P1; 82.24 ± 3.79), confirming oxidative stress. Treatment with SPE at 400 mg/kg BW (P4; 56.56 ± 3.06) and 600 mg/kg BW (P5; 78.96 ± 4.07) significantly increased SOD activity (p < 0.001). At 600 mg/kg BW, SOD levels returned close to normal and matched or exceeded levels in the NAC group (P3). ANOVA (Table 6) and LSD Post Hoc tests (Table 7) confirmed significant differences between groups (p < 0.001). P2 had the lowest SOD levels; P3, P4, and P5 were significantly higher. P4 was comparable to P3, differing from P1, P2, and P5, while P5 was distinct from all groups. Overall, 400 mg/kg BW SPE was similar to the positive control, and both doses significantly improved SOD activity compared to controls, as presented in Figure 2.
CCl₄ exposure significantly decreased SOD levels in the negative control group, underscoring the oxidative stress typical of this experimental model. SPE administration resulted in a dose-dependent increase in SOD activity, with the most pronounced effect observed in the 600 mg/kg group. Although this study did not assess antioxidant gene expression, redox signaling, or fibrosis-related pathways, the biochemical changes here are consistent with reports on various polyphenol-rich extracts [65], including grape seed [48], pomegranate [57], Vitis vinifera leaves [66], propolis [67], Shoumei tea [68], Sugarcane molasses [52], and Fimbristylis miliacea, which elevate SOD, CAT, and GSH levels in CCl₄ models, especially at higher doses and sometimes on par with NAC. These improvements are often accompanied by reduced MDA and healthier liver histology, though underlying molecular mechanisms are not always investigated [69]. Many studies increased SOD activity in relation to modulation of antioxidant systems such as the Nrf2-Keap1-ARE pathway [60] (Figure 3). Therefore, the current findings should be seen as evidence of biochemical recovery. SOD activity recovery in both SPE-treated groups, matching NAC at 400 mg/kg and surpassing it at 600 mg/kg, demonstrates a dose-dependent benefit for oxidative stress parameters. SPE improved SGPT, TNF-α, and SOD, highlighting its positive effects on biochemical markers of hepatocellular stress, inflammation, and antioxidant status.
Table 6 Results of descriptive analysis, normality test, homogeneity, and 1-way ANOVA for the SOD variable.
Group (n = 6) |
Mean ± SD (%) |
Saphiro-Wilk test (p) |
Levene’s test (p) |
1-way ANOVA test (p) |
P1 |
82.24 ± 3.79 |
0.80a |
0.986a |
< 0.001* |
P2 |
24.04 ± 3.54 |
0.96a |
|
|
P3 |
56.01 ± 4.07 |
0.79a |
|
|
P4 |
56.56 ± 3.06 |
0.96a |
|
|
P5 |
78.96 ± 4.07 |
0.79a |
|
|
Note: P1: healthy control; P2: CCl4-treated control (C-); P3: NAC-treated control (C+); P4: CCl4-induced + SZE 400 mg/kg BW/day); P5: CCl4-induced + SZE 600 mg/kg BW/day). SOD = superoxide dismutase; a = p > 0.05; * = p < 0.05.
Table 7 Results of the LSD post hoc test for the SOD variable.
Group (n = 6) |
P2 |
P3 |
P4 |
P5 |
P1 |
< 0.001* |
< 0.001* |
< 0.001* |
0.140a |
P2 |
|
< 0.001* |
< 0.001* |
< 0.001* |
P3 |
< 0.001* |
|
0.802a |
< 0.001* |
P4 |
< 0.001* |
0.802a |
|
< 0.001* |
Note: P1: healthy control; P2: CCl4-treated control (C-); P3: NAC-treated control (C+); P4: CCl4-induced + SZE 400 mg/kg BW/day); P5: CCl4-induced + SZE 600 mg/kg BW/day). SOD = superoxide dismutase; a = p > 0.05; * = p < 0.05.
SPE demonstrates antioxidant and anti-inflammatory effects by reducing SGPT and TNF-α levels and enhancing hepatic SOD levels. The present findings indicate biochemical improvement in SGPT, TNF-α, and SOD levels following SPE administration. However, without histopathological examination, structural recovery of hepatic tissues cannot be conclusively established. This represents a key limitation of the study and warrants future confirmation using H&E staining and fibrosis grading. Additionally, the assessment of SPE in this study was limited to a few liver injury biomarkers (SGPT, TNF-α, and SOD), which restricted a comprehensive evaluation of acute liver injury, antioxidant responses, and inflammatory processes. Furthermore, the absence of detailed histological evaluation and phytochemical profiling prevented the identification of the specific ethanolic content of SPE, highlighting the need for future studies to clarify its bioactive components and mechanisms.
In conclusion, ethanolic S. zalacca peel extract (SPE) demonstrated favorable modulation of biomarkers associated with liver injury in CCl₄-induced rats. Administration of 400 and 600 mg/kg BW was accompanied by reductions in SGPT and TNF-α levels and increases in SOD activity, with the higher dose showing the most pronounced improvement. Although this study did not include histological or molecular analyses, the overall biochemical pattern aligns with previous reports suggesting that polyphenol-rich plant materials may contribute to hepatic protection under oxidative stress conditions. SPE shows potential as a hepatoprotective agent in toxin-induced liver stress. However, further research involving tissue studies, mechanistic analysis, and phytochemical profiling is needed to confirm its effects and explain the mechanisms.
Acknowledgements
We would like to express our sincere appreciation to the research team of Integrated Biomedical Laboratories for kindly providing the Salacca zalacca peel extract and technical support throughout the study. We also extend our gratitude to the animal facility staff of the Center for Food and Nutrition Studies, Gadjah Mada University, Indonesia for their assistance in maintaining the experimental animals and ensuring the smooth execution of the in vivo procedures.
Declaration of Generative AI in Scientific Writing
The authors acknowledge the use of generative AI tools (e.g., QuillBot and ChatGPT by OpenAI) in the preparation of this manuscript, specifically for language editing and grammar correction. No content generation, data analysis, or interpretation was performed by AI. The authors take full responsibility for the content, scientific integrity, and conclusions presented in this work.
CRediT Author Statement
Danis Pertiwi: Conceptualization, Methodology, Supervision, Validation, Funding acquisition, Writing - original draft, and Writing - Reviewing; Matisatun Nisa: Data curation, Writing - Original draft preparation, Methodology, Investigation, and Validation; Laili Fitria Zulfa: Data curation, Formal analysis, Visualization, Software, Validation, Writing - original draft, Writing - Reviewing and Editing.
[1] J Vrba, B Papoušková, K Lněničková, P Kosina, V Křen and J Ulrichová. Metabolism of 2,3-dehydrosilybin A and 2,3-dehydrosilybin B: A study with human hepatocytes and recombinant UDP-glucuronosyltransferases and sulfotransferases. Antioxidants 2021; 10(6), 954.
[2] X Meng, GY Tang, PH Liu, CJ Zhao, Q Liu and HB Li. Antioxidant activity and hepatoprotective effect of 10 medicinal herbs on CCl4 -induced liver injury in mice. World Journal of Gastroenterology 2020; 26(37), 5629-5645.
[3] C Lee, S Yoon and JO Moon. Kaempferol suppresses carbon tetrachloride-induced liver damage in rats via the MAPKs/NF-κB and AMPK/Nrf2 signaling pathways. International Journal of Molecular Sciences 2023; 24(8), 6900.
[4] Y Ibn ElAchaouia, A Mahmoudi, D Dallali, R Marrakchi, G Marti, M Chamkha, S Sayadi and A Noureddine. UHPLC-HRMS-MS analysis, antioxidant and hepatoprotective effects of methanolic extracts from Ficus carica L., leaves and stem barks against acute CCl4 -induced liver damage. Natural Product Research 2025. https://doi.org/10.1080/14786419.2025.2498075
[5] S Munakarmi, L Chand, HB Shin, KY Jang and YJ Jeong. Indole-3-carbinol derivative DIM mitigates carbon tetrachloride-induced acute liver injury in mice by inhibiting inflammatory response, apoptosis and regulating oxidative stress. International Journal of Molecular Sciences 2020; 21(6), 2048.
[6] L Sun, Y Zhang, S Wen, Q Li, R Chen, X Lai, Z Zhang, Z Zhou, Y Xie, X Zheng, K Zhang, D Li and S Sun. Extract of Jasminum grandiflorum L. alleviates CCl4-induced liver injury by decreasing inflammation, oxidative stress and hepatic CYP2E1 expression in mice. Biomedicine & Pharmacotherapy 2022; 152, 113255.
[7] FT Johra, S Hossain, P Jain, AT Bristy, T Emran, R Ahmed, SM Sharker, AK Bepari and HM Reza. Amelioration of CCl4-induced oxidative stress and hepatotoxicity by Ganoderma lucidum in Long Evans rats. Scientific Reports 2023; 13(1), 9909.
[8] MM Erdogan, MA Erdogan, S Koc, A Yalcin, A Turk and EA Yetkin. Hepatoprotective effects of N-acetylcysteine on liver injury by irisin upregulation and oxidative stress reduction in diabetic rats. Egyptian Liver Journal 2023; 13(1), 33.
[9] B Wang, S Cui, B Mao, Q Zhang, F Tian, J Zhao, X Tang and W Chen. Cyanidin alleviated CCl4-induced acute liver injury by regulating the Nrf2 and NF-κB signaling pathways. Antioxidants 2022; 11(12), 2383.
[10] D Emad, AMA Bayoumi, SM Gebril, DME Ali and S Waz. Modulation of keap-1/Nrf2/HO-1 and NF-ĸb/caspase-3 signaling pathways by dihydromyricetin ameliorates sodium valproate-induced liver injury. Archives of Biochemistry and Biophysics. 2024; 758, 110084.
[11] P Hlásná Čepková, M Jágr, D Janovská, V Dvořáček, A Kotrbová Kozak and I Viehmannová. Comprehensive mass spectrometric analysis of snake fruit: Salak (Salacca zalacca). Journal of Food Quality 2021; 2021, 6621811.
[12] AAIM Padmiswari, NT Wulansari, KB Harditya and SD Megayanti. Antioxidant activity test of salak bali peel extract (Salacca zalacca var. amboinensis) against brain cells of male mice (Mus musculus L.) induced by alloxan. Jurnal Pijar Mipa 2025; 20(1), 35-39.
[13] Y Zhou, C Qian, Y Tang, M Song, T Zhang, G Dong, W Zheng, C Yang, C Zhong, A Wang, Y Zhao and Y Lu. Advance in the pharmacological effects of quercetin in modulating oxidative stress and inflammation related disorders. Phytotherapy Research 2023; 37(11), 4999-5016.
[14] H Khotimah, SN Alita, D Aninditha, A Weningtyas, WE Prima, U Kalsum, M Rahayu, D Handayani and SK Nandar. Ethanolic extract of Salacca zalacca peel reduce IL-1β and apoptosis in high glucose induced zebrafish embryo. GSC Biological and Pharmaceutical Sciences 2021; 16(3), 024-033.
[15] RM Hussein, DM Sawy, MA Kandeil and HS Farghaly. Chlorogenic acid, quercetin, coenzyme Q10 and silymarin modulate Keap1-Nrf2/heme oxygenase-1 signaling in thioacetamide-induced acute liver toxicity. Life Sciences 2021; 277, 119460.
[16] W Widowati, D Dani, V Vera and VA Yuninda. Antioxidant and antiaging potential of salak fruit extract (Salacca zalacca (Gaert.) Voss). Majalah Obat Tradisional 2023; 28(3), 230-236.
[17] PH Čepková, M Jágr, D Janovská, V Dvořáček, AK Kozak and I Viehmannová. Comprehensive mass spectrometric analysis of snake fruit: Salak (Salacca zalacca). Journal of Food Quality 2021; 2021, 662811.
[18] F Aghababaei and M Hadidi. Recent advances in potential health benefits of quercetin. Pharmaceuticals 2023; 16(7), 1020.
[19] OJ Sul and SW Ra. Quercetin prevents LPS-induced oxidative stress and inflammation by modulating NOX2/ROS/NF-kB in lung epithelial cells. Molecules 2021; 26(22), 6949.
[20] A Arhami, LT Djohan and H Rajagukguk. The effect of giving salak (Salacca Zalacca) bark extract on liver function and histopathological features of the liver of male wistar white rats obesity model. International Journal of Public Health Excellence 2024; 3(2), 607-616.
[21] DY Putri, Y Yueniwati, S Utami, MFR Syaban, N Halid, PP Putra, SP Putri and H Khotimah. Salacca zalacca skin as a potential adjuvant for metabolic syndrome therapy: Integrative in vitro and computational approaches. Trends in Sciences 2024; 22(11), 10678.
[22] S Utami, MFR Syaban, DY Putri, VAG Hose, H Khotimah and Y Yueniwati. Bioinformatics examination of quercetin from Salacca zalacca skin, fruit, and seed as a potent active compounds against hypercholesterolemia via PCSK9 inhibition. Trends in Sciences 2025; 22(4), 9237.
[23] E Zubaidah, Y Hasfiani and H Sujuti. Physicochemical properties and anti-cancer activity of Javanese turmeric kombucha (Curcuma xanthorrhiza) against T47D cell line. Trends in Sciences 2025; 22(9), 10417.
[24] E Girsang, C Ginting, I Lister, C Wijayanti, W Widowati and R Rizal. Antioxidant properties of Salacca zalacca (Gaertn.) voss peel ethanolic extract compared to chlorogenic acid. In: Proceedings of the 1st International Conference on Emerging Issues in Technology, Engineering and Science (ICE-TES 2021), Bandung, Indonesian, 2021, p. 87-94.
[25] E Coyago-Cruz, G Méndez, J Zúñiga-Miranda, N Jami, R Acurio-Vásconez and J Heredia-Moya. Dynamics of bioactive compounds and their relationship with antioxidant and antimicrobial activity in the pulp, peel, and seeds of ‘salak’ during ripening. Foods 2025; 14, 3476.
[26] JM Clarkson, JE Martin and DEF McKeegan. A review of methods used to kill laboratory rodents: Issues and opportunities. Laboratory Animals 2022; 56(5), 419-436.
[27] DL Hickman. Minimal exposure times for irreversible euthanasia with carbon dioxide in mice and rats. Journal of The American Association for Laboratory Animal Science 2022; 61(3), 283-286.
[28] MW Brunt, L Améndola and DM Weary. Attitudes of laboratory animal professionals and researchers towards carbon dioxide euthanasia for rodents and perceived barriers to change. Laboratory Animals 2021; 55(6), 531-539.
[29] C Dai, X Zhang, J Lin and J Shen. Nootkatone supplementation ameliorates carbon tetrachloride-induced acute liver injury via the inhibition of oxidative stress, NF-κB pathways, and the activation of Nrf2/HO-1 pathway. Antioxidants 2023; 12(1), 194.
[30] S Wang, W Li, W Liu, L Yu, F Peng, J Qin, L Pu, Y Tang, X Xie and C Peng. Total flavonoids extracted from Penthorum chinense Pursh mitigates CCl4-induced hepatic fibrosis in rats via inactivation of TLR4-MyD88-mediated NF-κB pathways and regulation of liver metabolism. Frontiers in Pharmacology 2023; 14, 1253013.
[31] B Pandey, S Thapa, A Kaundinnyayana and S Panta. Hepatoprotective effects of Juglans regia on carbon tetrachloride‐induced hepatotoxicity: In silico/in vivo approach. Food Science & Nutrition 2024; 12(9), 6482-6497.
[32] R Setyawaty, RB Aptuning and Dewanto. Preliminary studies on the content of phytochemical compounds on skin of salak fruit (Salacca zalacca). Pharmaceutical Journal of Indonesia 2020; 6(1), 1-6.
[33] Mohammed, SM Saleh, M Siddiqui, S Mat So’Ad, S Murugesu, A Khatib and M Rahman. Antioxidant and α-glucosidase inhibitory activities and gas chromatography-mass spectrometry profile of salak (Salacca zalacca) fruit peel extracts. Pharmacognosy Research 2018; 10(4), 385.
[34] Sukamto, KM Amalia and Sudiyono. Effect of drying temperature and wilting period on the physical and sensory properties of indonesian pondoh snake fruit (Salacca zalacca) powder. International Journal of Current Science Research and Review 2025; 08(5), 2439-2444.
[35] DPB Suri, A Ningrum, MP Kurniawan, ZA Athaillah and Manikharda. Valorization of snake fruit (Salacca edulis R) seeds for coffee subtitutes: Impact of drying and roasting on bioactive compounds and sensory profile. Biocatalysis and Agricultural Biotechnology 2025; 66, 103613.
[36] RK Saputri, A Al-Bari and DI Pramesti. Phytochemical screening and evaluation of transparent soap with addition of wedi snakefruit (Salacca zalacca) rind extract. In: Proceedings of the 4th Green Development International Conference (GDIC 2022), Jambi, Indonesia. 2023, p. 78-86.
[37] PD Andina, D Indarto and LO Wardhani. Phytochemical and nutrient analysis of ethanol extract of Salak Pondoh (Sallaca Edulis Reinw) peel for the development of anti-obesity. Media Penelitian dan Pengembangan Kesehatan 2025; 35(2), 467-476.
[38] D Pertiwi, AP Aulia, JW Wibowo, M Nisa, D Guscella and N Anggita. The effect of ethanol extract from Salacca zalacca (Gaertn.) voss peel on antioxidant and uric acid levels in hyperuricemia rats. Tropical Journal of Natural Product Research 2025; 9(4), 1706-1710.
[39] National Agency of Drug and Food Control of the Republic of Indonesia. Regulation of the national agency of drug and food control number 20 of 2023 on guidelines for preclinical pharmacodynamic testing of traditional medicines. National Agency of Drug and Food Control of the Republic of Indonesia, Central Jakarta, Indonesia, 2023.
[40] EJ Jo, E Bae, JH Yoon, JY Kim and JS Han. Comparison of murine retroorbital plexus and facial vein blood collection to mitigate animal ethics issues. Laboratory Animal Research 2021; 37(1), 12.
[41] US Aguwa, CE Eze, BN Obinwa, SN Okeke, SF Onwuelingo, DI Okonkwo, DK Ogbuokiri, AE Agulanna, IJ Obiesie and AJ Umezulike. Comparing the effect of methods of rat euthanasia on the brain of wistar rats: Cervical dislocation, chloroform inhalation, diethyl ether inhalation and formalin inhalation. Journal of Advances in Medicine and Medical Research 2020; 32(17), 8-16.
[42] G Ouais and DM Hassan. A comparative study on the hepatoprotective effect of selenium-nanoparticles and dates flesh extract on carbon tetrachloride induced liver damage in albino rats. Anatomy & Cell Biology 2023; 56(4), 538-551.
[43] X Yu, L Huang, C You and L Huang. Hepatoprotective Effects of Polysaccharide from Anoectochilus roxburghii (Wall.) Lindl. on Rat Liver Injury Induced by CCl4. Drug Design, Development and Therapy 2021; 15, 2885-2897.
[44] N Iqbal, HM Zubair, MH Almutairi, M Abbas, MF Akhtar, L Aleya, M Kamel, A Saleem, Q Jabeen, S Noreen, MMFA Baig and MM Abdel-Daim. Hepatoprotective effect of Cordia rothii extract against CCl4-induced oxidative stress via Nrf2–NFκB pathways. Biomedicine & Pharmacotherapy 2022; 156, 113840.
[45] J Juarros-Basterretxea, G Aonso-Diego, Á Postigo, P Montes-Álvarez, Á Menéndez-Aller and E García-Cueto. Post-hoc tests in 1-way ANOVA: The case for normal distribution. Methodology 2024; 20(2), 84-99.
[46] I Touiss, S Ouahhoud, M Harnafi, S Khatib, O Bekkouch, S Amrani and H Harnafi. Toxicological evaluation and hepatoprotective efficacy of rosmarinic acid-rich extract from Ocimum basilicum L. Evidence-Based Complementary and Alternative Medicine 2021; 2021, 6676998.
[47] S Sergazy, Z Shulgau, Y Kamyshanskiy, Z Zhumadilov, E Krivyh, A Gulyayev and M Aljofan. Blueberry and cranberry extracts mitigate CCL4-induced liver damage, suppressing liver fibrosis, inflammation and oxidative stress. Heliyon 2023; 9(4), e15370.
[48] NA Madbouly, DM Ali and AA Farid. Nanoparticles from grape seed extract inhibit inflammatory cytokines and ameliorate CCl4-induced hepatotoxicity. BMC Complementary Medicine and Therapies 2025; 25(1), 276.
[49] N Ammar, H Hassan, H Abdallah, S Afifi, A Elgamal, A Farrag, A El-Gendy, M Farag and A Elshamy. Protective effects of naringenin from Citrus sinensis (var. Valencia) peels against CCl4-induced hepatic and renal injuries in rats assessed by metabolomics, histological and biochemical analyses. Nutrients 2022; 14(4), 841.
[50] M Kim, SC Jee and JS Sung. Hepatoprotective effects of flavonoids against benzo[a]pyrene-induced oxidative liver damage along its metabolic pathways. Antioxidants 2024; 13(2), 180.
[51] H Yousefi-Manesh, AR Dehpour, S Ansari-Nasab, S Hemmati, MA Sadeghi, RH Shahraki, S Shirooie, SM Nabavi, JG Nkuimi Wandjou, S Sut, G Caprioli, S Dall’Acqua and F Maggi. Hepatoprotective effects of standardized extracts from an ancient italian apple variety (Mela Rosa Dei Monti Sibillini) against carbon tetrachloride (CCl4)-induced hepatotoxicity in rats. Molecules 2020; 25(8), 1816.
[52] M Wang, L Zhao, Y Wang, C Zhang and H Li. Sugarcane molasses polyphenol extract attenuates alcohol-induced chronic liver damage via antioxidant, anti-inflammatory, and CYP2E1/Keap1/NF-κB pathway modulation. Nutrients 2025; 17(9), 1589.
[53] D Wang, T Wang, Z Li, Y Guo and D Granato. Green tea polyphenols upregulate the Nrf2 signaling pathway and suppress oxidative stress and inflammation markers in D-galactose-induced liver aging in mice. Frontiers in Nutrition 2022; 9, 836112.
[54] IF Machado, RG Miranda, DJ Dorta, AP Rolo and CM Palmeira. Targeting oxidative stress with polyphenols to fight liver diseases. Antioxidants 2023; 12(6), 1212.
[55] GM Kondepudi, BG Rao and P Balakrishnaiah. Evaluation of hepatoprotective effects of selected fruit peel extracts and their poly herbal mixture on CCl4 induced hepatotoxicity: An in vivo study. Indian Drugs 2020; 57(9), 67-74.
[56] R Li, W Yang, Y Yin, X Ma, P Zhang and K Tao. 4-OI attenuates carbon tetrachloride-induced hepatic injury via regulating oxidative stress and the inflammatory response. Frontiers in Pharmacology 2021; 12, 651444.
[57] H Ali, A Jahan, S Samrana, A Ali, S Ali, N Kabir, A Ali, R Ullah, RA Mothana, BN Murtaza and M Kalim. Hepatoprotective potential of pomegranate in curbing the incidence of acute liver injury by alleviating oxidative stress and inflammatory response. Frontiers in Pharmacology 2021; 12, 694607.
[58] TI Tithi, MdR Tahsin, J Anjum, TS Zaman, F Aktar, NB Bahar, S Tasnim, A Sultana, I Jahan, SS Afrin, T Akter, P Sen, FJ Koly, MdS Reza, JA Chowdhury, S Kabir, AA Chowdhury and S Amran. An in vivo and in silico evaluation of the hepatoprotective potential of Gynura procumbens: A promising agent for combating hepatotoxicity. PLoS One 2023; 18(9), e0291125.
[59] A Allameh, R Niayesh-Mehr, A Aliarab, G Sebastiani and K Pantopoulos. Oxidative stress in liver pathophysiology and disease. Antioxidants 2023; 12(9), 1653.
[60] JS Park, N Rustamov and YS Roh. The roles of NFR2-regulated oxidative stress and mitochondrial quality control in chronic liver diseases. Antioxidants 2023; 12(11), 1928.
[61] NA Mohamed, MAM Hashem, AM Alzahrani, AM Abdel-Moneim and HM Abdou. Hepatoprotective effect of Spirulina platensis against carbon tetrachloride-induced liver injury in male rats. Journal of Pharmacy and Pharmacology 2021; 73(11), 1562-1570.
[62] FC Liu, HC Lee, CC Liao, AH Chou and HP Yu. Role of NADPH oxidase-derived ROS-mediated IL-6/STAT3 and MAPK/NF-κB signaling pathways in protective effect of corilagin against acetaminophen-induced liver injury in mice. Biology 2023; 12, 334.
[63] Q Zou, N Wang, Z Gao, H Xu, G Yang, T Zhang, Z Guo, Z Zhang, L Jia and X Chen. Antioxidant and hepatoprotective effects against acute CCl 4 ‐induced liver damage in mice from red‐fleshed apple flesh flavonoid extract. Journal of Food Science 2020; 85(10), 3618-3627.
[64] K Yin, X Li, X Luo, Y Sha, P Gong, J Gu and R Tan. Hepatoprotective effect and potential mechanism of aqueous extract from Phyllanthus emblica on carbon-tetrachloride-induced liver fibrosis in rats. Evidence-Based Complementary and Alternative Medicine 2021; 2021, 5345821.
[65] Y Lu, Y Hong, T Sun, F Li, Y Li, B Liu, W Wang and C Gao. Lycopus lucidus Turcz. extract ameliorates CCl4-induced liver damage by MAPK/Nrf2 signaling pathways and gut microbiota in mice. Journal of Functional Foods 2025; 128, 106834.
[66] N Saadaoui, A Mathlouthi, A Zaiter, S El-Bok, M Mokni, M Harbi, N Ghanem-Boughanmi, A Dicko and M Ben-Attia. Phytochemical profiling, antioxidant potential and protective effect of leaves extract of tunisian Vitis vinifera autochthonous accessions against acute CCl4-injured hepatotoxicity in mice. Heliyon 2023; 9(5), e16377.
[67] E Wani, T Nasihun and A Hussaana. The influence of continuous multi stage countercurrent extraction process (CMCE) propolis extract administration on superoxide dismutase activity and hepatocytes fibrosis degree (an experimental study on male wistar rats induced with carbon tetrachloride (CCl4)). Sains Medika 2020; 11(1), 42.
[68] R Yi, Y Wei, F Tan, J Mu, X Long, Y Pan, W Liu and X Zhao. Antioxidant capacity-related preventive effects of shoumei (slightly fermented Camellia sinensis) polyphenols against hepatic injury. Oxidative Medicine and Cellular Longevity 2020; 2020, 9329356.
[69] IG Ko, JJ Jin, L Hwang, SH Kim, CJ Kim, JH Han, S Lee, HI Kim, HP Shin and JW Jeon. Polydeoxyribonucleotide exerts protective effect against CCl4-induced acute liver injury through inactivation of NF-κB/MAPK signaling pathway in mice. International Journal of Molecular Sciences 2020; 21(21), 7894.