Trends
Sci.
2026;
23(8):
12564
Morphological and Morphometric Characteristics of Crustaceans (Decapoda) in the East Coast of Aceh Province, Indonesia
Irfannur
Irfannur1,*,
![]() ,
Syifa
Saputra2,
,
Syifa
Saputra2,
![]() ,
Agung
Setia Batubara3,
,
Agung
Setia Batubara3,
![]() ,
Siti
Annisa Ramadhani1,
Muliari
Muliari4,
,
Siti
Annisa Ramadhani1,
Muliari
Muliari4,
![]() ,
Muhammad
Radhi1,
,
Muhammad
Radhi1,
![]() ,
Yusrizal
Akmal1,
,
Yusrizal
Akmal1,
![]() and
Ilham
Zulfahmi5,
and
Ilham
Zulfahmi5,
![]()
1Department
of Aquaculture, Faculty of Agriculture, Universitas Almuslim,
Bireuen, Indonesia
2Department
of
Forestry,
Faculty of Agriculture, Universitas Almuslim, Bireuen, Indonesia
3Department of Biology, Faculty of Mathematics and Natural Sciences, Universitas Negeri Medan,
North Sumatra, Indonesia
4Department of Marine Science, Faculty of Agriculture, Universitas Malikussaleh, Aceh Utara, Indonesia
5Department of Fisheries Resources Utilization, Faculty of Marine and Fisheries, Universitas Syiah Kuala,
Banda Aceh, Indonesia
(*Corresponding author’s e-mail: [email protected])
Received: 7 November 2025, Revised: 21 December 2025, Accepted: 28 December 2025, Published: 5 March 2026
Abstract
The east coast of Aceh Province, including Pidie, North Aceh, and Aceh Tamiang Regencies, is a producer of Decapod (Crustacea), a marine capture fishery commodity. Decapod fishing activities are suspected to be excessive on the east coast of Aceh Province. Therefore, this study aims to map resources, identify morphology, and morphometrics of Decapods in these waters, as an initial step toward supporting sustainable management. This study was conducted from June to September 2025. Sampling was carried out in the waters of North Aceh Regency (Site 1), Pidie Regency (Site 2), and Aceh Tamiang Regency (Site 3). The collected samples were then taken to the Mathematics and Natural Sciences Laboratory of Almuslim University for further analysis. The analysis carried out included morphological and morphometric identification. There are 8 Decapods species caught in the waters of the east coast of Aceh Province, including Charybdis feriata, Portunus sanguinolentus, Portunus pelagicus, Penaeus monodon, Metapenaeus monoceros, Penaeus merguiensis, Panulirus homarus, and Panulirus ornatus. In the North Aceh Regency, the species caught were Portunus sanguinolentus, Penaeus monodon, Metapenaeus monoceros, and Panulirus ornatus. In the Pidie Regency, the species caught were Portunus sanguinolentus, Charybdis feriata, Penaeus merguiensis, and Panulirus homarus. Furthermore, in the Aceh Tamiang Regency, the species caught were Portunus pelagicus, Penaeus monodon, Metapenaeus monoceros, and Penaeus merguiensis. This study addresses this gap by providing a comprehensive morphometric baseline and taxonomic identification of 8 commercially vital species. The results reveal notable morphological variability in Panulirus homarus and high habitat specificity for Charybdis feriata, highlighting the influence of local ecological conditions on population structure. These findings offer a novel contribution to regional fisheries management, serving as a critical reference for establishing sustainable exploitation limits in the Malacca Strait ecosystem.
Keywords: Crustacea, Decapods, Lobsters, Morphology crabs, Shrimps
Introduction
Marine fishing activities play a crucial role in maintaining global food security and economic stability [1]. The crustacean fisheries sector, in particular,
accounts for nearly 30% of the total fish and shellfish catches worldwide [2] and possesses substantial economic value [3]. Crustacea represent an important source of aquatic dietary protein [4], making a significant contribution to the global aquatic animal protein supply [5]. This group is distributed across virtually all aquatic habitats, including the diverse order Decapoda, which encompasses economically vital species such as crabs, shrimp, and lobsters [6]. Decapoda is one of the most diverse crustacean orders [7], currently comprising approximately 14,756 species worldwide [8]. With over 4,400 species of decapod shrimp alone exhibiting significant economic value [9], the global wild capture fisheries of decapod crustaceans are reported to be rapidly growing [10]. Due to this high market potential, the aquaculture of decapod crustaceans has also become a main economic driver in the fisheries sector [11].
The crustacean fisheries sector holds a pivotal position within the Indonesian fisheries industry [12]. Specifically, the waters of Aceh Province, Indonesia, are notable for their high production of crustaceans, sourced from both wild capture and aquaculture activities, with a particular focus on the order Decapoda. The eastern coast of Aceh Province, encompassing the regencies of Pidie, North Aceh, and Aceh Tamiang, functions as a major production hub for commercially valuable decapod crustaceans [13]. The substantial output in this region is ultimately achieved through the combined contribution of aquaculture systems and capture fisheries [14]. Despite the economic prominence of these regencies, local scientific documentation regarding the specific species composition and morphological variations of Decapods in these waters remains fragmented. While broader studies have addressed Indonesian marine biodiversity, there is a distinct research gap concerning the updated taxonomic baseline and morphometric characteristics of commercially exploited Decapods specifically within Aceh’s coastal ecosystems. This lack of localized data hinders the development of province-specific conservation and harvesting quotas.
However, the exploitation and management of these resources necessitate prudent regulation. Unregulated fishing practices risk damaging stocks and habitats, thereby compromising both ecosystem integrity and economic sustainability [15-17]. Consequently, research focusing on the biology, distribution, habitat, and reproductive biology of decapod crustaceans is essential for implementing effective and sustainable fisheries management [18,19]. Therefore, this study aims to: (1) provide a comprehensive taxonomic identification of Decapod Crustacean species found across 3 major production hubs in Aceh; (2) analyze the morphological and morphometric characteristics of these species to establish a phenotypic baseline; and (3) map the distribution of these resources to support targeted, sustainable resource management strategies in the region.
Materials and methods
This research was conducted over a 4-month period, spanning from June to September 2025. Sampling was conducted at 3 designated locations within Aceh Province, Indonesia: North Aceh Regency (Location 1), Pidie Regency (Location 2), and Aceh Tamiang Regency (Location 3). The geographical context of the study area is illustrated in Figure 1. All collected specimens were subsequently transported to the Mathematics and Natural Sciences Laboratory at Universitas Almuslim, Aceh Province, for detailed analysis.
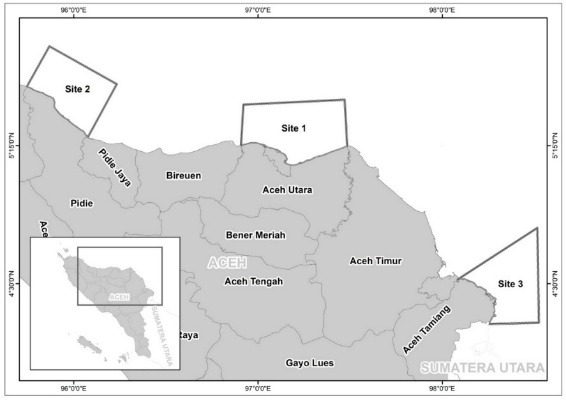
Figure 1 Map of research locations including North Aceh Regency (Site 1), Pidie Regency (Site 2), and Aceh Tamiang Regency (Site 3), Aceh Province, Indonesia.
Decapod Crustacean samples were systematically collected from direct landings at various Fish Landing Ports (PPIs) and Fish Auction Places (TPIs) across 3 locations: Pidie, North Aceh, and Aceh Tamiang Regencies. A total of 10 adult individuals per species were secured for analysis, representing a pooled sample from these 3 distinct study sites. The sampling criteria focused on adult specimens with weights up to 500 g and lengths ranging from 18 to 25 cm.
Morphometric measurements were performed on these 10 representative samples for each Decapod species. The measurements, which included the width, height, and length of specified morphological features, were executed using a digital caliper with an accuracy of 0.1 mm. The derived morphometric data (Tables 1 - 3) were subsequently processed using a modified analytical approach based on established methods [20-22]. Data transformation was performed according to the formula established by Batubara et al. [23]:
Mtrans = (M×100)/CL
where Mtrans is the transformed morphometric character data, M is the measured morphometric character data, and CL is the carapace length. The detailed parameters measured are outlined in Figure 2. The final morphometric data underwent descriptive statistical analysis, which included the calculation of mean values, intervals, and standard deviations.
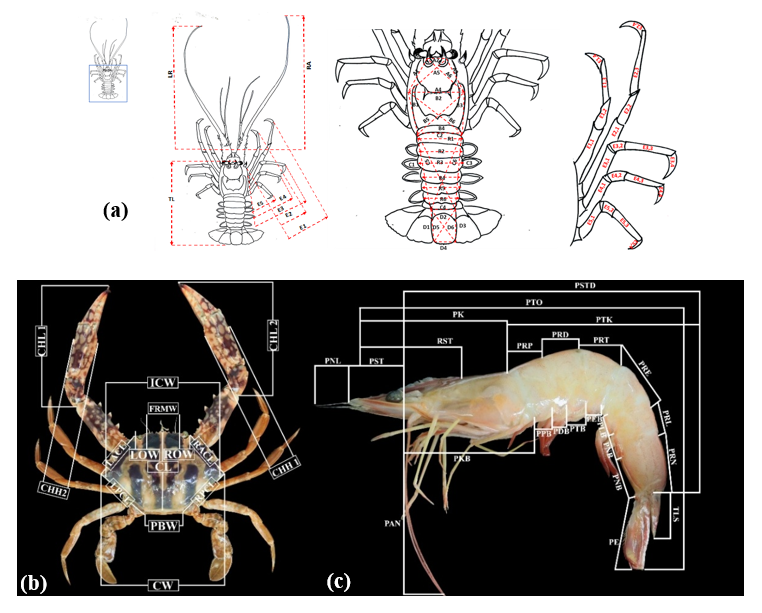
Figure 2 The morphometric measurement of (a) Lobster, (b) swimming crab, and (c) Shrimp. Measurement designations are given in Tables 1 - 3 [22].
Table 1 Lobster morphometric measurements [22].
Body parts |
Code |
Information |
Anterior Cephalothorax |
A1 |
The distance between the widest left point of the carapace to the base of the left eye stalk |
A2 |
The distance between the right and left of the eye stalks |
|
A3 |
The distance between the widest right point of the carapace to the base of the left eye stalk |
|
A4 |
The distance between the widest carapace points |
|
A5 |
Diagonally point left carapace to right eye |
|
A6 |
Diagonally point right carapace to left eye |
|
RA |
Right antenna |
|
LA |
Left antenna |
|
Posterior Cephalothorax |
B1 |
The distance from the posterior point of the carapace to the point on the widest part of the carapace on the left region |
B2 |
The distance between the widest carapace points |
|
B3 |
The distance between the posterior point of the carapace to the point of the widest part of the carapace on the right region |
|
B4 |
The distance between the left and right points of the widest carapace posteriorly |
|
B5 |
Diagonal from left posterior carapace point to right widest carapace point |
|
B6 |
Diagonal from the right posterior carapace point to the left widest carapace point |
|
Abdomen |
C1 |
The distance between the left posterior abdominal point to the left anterior abdominal point |
C2 |
The distance between the 2 left and right points of the posterior carapace |
|
C3 |
The distance between the posterior point of the right abdomen to the anterior point of the right abdomen |
|
C4 |
The distance between the right posterior abdominal point to the left side |
|
C5 |
Diagonal from the posterior point of the left abdomen to the anterior point of the right abdomen |
|
C6 |
Diagonal from the posterior point of the right abdomen to the anterior point of the left abdomen |
|
Walking Leg (Pereiopod) |
E1 |
The length between the base of the foot to the tip of the first foot |
E2 |
The length between the base of the foot to the tip of the second foot |
|
E3 |
The length between the base of the foot to the tip of the third foot |
|
E4 |
The length between the base of the foot to the tip of the fourth foot |
|
E5 |
The length between the base of the foot to the tip of the fifth foot |
|
E1,1 |
The distance from the ischium point to the merus point of the first foot |
|
E1,2 |
The distance between the merus point to the carpus point of the first foot |
|
E1,3 |
The distance from the carpus point to the propodus point of the first foot |
|
E1,4 |
Distance between propodus point to unguiculate point of the first foot |
|
E2,1 |
The distance from the ischium point to the merus point of the second foot |
|
Abdominal Somites |
R1 |
Diagonal from the right side point to the left side point of the first abdominal somites |
R2 |
Diagonal from the right side point to the left side point of the second abdominal somites |
|
R3 |
Diagonal from the right side point to the left side point of the third abdominal somites |
|
R4 |
Diagonal from the right side point to the left side point of the fourth abdominal somites |
|
R5 |
Diagonal from the right side point to the left side point of the fifth abdominal somites |
|
R6 |
Diagonal from the right side point to the left side point of the sixth abdominal somites |
|
Caudal (Telson) |
D1 |
The distance between the posterior point of the left telson to the anterior point of the left telson |
D2 |
The distance between the left and right points of the anterior telson |
|
D3 |
The distance between the posterior point of the right telson to the anterior point of the right telson |
|
D4 |
The distance between the left and right points of the posterior telson |
|
D5 |
Diagonal from the posterior point of the telson on the left to the anterior point of the right telson |
|
D6 |
Diagonal from the posterior point of the right telson to the anterior point of the left telson |
Table 2 Swimming crab morphometric measurements.
Character |
Code |
Information |
Carapace width |
CW |
Distance from the tip of the right marginal spine to the tip of the left marginal spine. |
Internal carapace width |
ICW |
Internal distance or internal limit. |
Carapace length |
CL |
Distance between the width or length of the body shell. |
Left anterolateral carapace length |
LACL |
The width of the left side of the carapace in the front and lateral areas of the midline of the body. |
Right anterolteral carapace length |
RACL |
The width of the right side of the carapace in the front and lateral areas of the midline of the body. |
Left posterolateral carapace length |
LPCL |
Distance from the center of the back of the carapace to the end of the posterolateral part of the carapace on the left side. |
Right posterolateral carapace length |
RPCL |
Distance from the center of the back of the carapace to the end of the posterolateral part of the carapace on the right side. |
Frontal margin width |
FRMW |
Distance between the front of the carapace that has spines. |
Left orbit width |
LOW |
Distance between the left eye and other body parts parallel to the carapace. |
Right orbit width |
ROW |
Distance between the right eye and other body parts parallel to the carapace. |
Posterior margin |
PBW |
Distance at the back of the posterior carapace. |
Left claw length |
CHH1 |
Left claw length. |
Right claw length |
CHH2 |
Right claw length. |
Left claw width |
CHL1 |
Left claw width. |
Right claw width |
CHL2 |
Right claw width. |
Table 3 Shrimp morphometric measurements.
Character |
Code |
Information |
Rostrum length |
RST |
Distance from the base of the eye to the tip of the rostrum. |
Carapace length |
CL |
Distance from the orbital margin to the posterior edge of the carapace. |
First abdominal segment length |
FSL |
Distance from the posterior margin of the carapace to the posterior end of the first abdominal segment. |
Second abdominal segment length |
SSL |
Distance from the posterior end of the first abdominal segment to the posterior end of the second abdominal segment. |
Third abdominal segment length |
TSL |
Distance from the posterior end of the second abdominal segment to the posterior end of the third abdominal segment. |
Fourth abdominal segment length |
FoSL |
Distance from the posterior end of the third abdominal segment to the posterior end of the fourth abdominal segment. |
Fifth abdominal segment length |
FiSL |
Distance from the posterior end of the fourth abdominal segment to the posterior end of the fifth abdominal segment. |
Sixth abdominal segment length |
SiSL |
Distance from the posterior end of the fifth abdominal segment to the posterior end of the sixth abdominal segment. |
Telson length |
TLS |
Distance from the base to the tip of the telson. |
Scaphocerite length |
SCL |
Distance from the base to the tip of the scaphocerite (antennal blade). |
Antennule length |
ANL |
Distance from the base to the tip of the antennule. |
Antenna length |
ANT |
Distance from the base to the tip of the antenna. |
Total length |
TL |
Distance from the tip of the rostrum to the tip of the telson with the abdomen straightened. |
Standard length |
SL |
Distance from the base of the rostrum to the base of the sixth abdominal segment. |
Lower carapace length |
LCL |
Distance from the tip of the antenna to the posterior edge of the lower carapace. |
First abdominal segment ventral length |
V1SL |
Distance from the posterior end of the lower carapace to the posterior end of the first ventral segment. |
Second abdominal segment ventral length |
V2SL |
Distance from the posterior end of the first ventral segment to the posterior end of the second ventral segment. |
Third abdominal segment ventral length |
V3SL |
Distance from the posterior end of the second ventral segment to the posterior end of the third ventral segment. |
Fourth abdominal segment ventral length |
V4SL |
Distance from the posterior end of the third ventral segment to the posterior end of the fourth ventral segment. |
Fifth abdominal segment ventral length |
V5SL |
Distance from the posterior end of the fourth ventral segment to the posterior end of the fifth ventral segment. |
Sixth abdominal segment ventral length |
V6SL |
Distance from the posterior end of the fifth ventral segment to the posterior end of the sixth ventral segment. |
Uropod length |
UL |
Distance from the posterior end of the sixth abdominal segment to the tip of the uropod. |
Body length without carapace |
BLC |
Distance from the base of the first abdominal segment to the tip of the tail. |
Results and discussion
Species diversity and distribution
Based on the identification of decapod crustaceans harvested along the eastern coast of Aceh Province, 8 commercially valuable species were identified (Figure 3). These species belong to the families Portunidae (crabs), Penaeidae (shrimps), and Palinuridae (spiny lobsters), and include: Charybdis feriata, Portunus sanguinolentus, Portunus pelagicus, Penaeus monodon, Metapenaeus monoceros, Penaeus merguiensis, Panulirus homarus, and Panulirus ornatus (Table 4).

Figure 3 Decapod species collected from the waters of the east coast of Aceh Province including (1) Charybdis feriata, (2) Portunus sanguinolentus, (3) Portunus pelagicus, (4) Penaeus monodon, (5) Metapenaeus monoceros, (6) Penaeus merguiensis, (7), Panulirus Homarus, and (8) Panulirus ornatus.
Table 4 The distribution of decapod species varied across the sampling locations.
Regency (Location) |
Identified decapod species |
North Aceh (Location 1) |
Portunus sanguinolentus, Penaeus monodon, Metapenaeus monoceros, and Panulirus ornatus. |
Pidie (Location 2) |
Portunus sanguinolentus, Charybdis feriata, Penaeus merguiensis, and Panulirus homarus. |
Aceh Tamiang (Location 3) |
Portunus pelagicus, Penaeus monodon, Metapenaeus monoceros, and Penaeus merguiensis. |
Morphological characteristics
The detailed external morphology of each species was documented for all identified species, focusing on key features of the carapace, appendages, and body coloration (Figures 4 - 11).
Crabs (portunidae)
The identified crab species exhibit distinct morphological features. The Coral Crab (C. feriata) is characterized by a broad, round carapace featuring 6 prominent spines on each anterolateral margin and a conspicuous red and white striped pattern. The Star Crab (P. sanguinolentus) possesses a relatively slender, grayish-green carapace marked by 3 distinct red spots on its posterior aspect. In contrast, the Common Swimming Crab (P. pelagicus) is identified by its wide, flattened carapace, which bears 9 spines lateral to the eyes, with the terminal spine being notably the largest and longest. For all species, the general body structure is divided into distinct dorsal and ventral regions, comprising structures such as the dactylus, carapace, pleon, and associated musculature (tendons of adductor/abductor muscles) (Figures 4 - 6).
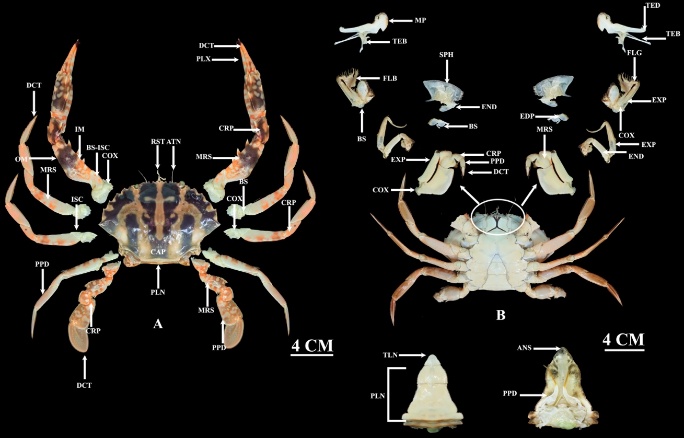
Figure 4 Anatomy of Charybdis feriata, (A) Dorsal, where DCT: Dactylus,PLX: Pollex, CRP: Carpus, MRS: Merus, BS: Basis, COX: COXA, PPD: Propodus, PLN: Pleon, CAP: Carapace, ISC: Ischium, ATN: Antenna, RST: Rostrum, BS-ISC: Basis-Ischium IM: Inner margin, OM: Outer margin, and (B) Ventral, where TED: Tendon of external adductor muscle, TEB: Tendon of external abductor muscle, FLG: Flagellum, EXP: Exopod, EDP: Endopod, SPH: Scaphognathite, FLB: Flabellum, MP: Mandibullar palp, TLN: Telson, ANS: Anus.

Figure 5 Anatomy of Portunus sanguinolentus, (A) Dorsal, where DCT: Dactylus,PLX: Pollex, CRP: Carpus, MRS: Merus, BS: Basis, COX: COXA, PPD: Propodus, PLN: Pleon, CAP: Carapace, ISC: Ischium, ATN: Antenna, RST: Rostrum, BS-ISC: Basis-Ischium IM: Inner Margin, OM: Outer margin, and (B) Ventral, where TED: Tendon of external adductor muscle,TEB: Tendon of external abductor muscle, FLG: Flagellum, EXP: Exopod, EDP: Endopod, SPH: Scaphognathite, FLB: Flabellum, MP: Mandibullar palp, TLN: Telson, ANS: Anus.

Figure 6 Anatomy of Portunus pelagicus, (A) Dorsal, where DCT: Dactylus,PLX: Pollex, CRP: Carpus, MRS: Merus, BS: Basis, COX: Coxa, PPD: Propodus, PLN: Pleon, CAP: Carapace, ISC: Ischium, ATN: Antenna, RST: Rostrum, BS-ISC: Basis-Ischium IM: Inner Margin, OM: Outer margin, and (B) Ventral, where TED: Tendon of external adductor muscle,TEB: Tendon of external abductor muscle, FLG: Flagellum, EXP: Exopod, EDP: Endopod, SPH: Scaphognathite, FLB: Flabellum, MP: Mandibullar palp, TLN: Telson, ANS: Anus.
Shrimps (penaeidae)
The external morphology of the 3 penaeid shrimp species follows the typical decapod pattern, consisting of a fused cephalothorax and a segmented abdomen. Key external features, detailed in Figures 7 - 9, include the telson, protopod, endopod, exopod, and pereopods on the dorsal side, while the ventral side comprises structures like the pleopods, coxa, mandible, and various sensory appendages (e.g., scaphocerite, basicerite). Distinctive coloration aids in identification: the Tiger Shrimp (P. monodon) is characterized by prominent striping; the Dogol Shrimp (M. monoceros) typically exhibits a pale gray coloration with scattered dark brown spots; and the Jerbung Shrimp (P. merguiensis) has a yellowish-white base color, with brown and green spots on the carapace and a reddish tail tip.
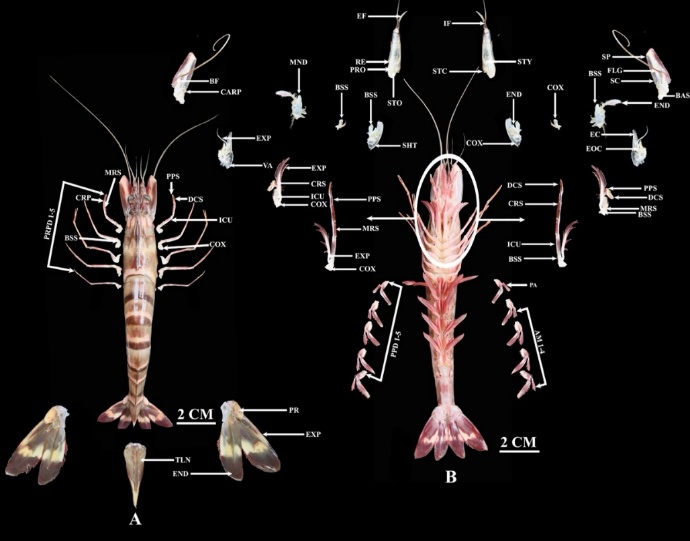
Figure 7 Anatomy of Penaeus monodon, (A) Dorsal, where PR: Protopod, TLS: Telson, END: Endopod, EXP: Exopod, PRPD: Pereopod, and (B) Ventral, where AM: Appendix masculina, PPD: Pleopod, COX: Coxa, EXP: Exopod, MRS: Merus, PPS: Propodus, ICU: Ischium, CRS: Carpus, VA: Vestigial arthrobranch, MND: Mandible, BSS: Basis, SHT: Scaphognathite, CARP: Carpocerite, BF: Base of flagellum, STO: Statocyst openin, RE: Recess for eye, PRO: Prosartema, EF: Recess for eye, IF: Inner flagellum, STC: Statocyst cover, STY: Stylocerite, SP: Spine, FLG: Flagellum, SC: Scaphocerite, BAS: Basicerite, EC: Exite of coxa, EOC: Endite of coxa, DCS: Dactyl.
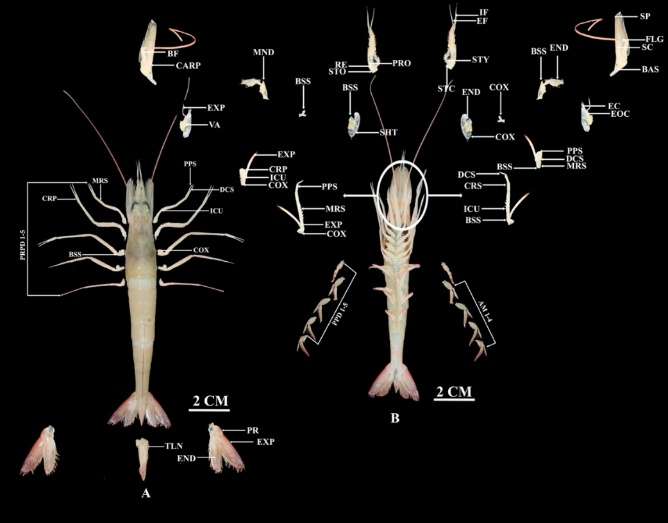
Figure 8 Anatomy of Metapenaeus monoceros, (A) Dorsal, where PR: Protopod, TLS: Telson, END: Endopod, EXP: Exopod, PRPD: Pereopod, and (B) Ventral, where AM: Appendix masculina, PPD: Pleopod, COX: Coxa, EXP: Exopod, MRS: Merus, PPS: Propodus, ICU: Ischium, CRS: Carpus, VA: Vestigial arthrobranch, MND: Mandible, BSS: Basis, SHT: Scaphognathite, CARP: Carpocerite, BF: Base of flagellum, STO: Statocyst openin, RE: Recess for eye, PRO: Prosartema, EF: Recess for eye, IF: Inner flagellum, STC: Statocyst cover, STY: Stylocerite, SP: Spine , FLG: Flagellum, SC: Scaphocerite, BAS: Basicerite, EC: Exite of coxa, EOC: Endite of coxa, DCS: Dactyl.
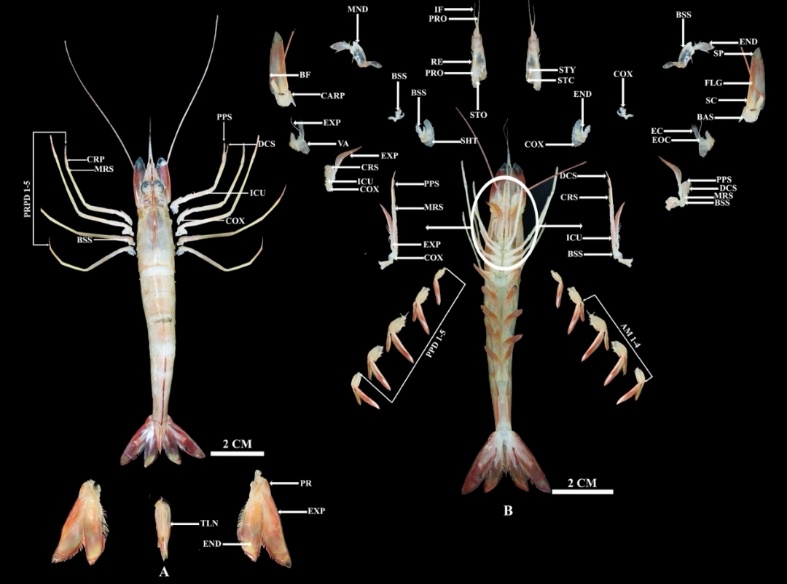
Figure 9 Anatomy of Penaeus merguensis, (A) dorsal, where PR: Protopod, TLS: Telson, END: Endopod, EXP: Exopod, PRPD: Pereopod, and (B) Ventral, where AM: Appendix masculina, PPD: Pleopod, COX: Coxa, EXP: Exopod, MRS: Merus, PPS: Propodus, ICU: Ischium, CRS: Carpus, VA: Vestigial arthrobranch, MND: Mandible, BSS: Basis, SHT: Scaphognathite, CARP: Carpocerite, BF: Base of flagellum, STO: Statocyst openin, RE: Recess for eye, PRO: Prosartema, EF: Recess for eye, IF: Inner flagellum, STC: Statocyst cover, STY: Stylocerite, SP: Spine , FLG: Flagellum, SC: Scaphocerite, BAS: Basicerite, EC: Exite of coxa, EOC: Endite of coxa, DCS: Dactyl.
Spiny lobsters (Palinuridae)
The lobsters identified, the Sand Lobster (P. homarus) and Pearl Lobster (P. ornatus), share a robust morphology (Figures 10 and 11). The head and thorax feature small spines on the carapace, and the abdomen displays distinct transverse segments. The Sand Lobster is typically greenish or brownish with white, sand-like spots, whereas the Pearl Lobster is recognized by its striking blue-green body color and distinct white and yellow speckled texture. Morphological structures common to both include the mandibular palp, processes (molar and incisor), various podites (carpopodite, ischiopodite, etc.), pleopod, and telson.
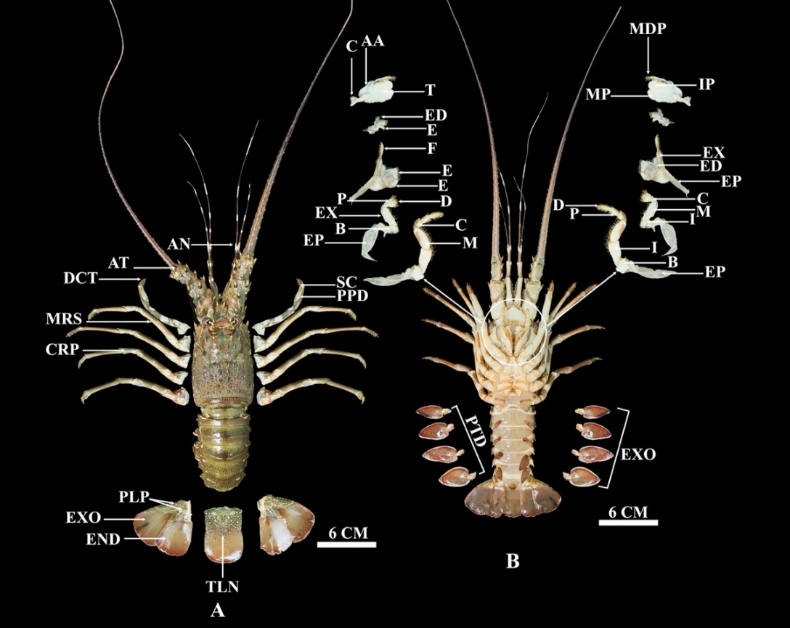
Figure 10 Anatomy of Panulirus homarus, (A) Dorsal, where MP: Molar process, MDP: Mandibular palp, IP: Incisor process, EX: Exopodite, ED: Endopodite, EP: Epipodite, C: Carpopodite, M: Meropodite, I: Ischiopodite, D: Dactylopodite, P: Propodite, I: ischiopodite, B: Basis, EP: Epipodite, EXO: Exopod, PTD: Protopod, E: Endite, T: Tubercles, AA: Anteror apodeme, C: Condyle, and (B) Ventral, where PPD: Propodus, SC: Subchela, AN: Antennulary, AT: Antennary, DCT: Dactylus, MRS: Merus, CRP: Carpus, PLP: Pleopod, EXO: Exopod, END: Endopod, TLN: Telson.
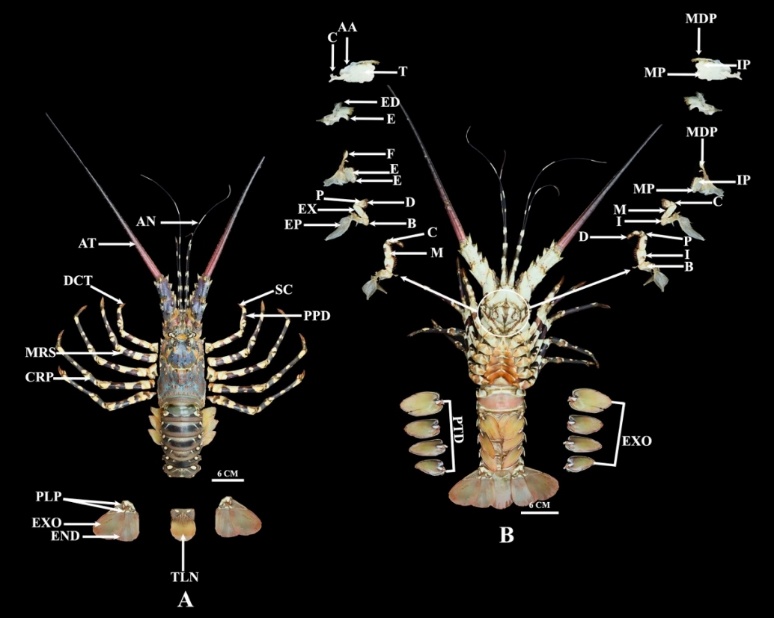
Figure 11 Anatomy of Panulirus ornatus, (A) Dorsal, where MP: Molar process, MDP: Mandibular palp, IP: Incisor process, EX: Exopodite, ED: Endopodite, EP: Epipodite, C: Carpopodite, M: Meropodite, I: Ischiopodite, D: Dactylopodite, P: Propodite, I: ischiopodite, B: Basis, EP: Epipodite, EXO: Exopod, PTD: Protopod, E: Endite, T: Tubercles, AA: Anteror apodeme, C: Condyle, and (B) Ventral, where PPD: Propodus, SC: Subchela, AN: Antennulary, AT: Antennary, DCT: Dactylus, MRS: Merus, CRP: Carpus, PLP: Pleopod, EXO: Exopod, END: Endopod, TLN: Telson.
Morphometric analysis of swimming crab species
The morphometric analysis was conducted on 3 commercially important decapod crab species: P. sanguinolentus, P. pelagicus, and C. feriata (n = 10 for each). The data, including the mean, range (min-max), and standard deviation (SD) for various carapace and appendage measurements, highlight significant interspecific differences in body dimensions and growth characteristics (Table 5).
Table 5 Morphometric analysis of swimming crabs collected from the east coast of Aceh Province, Indonesia
Parameters |
Portunus sanguinolentus (n = 10) |
Portunus pelagicus (n = 10) |
Charybdis feriata (n = 10) |
|||||||||
min |
max |
mean |
Standar deviation |
min |
max |
mean |
Standar Deviation |
min |
max |
mean |
Standar deviation |
|
(CW) |
6.47 |
9.81 |
8.12 |
1.05 |
9.24 |
12.37 |
10.64 |
1.07 |
9.23 |
11.85 |
10.95 |
0.90 |
(ICW) |
4.92 |
7.75 |
6.42 |
0.70 |
7.30 |
10.72 |
8.57 |
1.04 |
8.89 |
11.89 |
10.39 |
0.93 |
(CL) |
2.60 |
4.08 |
3.32 |
0.48 |
4.22 |
6.26 |
4.91 |
0.63 |
6.16 |
8.05 |
7.12 |
0.58 |
(LACL) |
2.63 |
4.08 |
3.45 |
0.46 |
3.81 |
5.31 |
4.42 |
0.47 |
3.14 |
5.17 |
4.41 |
0.66 |
(RACL) |
2.63 |
4.09 |
3.44 |
0.46 |
3.81 |
5.31 |
4.42 |
0.47 |
3.94 |
4.99 |
4.60 |
0.41 |
(LPCL) |
2.74 |
4.00 |
3.36 |
0.41 |
3.80 |
5.13 |
4.44 |
0.44 |
4.06 |
5.52 |
4.97 |
0.46 |
(RPCL) |
2.77 |
4.00 |
3.39 |
0.39 |
3.80 |
5.12 |
4.45 |
0.44 |
4.08 |
5.58 |
4.94 |
0.46 |
(FRMW) |
0.25 |
1.70 |
1.12 |
0.49 |
1.68 |
2.46 |
1.93 |
0.22 |
2.95 |
3.85 |
3.29 |
0.36 |
LOW |
0.32 |
0.77 |
0.48 |
0.11 |
0.38 |
0.70 |
0.50 |
0.10 |
0.27 |
0.91 |
0.58 |
0.21 |
(ROW) |
0.28 |
0.52 |
0.41 |
0.07 |
0.39 |
5.53 |
1.00 |
1.59 |
0.25 |
0.92 |
0.53 |
0.22 |
(PBW) |
1.13 |
2.91 |
2.29 |
0.45 |
2.82 |
4.72 |
3.48 |
0.57 |
3.12 |
3.68 |
3.49 |
0.16 |
(CHL2) |
6.52 |
10.77 |
7.87 |
1.20 |
8.28 |
12.81 |
10.55 |
1.47 |
10.18 |
23.88 |
18.46 |
5.54 |
(CHL1) |
6.12 |
10.93 |
7.62 |
1.20 |
8.48 |
12.29 |
10.25 |
1.37 |
10.27 |
24.36 |
19.04 |
5.67 |
(CHH2) |
0.42 |
0.98 |
0.77 |
0.17 |
0.80 |
1.39 |
1.05 |
0.18 |
1.23 |
2.56 |
1.78 |
0.42 |
(CHH1) |
0.50 |
0.98 |
0.77 |
0.16 |
0.79 |
1.30 |
1.05 |
0.16 |
1.35 |
2.46 |
1.88 |
0.38 |
Weight |
10.06 |
48.27 |
28.55 |
9.91 |
52.29 |
122.80 |
77.70 |
23.69 |
156.46 |
341.15 |
268.33 |
76.84 |
The analysis of carapace dimensions established a distinct size hierarchy among the 3 crab species. The Carapace Width (CW) consistently showed C. feriata as the largest species, recording the highest mean value at 10.95±0.90 cm (range: 9.23 - 11.85 cm), closely followed by P. pelagicus (10.64 ± 1.07 cm; range: 9.24 - 12.37 cm). In contrast, P. sanguinolentus was significantly smaller, with a mean CW of only 8.12 ± 1.05 cm. This size trend was similarly reflected in the Carapace Length (CL) and the Interorbital Carapace Width (ICW), where C. feriata maintained the largest dimensions (7.12 ± 0.58 cm for CL and 10.39 ± 0.93 cm for ICW), underscoring its generally larger size profile compared to the Portunus species.
Measurements of the anterior and posterior carapace length (LACL, RACL, LPCL, RPCL) and the major chela (claw) dimensions revealed significant structural differences, particularly concerning predatory and defensive morphology. The Chela Length (CHL1) for C. feriata averaged 19.04 ± 5.67 cm, which was approximately double the mean length observed in P. pelagicus (10.25 ± 1.37 cm) and P. sanguinolentus (7.62 ± 1.20 cm). Furthermore, the high standard deviation in C. feriata’s chela measurements (e.g., 5.67 cm for CHL1) indicates substantial allometric variability in claw development within this species. Correspondingly, Chela Height (CHH1) was also greatest in C. feriata (1.88 ± 0.38 cm), suggesting a more robust and powerful claw structure when compared to the appendages of the 2 Portunus species.
The Body Weight data provided the clearest distinction in biomass among the species. The mean weight for C. feriata was the highest at 268.33 ± 76.84 g (range: 156.46 - 341.15 g), confirming its status as the heaviest species captured. This substantial size difference is further emphasized when compared to P. pelagicus, which averaged 77.70 ± 23.69 g, and P. sanguinolentus, which registered the lowest mean weight at 28.55 ± 9.91 g. Collectively, these morphometric results consistently establish that C. feriata is the most dimensionally extensive and heaviest species among the decapod crabs analyzed in the study.
Overall, the morphometric data consistently indicate that C. feriata is the largest and most morphologically robust species among the 3 analyzed crabs, while the dimensions of P. pelagicus and P. sanguinolentus are considerably smaller. Furthermore, the higher standard deviation values observed in the chela measurements of C. feriata (e.g., 5.67 cm for CHL1) suggest greater allometric variability in claw development within this species compared to the Portunus species.
Morphometric analysis of penaeid shrimp species
The morphometric analysis, utilizing ten individuals (n = 10) for each species, was conducted on 3 commercially important penaeid shrimps: P. monodon, M. monoceros, and P. merguiensis. The results demonstrate clear dimensional differences, with P. monodon consistently exhibiting the largest average measurements across most parameters, followed by P. merguiensis.
Body Weight served as the clearest indicator of size disparity. P. monodon (Tiger Shrimp) was the heaviest species, recording a mean weight of 82.67 ± 18.85 g (ranging from 61.10 - 126.65 g). This mean weight was more than double that of P. merguiensis (Jerbung Shrimp), which averaged 38.15 ± 13.16 g. The smallest species was M. monoceros (Dogol Shrimp), with a mean weight of only 18.6 ± 5.1 g, reflecting its generally smaller commercial size. This size hierarchy established by weight was supported by the Total Length of the Pleon (PTO), where P. monodon (23.89 ± 2.60 cm) and P. merguiensis (19.82 ± 5.62 cm) were significantly larger than M. monoceros (13.6 ± 3.3 cm) (Table 6).
Table 6 Morphometric analysis of shrimps collected from the east coast of Aceh Province, Indonesia
Parameters |
Penaeus monodon (n = 10) |
Metapenaeus monoceros (n = 10) |
Penaeus merguiensis (n = 10) |
|||||||||
min |
max |
mean |
Standar deviation |
min |
max |
mean |
Standar deviation |
min |
max |
mean |
Standar deviation |
|
(RST) |
3.88 |
6.07 |
4.92 |
0.58 |
2.0 |
4.1 |
3.0 |
0.7 |
3.09 |
7.93 |
4.00 |
1.03 |
(PK) |
6.90 |
9.43 |
8.08 |
0.66 |
3.5 |
6.1 |
4.9 |
0.7 |
4.91 |
12.42 |
6.15 |
1.54 |
(PRP) |
0.81 |
2.35 |
1.95 |
0.28 |
0.7 |
1.9 |
1.2 |
0.3 |
1.15 |
3.66 |
1.64 |
0.53 |
(PRD) |
1.50 |
2.39 |
1.78 |
0.22 |
0.5 |
1.6 |
1.1 |
0.3 |
1.04 |
2.84 |
1.58 |
0.40 |
(PRT) |
1.26 |
2.31 |
1.81 |
0.24 |
0.3 |
1.5 |
1.2 |
0.3 |
1.05 |
3.25 |
1.77 |
0.49 |
(PRE) |
1.34 |
2.10 |
1.66 |
0.24 |
0.9 |
1.9 |
1.3 |
0.3 |
1.14 |
3.70 |
1.67 |
0.53 |
(PRL) |
1.03 |
1.80 |
1.27 |
0.19 |
0.5 |
1.4 |
0.9 |
0.2 |
0.92 |
2.54 |
1.29 |
0.37 |
(PRN) |
1.99 |
2.74 |
2.37 |
0.21 |
0.9 |
1.8 |
1.4 |
0.3 |
1.78 |
4.09 |
2.20 |
0.49 |
(TLS) |
1.95 |
2.95 |
2.54 |
0.21 |
1.4 |
2.0 |
1.7 |
0.2 |
1.73 |
4.18 |
2.13 |
0.56 |
(PST) |
1.89 |
3.43 |
2.64 |
0.42 |
1.1 |
2.1 |
1.7 |
0.3 |
1.61 |
3.71 |
2.20 |
0.47 |
(PNL) |
1.92 |
4.20 |
3.22 |
0.61 |
0.5 |
3.8 |
1.8 |
1.2 |
0.46 |
2.65 |
1.50 |
0.81 |
(PAN) |
21.72 |
38.75 |
30.19 |
4.41 |
10.1 |
32.4 |
18.1 |
8.1 |
21.70 |
44.74 |
27.57 |
4.80 |
(PTO) |
20.90 |
33.42 |
23.89 |
2.60 |
7.3 |
20.2 |
13.6 |
3.3 |
7.30 |
39.09 |
19.82 |
5.62 |
(PSTDR) |
15.47 |
28.59 |
17.85 |
2.87 |
9.2 |
13.5 |
10.8 |
1.3 |
11.41 |
32.05 |
15.12 |
4.38 |
(PKB) |
5.93 |
8.79 |
7.04 |
0.76 |
2.8 |
4.8 |
3.9 |
0.5 |
0.65 |
10.50 |
4.97 |
1.71 |
(PPB) |
1.42 |
2.00 |
1.67 |
0.14 |
0.4 |
1.2 |
0.8 |
0.2 |
0.63 |
2.77 |
1.41 |
0.46 |
(PDB) |
0.91 |
1.27 |
1.10 |
0.09 |
0.3 |
1.4 |
0.6 |
0.2 |
0.58 |
2.28 |
0.90 |
0.35 |
(PTB) |
0.99 |
1.31 |
1.11 |
0.09 |
0.4 |
0.8 |
0.6 |
0.1 |
0.58 |
1.81 |
0.87 |
0.26 |
(PEB) |
1.09 |
1.51 |
1.22 |
0.09 |
0.4 |
1.2 |
0.7 |
0.2 |
0.69 |
2.26 |
0.99 |
0.33 |
(PLB) |
1.15 |
1.45 |
1.27 |
0.09 |
0.3 |
1.0 |
0.6 |
0.2 |
0.82 |
2.27 |
1.09 |
0.32 |
(PNB) |
1.20 |
2.01 |
1.73 |
0.17 |
0.6 |
1.4 |
1.1 |
0.2 |
1.28 |
3.39 |
1.65 |
0.44 |
(PE) |
3.10 |
4.43 |
3.85 |
0.33 |
1.5 |
3.3 |
2.3 |
0.4 |
0.51 |
6.09 |
3.12 |
0.93 |
(PTK) |
13.14 |
17.82 |
15.15 |
1.24 |
8.2 |
12.3 |
10.0 |
1.4 |
10.95 |
27.12 |
14.19 |
3.67 |
Weight |
61.10 |
126.65 |
82.67 |
18.85 |
10.6 |
29.6 |
18.6 |
5.1 |
21.11 |
62.44 |
38.15 |
13.16 |
Parameters related to the carapace and rostrum consistently highlighted the dominance of P. monodon. The Rostrum Length (RST) averaged 4.92 ± 0.58 cm for P. monodon, which was considerably greater than P. merguiensis (4.00 ± 1.03 cm) and M. monoceros (3.0 ± 0.7 cm). Similarly, the Carapace Length (PK) for P. monodon was the largest at 8.08 ± 0.66 cm. Notably, P. merguiensis displayed the highest variability across several key parameters, evidenced by its large Standard Deviation (SD) for PK (1.54 cm) and PTO (5.62 cm), suggesting a broader range of sizes captured or higher morphological plasticity compared to the tightly distributed measurements of P. monodon (SD for PK: 0.66 cm; SD for PTO: 2.60 cm).
Appendage measurements, such as the length of the pleopods (e.g., PRP, PRD, PRT) and the length of the Telson (TLS), generally followed the overall body size trend: P. monodon ≥ P. merguiensis ≥ M. monoceros. For instance, the Telson Length (TLS) averaged 2.54 ± 0.21 cm for P. monodon and 1.7 ± 0.2 cm for M. monoceros. Despite its smaller mean size, P. merguiensis exhibited notably high variability in most appendage measurements (e.g., 0.56 cm for TLS), similar to its larger carapace dimensions, while P. monodon showed the lowest variability across nearly all measured parameters, indicating a more uniform sample set.
Morphometric analysis of spiny lobster species
The morphometric analysis, utilizing ten individuals (n = 10) for both species, compared P. ornatus (Pearl Lobster) and P. homarus (Sand Lobster) based on various body, appendage, and caudal measurements. The data reveal notable differences, particularly in overall size and the symmetry of certain morphological features.
P. homarus consistently exhibited a larger mean size profile than P. ornatus. P. homarus recorded a significantly higher mean Body Weight at 218.88 ± 46.68 g (range: 94.08 - 273.91 g), substantially exceeding the mean weight of P. ornatus (177.40 ± 51.26 g). Similarly, the Total Length was greater for P. homarus (18.41 ± 2.53 cm) compared to P. ornatus (15.47 ± 1.50 cm). The higher standard deviation in the total length of P. homarus (2.53 cm) suggests greater dimensional variability within the sampled population of this species (Table 7).
Table 7 Morphometric analysis of lobsters collected from the east coast of Aceh Province, Indonesia.
Parameters |
Panulirus ornatus (n = 10) |
Panulirus homarus (n = 10) |
||||||
min |
max |
mean |
Standar deviation |
min |
max |
mean |
Standar deviation |
|
Weight (g) |
112.45 |
272.76 |
177.40 |
51.26 |
94.08 |
273.91 |
218.88 |
46.68 |
Total Length (cm) |
13.47 |
17.63 |
15.47 |
1.50 |
12.92 |
20.82 |
18.41 |
2.53 |
A (Anterior of the head) (mm) |
|
|
|
|
|
|
|
|
A1 |
2.57 |
4.01 |
3.09 |
0.45 |
2.68 |
4.32 |
3.89 |
0.53 |
A2 |
1.27 |
2.10 |
1.64 |
0.27 |
1.01 |
1.96 |
1.27 |
0.29 |
A3 |
2.57 |
4.02 |
3.09 |
0.45 |
2.68 |
4.32 |
3.89 |
0.53 |
A4 |
3.46 |
4.81 |
4.08 |
0.44 |
3.70 |
5.36 |
4.97 |
0.52 |
A5 |
3.21 |
4.98 |
3.95 |
0.61 |
3.20 |
5.30 |
4.54 |
0.54 |
A6 |
3.21 |
4.91 |
3.94 |
0.60 |
3.20 |
5.31 |
4.54 |
0.54 |
B (Posterior of the head) (mm) |
|
|
|
|
|
|
|
|
B1 |
2.93 |
4.00 |
3.44 |
0.41 |
2.54 |
4.84 |
3.84 |
0.66 |
B2 |
3.52 |
4.82 |
4.13 |
0.44 |
3.69 |
5.36 |
4.98 |
0.49 |
B3 |
2.93 |
4.00 |
3.43 |
0.39 |
2.54 |
4.84 |
3.84 |
0.66 |
B4 |
3.12 |
4.66 |
3.88 |
0.52 |
3.07 |
4.31 |
4.03 |
0.35 |
B5 |
4.47 |
6.09 |
5.27 |
0.55 |
4.40 |
5.96 |
5.55 |
0.45 |
B6 |
4.47 |
6.08 |
5.27 |
0.54 |
4.40 |
5.96 |
5.55 |
0.46 |
Body (C) (mm) |
|
|
|
|
|
|
|
|
C1 |
5.61 |
7.36 |
6.36 |
0.53 |
4.62 |
7.75 |
6.76 |
1.07 |
C2 |
3.07 |
4.20 |
3.64 |
0.37 |
2.87 |
4.31 |
3.98 |
0.40 |
C3 |
5.61 |
7.36 |
6.36 |
0.54 |
4.62 |
7.75 |
6.76 |
1.07 |
C4 |
1.92 |
2.97 |
2.45 |
0.33 |
1.86 |
3.68 |
2.66 |
0.47 |
C5 |
6.15 |
7.95 |
6.92 |
0.60 |
5.16 |
8.43 |
7.53 |
1.05 |
C6 |
6.15 |
7.83 |
6.91 |
0.57 |
5.16 |
8.43 |
7.53 |
1.05 |
Body Sections (R) (mm) |
|
|
|
|
|
|
|
|
R1 |
3.05 |
4.15 |
3.60 |
0.38 |
3.09 |
4.69 |
4.21 |
0.44 |
R2 |
2.76 |
4.23 |
3.46 |
0.42 |
2.92 |
4.62 |
4.15 |
0.49 |
R3 |
2.74 |
3.91 |
3.32 |
0.38 |
2.83 |
4.44 |
3.96 |
0.47 |
R4 |
2.56 |
3.76 |
3.16 |
0.39 |
2.50 |
4.17 |
3.67 |
0.50 |
R5 |
2.44 |
3.50 |
2.93 |
0.37 |
2.42 |
3.76 |
3.35 |
0.40 |
R6 |
2.33 |
3.19 |
2.71 |
0.31 |
2.39 |
3.66 |
3.33 |
0.41 |
Caudal (D) (mm) |
|
|
|
|
|
|
|
|
D1 |
2.14 |
2.87 |
2.45 |
0.27 |
1.57 |
3.64 |
2.92 |
0.77 |
D2 |
1.62 |
2.35 |
2.02 |
0.27 |
1.26 |
3.68 |
2.29 |
0.68 |
D3 |
2.10 |
2.85 |
2.45 |
0.28 |
1.57 |
3.64 |
2.92 |
0.77 |
D4 |
1.32 |
2.00 |
1.72 |
0.25 |
1.30 |
3.68 |
2.36 |
0.69 |
D5 |
2.59 |
3.48 |
2.93 |
0.29 |
2.42 |
4.30 |
3.62 |
0.79 |
D6 |
2.45 |
3.48 |
2.91 |
0.31 |
2.42 |
4.30 |
3.62 |
0.79 |
Leg (E: front legs to the bottom) (mm) |
|
|
|
|
|
|
|
|
E1 |
6.80 |
9.26 |
7.79 |
0.81 |
5.78 |
8.21 |
7.58 |
0.73 |
E2 |
3.66 |
6.08 |
5.00 |
0.68 |
3.97 |
10.27 |
7.66 |
2.37 |
E3 |
6.71 |
11.87 |
9.31 |
1.34 |
7.27 |
12.41 |
10.20 |
1.42 |
E4 |
6.68 |
10.54 |
8.48 |
1.10 |
6.17 |
10.45 |
9.27 |
1.28 |
E5 |
5.20 |
10.14 |
7.03 |
1.27 |
5.36 |
8.45 |
7.45 |
0.94 |
E1 (from top to bottom) (mm) |
|
|
|
|
|
|
|
|
E1, 1 |
0.90 |
2.36 |
1.65 |
0.53 |
1.35 |
3.71 |
2.68 |
0.94 |
E1, 2 |
0.88 |
1.43 |
1.13 |
0.18 |
0.93 |
1.99 |
1.33 |
0.28 |
E1, 3 |
1.19 |
1.84 |
1.42 |
0.18 |
1.00 |
1.94 |
1.61 |
0.32 |
E1, 4 |
0.44 |
0.83 |
0.64 |
0.15 |
0.70 |
1.39 |
1.17 |
0.25 |
E2 (from top to bottom) (mm) |
|
|
|
|
|
|
|
|
E2, 1 |
3.02 |
3.78 |
3.29 |
0.25 |
2.38 |
4.56 |
3.64 |
0.72 |
E2, 2 |
1.09 |
1.61 |
1.24 |
0.16 |
1.10 |
1.47 |
1.31 |
0.11 |
E2, 3 |
2.05 |
2.77 |
2.24 |
0.22 |
1.64 |
2.82 |
2.33 |
0.40 |
E2, 4 |
0.77 |
1.17 |
0.98 |
0.12 |
0.77 |
1.33 |
1.12 |
0.17 |
E3 (from top to bottom) (mm) |
|
|
|
|
|
|
|
|
E3, 1 |
3.69 |
4.83 |
4.14 |
0.37 |
3.20 |
4.71 |
4.24 |
0.44 |
E3, 2 |
1.15 |
1.70 |
1.35 |
0.17 |
1.21 |
1.64 |
1.43 |
0.13 |
E3, 3 |
2.43 |
3.52 |
2.81 |
0.34 |
2.08 |
3.70 |
3.02 |
0.44 |
E3, 4 |
1.15 |
1.41 |
1.26 |
0.09 |
1.05 |
1.78 |
1.42 |
0.21 |
E4 (from top to bottom) (mm) |
|
|
|
|
|
|
|
|
E4, 1 |
2.76 |
4.46 |
3.53 |
0.54 |
3.05 |
4.20 |
3.82 |
0.36 |
E4, 2 |
0.94 |
1.48 |
1.24 |
0.16 |
0.99 |
1.74 |
1.39 |
0.19 |
E4, 3 |
2.12 |
2.91 |
2.50 |
0.23 |
1.89 |
3.07 |
2.76 |
0.35 |
E4, 4 |
0.98 |
1.51 |
1.28 |
0.17 |
0.95 |
1.69 |
1.44 |
0.24 |
E5 (from top to bottom) (mm) |
|
|
|
|
|
|
|
|
E5, 1 |
2.34 |
3.71 |
2.84 |
0.39 |
2.21 |
3.14 |
2.84 |
0.26 |
E5, 2 |
1.01 |
1.33 |
1.15 |
0.11 |
0.99 |
1.42 |
1.29 |
0.13 |
E5, 3 |
1.80 |
2.92 |
2.26 |
0.32 |
1.72 |
2.69 |
2.44 |
0.30 |
E5, 4 |
0.59 |
1.27 |
0.93 |
0.26 |
0.79 |
1.47 |
1.06 |
0.20 |
Antena (mm) |
|
|
|
|
|
|
|
|
Left |
28.90 |
49.65 |
37.50 |
7.10 |
23.30 |
46.50 |
38.52 |
6.24 |
Right |
26.93 |
44.73 |
35.92 |
6.69 |
22.50 |
46.10 |
36.91 |
8.57 |
Measurements across the head (A,B) and body sections (C,R) generally supported the size hierarchy, with P. homarus often showing slightly larger mean values. For instance, the posterior head measurements (B2) averaged 4.98 ± 0.49 mm for P. homarus compared to 4.13 ± 0.44 mm for P. ornatus. Similarly, body section measurements (C2) were larger in P. homarus (3.98 ± 0.40 mm) than in P. ornatus (3.64 ± 0.37 mm). Notably, the caudal (D) dimensions displayed a particularly high degree of variability in P. homarus. Parameters such as D1 (mean 2.92 ± 0.77 mm) and D5 (mean 3.62 ± 0.79 mm) showed standard deviations approximately 3 times higher than those recorded for P. ornatus (e.g., 0.27 mm for D1; 0.29 mm for D5). This indicates a marked heterogeneity in the caudal structure size of P. homarus.
Measurements of the pereopods (Leg E) revealed complex differences. While P. homarus showed a larger mean for several proximal segments (e.g., E2 mean 7.66 ± 2.37 vs. 5.00 ± 0.68 mm for P. ornatus), P. ornatus displayed greater means in others. Importantly, the E2 measurement in P. homarus exhibited a highly substantial standard deviation (SD = 2.37), the largest variation observed across all limb parameters, reflecting significant allometric or sexual differences in this particular appendage.
Regarding Antenna Length, P. homarus possessed slightly longer mean left and right antennae (38.52 ± 6.24 and 36.91 ± 8.57 mm, respectively) compared to P. ornatus. Both species showed some degree of asymmetry between the left and right antennae, though the difference in the right antenna measurement for P. homarus (SD = 8.57) suggests greater variability in this feature than in P. ornatus (SD = 6.69).
In conclusion, the morphometric results confirm that P. homarus is generally the larger and heavier species. Furthermore, it exhibits significantly higher morphological heterogeneity in its caudal section and certain appendage segments compared to the more uniformly sized P. ornatus.
Discussion
The Decapoda Crustacean group represents a globally significant component of marine biodiversity, comprising an immense 17,229 recorded species spanning 2,550 genera and 203 families [24]. Recognized as the taxon with the largest number of species [25], decapods are critically important for both marine ecology and global fisheries. Locally, this research focused on the eastern coast of Aceh Province, Indonesia, identified 8 commercially relevant decapod species: C. feriata, P. sanguinolentus, P. pelagicus, P. monodon, M. monoceros, P. merguiensis, P. homarus, and P. ornatus. However, our findings reveal distinct ecological patterns and morphological variations that reflect the local environmental conditions of the Malacca Strait. Given the ecological and economic significance of these taxa, the conservation of their natural habitats is imperative in Indonesia to ensure sustainable resource utilization [26].
The family Portunidae includes highly valuable resources such as the blue swimming crab (P pelagicus), an important organism globally [27] and a key fishery commodity in Indonesia [28]. Similarly, the coral crab (C. feriata) is a major economic contributor to fishermen worldwide [29]. The widespread distribution of P. pelagicus spans 17 Provinces and 10 Fisheries Management Areas (WPP) across Indonesia [30], often appearing as bycatch in regions like South Sumatra [31], and Southeast Sulawesi [32]. The management of P. sanguinolentus, however, is increasingly critical, as its population is declining due to intensive exploitation pressure and habitat destruction [33,34]. Detailed morphological analysis is foundational for accurate stock assessment. The general external anatomy of these crab species includes characteristic dorsal features such as the carapace, pleon, dactylus, and rostrum, while the ventral structure is defined by muscles (tendons of external adductor/abductor), appendages (exopod, endopod, flagellum), and the telson. Such structural details are directly linked to morphometric studies, particularly those correlating carapace width with body weight, which is essential for defining management parameters [35]. By correlating our morphometric data - specifically carapace width to body weight - we provide the foundational parameters necessary for local fisheries authorities to establish sustainable catch quotas.
Interestingly, the C. feriata was recorded exclusively in the Pidie sampling site. This spatially restricted occurrence may be attributed to habitat specificity and the unique seafloor topography of Pidie, which features more prominent rocky and coral reef patches compared to the predominantly muddy estuaries of North Aceh and Aceh Tamiang. C. feriata is known to prefer reef-associated or rocky substrates [29]. Additionally, this distribution may be influenced by gear selectivity; the traditional bottom gillnets and traps (bubu) used by Pidie fishermen are specifically deployed in these rocky corridors, whereas the trawling and push-nets common in other regencies are less suited for capturing this species. This localized presence suggests that Pidie serves as a critical habitat for C. feriata in Aceh, necessitating specialized management of its rocky-coastal ecosystems.
The family Penaeidae encompasses some of the most economically and ecologically important marine shrimps globally [36]. Ecological and morphometric studies are vital for gathering information on population structure, growth rates, and morphological relationships within these species [37]. The presence of the decapod Penaeus species is widely confirmed across marine waters worldwide [38]. P. monodon (Tiger Prawn), a native Indonesian species, commands high economic value and offers enormous potential for fishery products [39]. Other key catches include M. monoceros, a commercially important penaeid shrimp in the Indo-West Pacific [40,41], and P. merguiensis, which is a primary commercial target along the coastal areas of Sumatra, Indonesia, possessing high economic and ecological value [42,43]. The dorsal morphology of these penaeid species is characterized by the protopod, telson, exopod, and pereopods. The ventral section is complex, featuring the masculine appendix, pleopods, mandibles, and specialized sensory structures like the statocyst opening and scaphocerite.
Lobsters (Panulirus spp.) represent another high-value fishery resource, with the majority of catches derived from marine environments [44]. Aceh Province is known to harbor several species, including P. homarus and P. ornatus [22]. The Pearl Lobster (P. ornatus) has a wide distribution across the Indo-West Pacific and is recognized as one of the world's most valuable seafood products [45]. Conversely, the Sand Lobster (P. homarus) is characterized by a greenish or brownish body color with sand-like white spots, small carapace spines, and segmented abdomen [46]. Morphological features, such as antenna shape and body pattern color, serve as significant distinguishing characteristics between the 2 species [47]. The dorsal morphology of Panulirus species is defined by structures like the mandibular palp, incisor process, and various podites (e.g., carpopodite, ischiopodite). Ventrally, key features include the propodus, subchela, pleopod, and telson. Morphological measurements provide essential data for the accurate characterization of Panulirus stocks [48]. To ensure resource sustainability and expand scientific understanding, continuous monitoring and further research in this area are necessary [49].
Notable morphological variability was observed in P. homarus across the sampling sites. This variability, particularly in carapace coloration and spine density, likely reflects adaptations to the diverse benthic substrates found between Pidie and Aceh Tamiang. The eastern coast of Aceh is characterized by a gradient of sandy-muddy bottoms influenced by riverine discharge. Such environmental heterogeneity, combined with varying levels of fishing pressure, may drive localized phenotypic plasticity or population structuring. In areas with higher turbidity, P. homarus exhibited darker pigmentation, a trait potentially linked to camouflage and survival in low-visibility environments. Furthermore, the morphometric range observed suggests that the population consists of multiple age cohorts, highlighting the need for site-specific size limits to prevent growth overfishing.
Conclusions
This study provides foundational morphological and morphometric baselines for the decapod crustacean resources along the eastern coast of Aceh Province, Indonesia, identifying 8 commercially important species across the Portunidae, Penaeidae, and Palinuridae families. The morphometric analysis successfully established a clear size hierarchy, positioning C. feriata, P. monodon, and P. homarus as the largest species within their respective groups based on mean body weight and carapace dimensions. Furthermore, the high standard deviation observed in key measurements for species like P. merguiensis and P. homarus highlights significant morphological heterogeneity, suggesting potential variations in allometric growth or local environmental influences within these stocks. Given the ecological and economic significance of these species - particularly the evident exploitation pressure on taxa such as P. sanguinolentus - the data from this research are indispensable for informing species-specific management strategies, monitoring growth rates, and establishing sustainable resource utilization protocols necessary to mitigate the threats posed by unregulated harvesting.
Acknowledgements
The authors would like to thank the Ministry of Higher Education, Science, and Technology of Indonesia for funding this research through the Fundamental-Regular Research scheme (Number: 134/C3/DT.05.00/PL/2025, 29/LL13/AL.04/AKA.PL/2025, 255/LPPM-Umuslim/KP-PFR/2025).
Conflict of interest declare
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Declaration of Generative AI in Scientific Writing
This manuscript utilized generative AI tools, namely QuillBot and Grammarly, to enhance language clarity, grammar, and overall readability.
CRediT author statement
Irfannur Irfannur: Investigation, Methodology, Funding acquisition, Conceptualization, Writing - original draft, Writing - review & editing. Agung Setia Batubara: Investigation, Writing - review & editing, Writing - original draft. Siti Annisa Ramadhani: Investigation, Writing - original draft. Muliari Muliari: Investigation, Formal analysis, Writing - review & editing. Muhammad Radhi: Investigation, Methodology, Writing - original draft. Yusrizal Akmal: Investigation, Methodology, Writing - original draft. Ilham Zulfahmi: Investigation, Formal analysis, Writing - review & editing.
References
IJ Jensen, N Bodin, R Govinden and EO Elvevoll. Marine capture fisheries from western Indian ocean: an excellent source of proteins and essential amino acids. Foods 2023; 12, 1015.
MT Smith and JT Addison. Methods for stock assessment of Crustacean fisheries. Fisheries Research 2003; 65(1-3), 231-256.
JM Mafuca, RJ Mutombene, O Filipe, S Abdula, BS Malauene, N Dias, RA Jubb Quick, S Fennessy, B Everett and M Roberts. Planning for climate change resilience - Collation, update and assessment of Mozambique’s marine fisheries data and management. PloS Climate 2024; 3(10), e0000494.
MG Bondad-Reantaso, RP Subasinghe, H Josupeit, J Cai and X Zhou. The role of crustacean fisheries and aquaculture in global food security: Past, present and future. Journal of Invertebrate Pathology 2012; 110, 158-165.
CE Boyd, AA McNevin and RP Davis. The contribution of fisheries and aquaculture to the global protein supply. Food Security 2022; 14, 805-827.
DC Behringer and E Duermit-Moreau. Crustaceans, one health and the changing ocean. Journal of Invertebrate Pathology 2021; 186, 107500.
P Briones-Fourzán and ME Hendrickx. Ecology and diversity of marine decapod crustaceans. Diversity 2022; 14, 614.
N Sammy, De Grave, ST Ahyong, TY Chan, KA Crandall, PC Dworschak, DL Felder, RM Feldmann, CHM Fransen, LYD Goulding, R Lemaitre, MEY Low, JW Martin, PKL Ng, C Schweitzer, SH Tan, D Tshudy and R Wetzer. A classification of living and fossil genera of decapod crustaceans. Raffles Bulletin of Zoology 2009; 21, 1-109.
RT Bauer. The families of decapod shrimps. In: RT Bauer (Ed.). Shrimps: Their diversity, intriguing adaptations and varied lifestyles. Springer, Cham, Switzerland, 2023, p. 69-194.
R Boenish, JP Kritzer, K Kleisner, RS Steneck, KM Werner, W Zhu, F Schram, D Rader, W Cheung, J Ingles, Y Tian and J Mimikakis. The global rise of crustacean fisheries. Frontiers in Ecology and the Environment 2022; 20, 102-110.
A Albalat, S Zacarias, CJ Coates, DM Neil and SR Planellas. Welfare in farmed decapod crustaceans, with particular reference to Penaeus vannamei. Frontiers in Marine Science 2022; 9, 886024.
DR Lawa, Y Jasmanindar and Y Linggi. Krustasea potensial untuk budidaya dari perairan Panmuti Kecamatan Kupang Tengah, Kabupaten Kupang (in Indonesian). Jurnal Vokasi Ilmu-Ilmu Perikanan 2024; 5, 36-41.
M Jamil, A Putriningtias, S Anzhita, H Akbar, S Syahrial, H Hanisah, TM Faisal and S Sinaga. Kepiting genera Scylla di pesisir Kota Langsa: Distribusi, dimensi tangkapan alami dan analisis bioekonominya (in Indonesian). Jurnal Kelautan Tropis 2023; 26, 357-368.
F Conte, E Voslarova, V Vecerek, RW Elwood, P Coluccio, M Pugliese and A Passantino. Humane slaughter of edible decapod crustaceans. Animals 2021; 11, 1089.
RW Elwood and RW Ingle. Decapod crustaceans. In: H Golledge and C Richardson (Eds.). The UFAW Handbook on the care and management of laboratory and other research animals. Wiley, New Jersey, 2024, p. 991-1011.
BG Stevens. The ups and downs of traps: Environmental impacts, entanglement, mitigation, and the future of trap fishing for crustaceans and fish. ICES Journal of Marine Science 2021; 78, 584-596.
MA Saputra. Moving within and beyond illegal crustacean fishery: Why do Indonesian fishermen not comply with the crustacean catch ban rule? Maritime Studies 2020; 19, 457-473.
DD Kembaren, P Lestari and R Ramadhani. Parameter biologi lobster pasir (Panulirus homarus) di perairan Tabanan, Bali (in Indonesian). BAWAL Widya Riset Perikanan Tangkap 2015; 7, 35-42.
R Lagarde, N Teichert, P Valade and D Ponton. Structure of small tropical island freshwater fish and crustacean communities: A niche‐or dispersal‐based process? Biotropica 2021; 53, 243-254.
P Borsa, DR Hsiao, KE Carpenter and WJ Chen. Cranial morphometrics and mitochondrial DNA sequences distinguish cryptic species of the longface emperor (Lethrinus olivaceus), an emblematic fish of Indo-West Pacific coral reefs. Comptes Rendus Biologies 2013; 336, 505-514.
A Pérez-Olivas, MS Irigoyen-Arredondo, XG Moreno-Sánchez, MT Villalejo-Fuerte, LA Abitia-Cárdenas and O Escobar-Sánchez. Reproductive biology of the leopard grouper Mycteroperca rosacea (Streets, 1877) in the coastal area of Santa Rosalía, BCS, Mexico. Latin American Journal of Aquatic Research 2018; 46, 699-708.
I Irfannur, S Saputra, M Muliari, Y Akmal and AS Batubara. Characterization of 6 lobster species of genus Panulirus (Decapoda, Palinuridae) from Aceh Waters, Indonesia based on morphometric analysis. Zoodiversity 2024; 58(2), 89-102.
AS Batubara, ZA Muchlisin, D Efizon, R Elvyra, N Fadli and M Irham. Morphometric variations of the genus Barbonymus (Pisces, Cyprinidae) harvested from Aceh waters, Indonesia. Fisheries & Aquatic Life 2018; 26, 231-237.
S De Grave, W Decock, S Dekeyzer, PJ Davie, C Fransen, CB Boyko, GCB Poore, E Macpherson, ST Ahyong, KA Crandall, V de Mazancourt, M Osawa, TY Chan, PKL Ng, R Lemaitre, SET van der Meij and S Santos. Benchmarking global biodiversity of decapod crustaceans (Crustacea: Decapoda). Journal of Crustacean Biology 2023; 43(3), ruad042.
J Jarquín-González, M Valdez-Moreno and R Rosas-Luis. First southern Mexican Caribbean Sea record of the Indo-Pacific crab Plagusia immaculata Lamarck, 1818 (Decapoda: Brachyura: Plagusiidae). Journal of Crustacean Biology 2025; 45(2), ruaf039.
DC Murniati. The diversity and distribution of Gecarcinidae MacLeay, 1838 (Crustacea: Decapoda: Brachyura), a potential land crab as a valuable food resource, in Indonesia. IOP Conference Series: Earth and Environmental Science 2023; 1275(1), 012038.
AG Dvoretsky and VG Dvoretsky. Epibiotic communities of common crab species in the coastal Barents Sea: Biodiversity and infestation patterns. Diversity 2021; 14(1), 6.
MI Joesidawati, N Nursalim, N Kholilah, EM Kurniasih, NKD Cahyani and Ambariyanto. Utilizing DNA barcoding approach to study the diversity of the blue swimming crab from Tuban District, East Java, Indonesia. Biodiversitas Journal of Biological Diversity 2023; 24(9), 4731-4737.
J He, J Wu, L Wan, W Xu and T Yang. Genome-wide genetic diversity and population structure of Charybdis feriata (Crustacea, Decapoda, and Portunidae) along the southeast coast of China inferred from Genotyping-by-Sequencing (GBS) approach. Genes 2024; 15, 1421.
H Madduppa, LMI Sani, KC Nugroho, DG Bengen, ZA Muchlisin and N Fadli. eDNA metabarcoding of decapod crustaceans across Indonesian seas has implications for biodiversity conservation and fisheries sustainability. Frontiers in Marine Science 2022; 9, 918295.
AIS Purwiyanto. Discarded species in artisanal fisheries South Sumatra, Indonesia: Case study on crab gill nets. Sains Malaysiana 2022; 51(9), 2745-2756.
A Hamid and S Kamri. Bycatch biodiversity of blue swimming crab (Portunus pelagicus) fisheries in Kolono Bay, Southeast Sulawesi, Indonesia. AACL Bioflux 2021; 14, 1548-1560.
K Mustabiq, N Fairuz, D Darmawan, A Ervinia, KC Nugroho and W Setioko. Population dynamics of the 3-spot swimming crab (Portunus sanguinolentus) in Pati waters, Central Java. Jurnal Ilmu dan Teknologi Kelautan Tropis 2023; 15(3), 301-308.
N Onsri, I Sivaipram, P Boonsanit, K Sagulsawasdipan and S Saramul. Larval dispersal modelling of the blue swimming crab Portunus pelagicus (Linnaeus, 1758) from the Crab Banks along the coast of Trang Province, Southern Thailand. Water 2024; 16(2), 349.
KY Hira, I Pratikto and I Riniatsih. Kajian morfometri rajungan (Portunus pelagicus) di perairan Kabupaten Pemalang Jawa Tengah. Journal of Marine Research 2024; 13(1), 73-82.
JL Ramirez, L Simbine, CG Marques, E Zelada-Mázmela, LE Reyes-Flores, AS Lopez, J Gusmao, C Tavares, PM Galetti and PD Freitas. DNA barcoding of penaeidae (Decapoda; Crustacea): Non-distance-based species delimitation of the most economically important shrimp family. Diversity 2021; 13(10), 460.
R Pratiwi and S Sukardjo. The ecological studies of shrimp Penaeus monodon Fabricius, 1798 (Penaeidae) in the Segara Anakan Lagoon (SAL), Cilacap, Central Java, Indonesia. IOP Conference Series: Earth and Environmental Science 2021; 789(1), 012008.
S Sadek, WA El-Soud and BS Galil. The brown shrimp Penaeus aztecus Ives, 1891 (Crustacea, Decapoda, Penaeidae) in the Nile Delta, Egypt: an exploitable resource for fishery and mariculture. Bioinvasions Records 2018; 7, 51-54.
N Fauziah, MB Mulya and TA Barus. Density, length-frequency distribution and growth pattern of Penaeus monodon in Karang Gading estuary. AACL Bioflux 2022; 15, 1422-1434.
MM Alam and S Pálsson. Mitochondrial DNA variation of Metapenaeus monoceros (Decapoda, Penaeidae) reveals high genetic variation within Bangladesh with distinct lineages in the Indian Ocean. Marine Ecology 2021; 42, e12653.
RT Pradana and A Abdunnur. Population dynamics of yellow prawn (Metapenaeus monoceros) captured at night in the waters of Samboja Kuala, Kutai Kartanegara Regency. Nusantara Tropical Fisheries Science Journal 2023; 2, 113-121.
A Hadi, DA Putri and L Fitrahsaeri. Analysis identification of composition catches Bubu Naga operated in Cangkring pond waters, Indramayu Regency, West Java. Barakuda 45: Jurnal Ilmu Perikanan dan Kelautan 2022; 4, 281-289.
I Widiani, TA Barus and H Wahyuningsih. Population of white shrimp (Penaeus Merguiensis) in a mangrove ecosystem, Belawan, North Sumatra, Indonesia. Biodiversitas: Journal of Biological Diversity 2021; 22(12), 5367-5374.
A Setyanto, MA Rahman, Y Fatmasari, VB Irawan, A Isdianto, MA Asadi, K Marsela, LA Dhea, NR Caesar and I Jatmiko. Species composition and length-weight frequency distribution of lobsters (Panulirus spp.) landed on the Puger Coast, Jember Regency, East Java. Berkala Sainstek 2025; 13(2), 96-108.
A Farhadi, F Pichlmueller, B Yellapu, S Lavery and A Jeffs. Genome-wide SNPs reveal fine-scale genetic structure in ornate spiny lobster Panulirus ornatus throughout Indo-West Pacific Ocean. ICES Journal of Marine Science 2022; 79(6), 1931-1941.
Y Akmal, I Irfannur, M Muliari, AS Batubara, M Yunus, H Plumeriastuti and Y Dhamayanti. A comprehensive description of the exoskeleton of 6 lobster species (genus Panulirus) in Aceh Province, Indonesia. Fisheries Research 2023; 264, 106731.
I Saputra and B Priyambodo. The development of lobster puerulus (Panulirus ornatus and P. homarus) in captivity environment. Indonesian Aquaculture Journal 2023; 18, 169-177.
CL Lewis, QP Fitzgibbon, GG Smith, A Elizur and T Ventura. Transcriptomic analysis and time to hatch visual prediction of embryo development in the ornate spiny lobster (Panulirus ornatus). Frontiers in Marine Science 2022; 9, 889317.
B Almón and R Bañón. Research trends and state of knowledge of decapod crustaceans in Spain: A bibliometric analysis. Ecologies 2025; 6, 63.