Trends
Sci.
2026; 23(8): 13647
Review on Coumarins from the Genus Calophyllum: Molecular Interactions Against HIV Targets and Insights from Computational and Experimental Studies
Nur Nabilah Mohd Zaini1, Wan Mohd Nuzul Hakimi Wan Salleh1,*, Abubakar Siddiq Salihu2, Nadtanet Nunthaboot3, Nurunajah Ab Ghani4,5, Farkhod Eshboev6 and Alfred Ngenge Tamfu7
1Department of Chemistry, Faculty of Science and Mathematics, Universiti Pendidikan Sultan Idris,
Tanjong Malim, Perak, Malaysia
2Department of Pure and Industrial Chemistry, Faculty of Natural and Applied Sciences,
Umaru Musa Yar’adua University, Katsina, Nigeria
3Chemistry Department, Faculty of Science, Mahasarakham University, Maha Sarakham, Thailand
4Atta-ur-Rahman Institute for Natural Product Discovery (AuRIns), Universiti Teknologi MARA,
Puncak Alam Campus, Selangor, Malaysia
5Faculty of Applied Sciences, Universiti Teknologi MARA, Shah Alam, Selangor, Malaysia
6Institute for Advanced Studies, New Uzbekistan University, Tashkent, Uzbekistan
7Department of Chemical Engineering, School of Chemical Engineering and Mineral Industries,
University of Ngaoundere, Ngaoundere, Cameroon
(*Corresponding author’s e-mail: [email protected])
Received: 18 February 2026, Revised: 12 March 2026, Accepted: 22 March 2026, Published: 5 April 2026
Abstract
The Calophyllum genus is a diverse group of tropical trees known for producing a wide variety of bioactive compounds, including xanthones, coumarins, triterpenoids, and flavonoids. Coumarins possess several pharmacological activities such as anti-HIV, anti-inflammatory, anti-bacterial, and anti-coagulation effects. Calanolide A, Calanolide B, and several Inophyllum derivatives are among the compounds that exhibit a significant inhibition of HIV-1 reverse transcriptase (RT) that is beyond the action of some HIV-1 RT inhibitors in market drugs. This research conducted the docking of HIV-1 RT and integrase (IN) to assess its antiviral coumarins derived from Calophyllum species. Coumarins are benzopyrones characterized by a conjugated aromatic system that is crucial for anchoring at the RT- NNRTI binding pocket and the IN catalytic core. Hydroxyl, methoxy, and acetoxy groups increase their binding affinity for enzymes and specificity. The molecular docking scores support these findings, where Inophyllum E, Soulattrolone, and other compounds show significantly better binding with RT and IN than the controls Efavirenz and Raltegravir. The impressive effectiveness can be attributed to their structural variety, hydrophobic interactions, and optimally placed functional groups, which allow for considerable and stable complexing within the active sites. Consequently, the Calophyllum coumarins are deemed valuable for the future development of novel antiviral drugs.
Keywords: Calophyllaceae, Calophyllum, Coumarin, Molecular docking, ADMET, Anti-HIV, Calanolides
Introduction
Calophyllum is a widely recognized genus consisting of over 200 species of shrubs and evergreen trees and has ecological and medicinal value. These tropical trees are found all over the regions of Asia, Africa, the Pacific Islands, and America, which are usually grown in mangroves, lowland forests, and coastal areas. Morphologically, Calophyllum species are characterized by glossy, leathery leaves, round-shaped fruits, and resinous latex, which have been used for various purposes and in traditional medicine for ages. The timber is also used in construction and furniture owing to its strength and beauty in its aesthetic value [1,2].
Calophyllum species were cultivated to produce different bioactive compounds, including xanthones, coumarins, triterpenoids, and flavonoids. These chemicals have different uses in medicine. However, researchers have pointed out that coumarins have significant antiviral, antibacterial, anti-inflammatory, and anticancer properties [3-5]. Coumarins, mostly located in the leaves, bark, and seeds of these plants, have been the subject of both modern pharmacological research and traditional medicine. For instance, C. inophyllum species plants, known in the local community as "Bintangor Laut,” have properties for healing wounds, burns, and skin infections [6]. Additionally, the seeds have skin-healing properties and reduce scarring.
Recent studies have validated the medicinal properties of several Calophyllum coumarins. One of the most innovative prototypes in anti-AIDS research, Calanolide A (2), was identified back in 1992 from C. lanigerum var. austrocoriaceum and has been proven to inhibit Human Immunodeficiency Virus-1 (HIV-1) reverse transcriptase (RT) [7]. This antiviral activity of HIV-1 RT stems from its dependency on non-nucleoside reverse transcriptase inhibitors (NNRTIs) and is known to inhibit HIV-1 RT severely. Its structure with the hydrophobic benzopyran core and other substituent groups has bested the resonance cavities of RT, thus calanolides have turned the enzyme HIV replication into a dead-end pathway. This has positioned Calanolide A (2) to be sought for further development as an NNRTI and has brought attention to Calophyllum species to be further explored for their compounds as a novel antiviral framework [8,9].
HIV is a specialized type of retrovirus that breaks into the immune system by infecting CD4⁺ T-cells, which slowly leads to Acquired Immunodeficiency Syndrome (AIDS), if untreated. The virus propagates by assimilating its genome into a host cell DNA sequence through the actions of RT, IN, and other viral enzymes. With the introduction of antiretroviral therapy (ART), like NNRTIs, protease (PR) and integrase (IN) inhibitors, it has significantly extended the lifespan of people with HIV [10]. However, issues like drug resistance and the presence of dormant viral reservoirs demand constant evolution of treatment techniques [11,12]. One avenue of cross-disciplinary research employs monoclonal antibodies, therapeutic vaccines, natural compounds derived from plants, and gene-editing technologies such as CRISPR/Cas9, aimed at deeper suppression of vital HIV proteins and scaffolding for future drugs. Although a definitive treatment standard remains elusive, the objective transitions to achieving a conclusive cure. However, there is optimism that subsequent advancements in research, together with future technological innovations, will enable the permanent inhibition or inactivation of the virus [13-15].
Considering this, there is growing interest in finding small molecules that can specifically target viral enzymes. Natural products are especially interesting for this goal because they exhibit a wide range of structures and their biological activities are well known. Computational methods, particularly molecular docking, provide an effective method to anticipate the interactions of these drugs with essential viral proteins and to rank the most promising candidates before experimental validation. These in silico techniques allow researchers to better understand the different parts of the NNRTI-binding pocket and identify the most promising ones to explore further. The integration of docking studies with conventional methodologies and experimental assays has augmented the understanding of Calophyllum coumarins as potential antiviral agents, thereby establishing the Calophyllum genus as a central focus for the development of innovative pharmaceuticals and therapeutics.
Coumarins are widespread in nature and appear in many plant families; species of Calophyllum are a well-known example. Their parent framework, C₉H₆O₂, belongs to the benzo-α-pyrone group, or sometimes referred to as 1,2-benzopyrones. Chemically, a coumarin arises when a benzene ring fuses with a pyrone ring carrying a carbonyl group. This union creates the lactone skeleton that defines the compound’s reactivity and much of its biological behavior [16]. With minor structural modifications such as a methyl or hydroxyl substituent at different ring positions, many related molecules are generated, with each showing its own physical or pharmacological character [17,18]. Researchers usually classify plant-derived coumarins by their core structure: simple, prenylated, geranylated, pyrano, furano, sesquiterpenyl, oligomeric, and a few miscellaneous coumarins, as summarized in Figure 1 [19,20]. These structural groups account for much of the biological activity observed in Calophyllum extracts. Historically, the word “Coumarin” itself comes from the French “Coumarou” the old name for the Tonka bean (Dipteryx odorata), where Vogel first isolated the compound in 1820 [19].

Figure 1 Chemical structures of isolated coumarins.
Coumarins have been of interest for many years since they smell pleasant and have a wide range of medicinal uses [17]. Extracts from Calophyllum species exhibit intricate molecular patterns and are frequently employed as chemical identifiers for the genus [21]. Among them, calanolides and inophyllums stand out for their significant antiviral and anticancer actions. Several dipyrano-tetracyclic coumarins, such as calanolides, inophyllums, and cordatolides, also inhibit HIV through the NNRTI site of the virus [22]. The coexistence of these metabolites makes Calophyllum a promising source of new therapeutic candidates. In traditional healing, Calophyllum preparations have long been used to relieve inflammation and infections, and modern pharmacological work has confirmed many of those early observations. Demonstrated antiviral, anticancer, and anti-inflammatory effects now provide scientific backing for their ethnomedicinal use. Previous studies have suggested that certain structural features, such as the presence of aromatic substituents at the C-3 position of the coumarin core connected through an amide linkage, may enhance anti-inflammatory activity [23]. Biosynthesis of these compounds proceeds through the shikimate-derived phenylpropanoid route. Key enzymes such as phenylalanine ammonia-lyase (PAL), cinnamate-4-hydroxylase (C4H), and coumarin synthases are involved in crucial reactions, including hydroxylation, cyclization, and methylation, that diversify the skeletons [24]. The pathway begins with phenylalanine, converted by PAL into cinnamic acid, which is then transformed through a series of steps into coumarins and related phenolics. Most of these compounds accumulate in the leaves of Calophyllum [21].
The objective of this review is to critically evaluate the current knowledge on coumarin compounds derived from the genus Calophyllum and their potential as anti-HIV agents. This study aims to compile and analyse reported phytochemical data, experimental bioassays, and molecular docking investigations related to Calophyllum coumarins and their interactions with key HIV targets such as reverse transcriptase (RT), and integrase (IN). By integrating findings from both experimental and computational studies, this review highlights important structural features that may influence binding interactions and biological activity. Furthermore, the review highlights existing research gaps and proposes future directions for the development of Calophyllum-derived coumarins as promising candidates for anti-HIV drug discovery and design.
Materials and methods
Search strategy
A systematic literature search was conducted to identify relevant studies on Calophyllum coumarins and their biological activities. The databases PubMed, Scopus, Web of Science, and Google Scholar were searched using Boolean operators combining the keywords (“Calophyllum” OR “Calophyllum coumarins”) AND (“anti-HIV” OR “antiviral”) AND (“molecular docking” OR “reverse transcriptase” OR “integrase” OR “1FK9” OR “3LPT”). The search was limited to peer-reviewed articles published in English between June 1968 and October 2025. Titles and abstracts were screened to assess their relevance to the topic, and the reference lists of selected articles were manually examined to identify additional relevant publications that may not have been captured during the initial database search.
Preparation of ligands
A total of 60 coumarins that have been isolated from the Malaysian genus Calophyllum were retrieved from the PubChem database (https://pubchem.ncbi.nlm.nih.gov/), and ChemDraw Ultra 12.0 was used to hand sketch the structures of compounds that were not found in PubChem; all these ligands were drawn, saved as SDF (Structure Data File) format and transformed into 3-dimensional structures, and geometry-optimized using Chem3D 23.1.1 (64-bit) before docking. The reference compounds Efavirenz (PubChem CID: 64139) and Raltegravir (PubChem CID: 54671008) were retrieved from the PubChem database in SDF format and then converted to PDB files using Open Babel integrated in PyRx 0.9.8 software. The openbabel tool embedded in PyRx was used to perform energy minimization for each ligand separately using the default parameters of steepest descent steps 100 with step size 0.02 (Å) and conjugate gradient steps 100 with step size 0.02 (Å), whereas the update interval was fixed at 10 [53]. Before or during molecular docking, energy minimization is an important step to make the structure less rigid and get rid of any steric interference for more accurate results [54].
Preparation of the target protein
The crystal structures of HIV-1 RT and IN were sourced from the Protein Data Bank with PDB IDs: 1F9K and 3LPT, respectively. Before docking, water molecules, other atoms, and ligand cocrystallized with the protein were removed with the Biovia Discovery Studio 2021 Client. The protein structure was minimized by using the conjugate gradient algorithm and the AMBER force field with UCSF Chimera 1.10.1 [55].
The PyRx virtual screening tool and the AutoDock Vina Wizard 4.2 were used for molecular docking. The grid box was adjusted to cover the binding sites, with the center size set to X: 23.5181, Y: 22.1833, Z: 21.1452 for 1FK9 and X: Y: Z: 15.4789 for 3LPT. This made sure that the ligand could fit into the pocket where it would bind. An exhaustive number of 8 was employed to make the docking results more accurate. The docked compounds were then evaluated based on their lowest binding energy (kJ/mol). The binding energy (ΔG) in kJ/mol of isolated ligands and standard drugs was determined by duplicating the docking experiments. The 2D and 3D depictions of the docking complexes were generated using Discovery Studio 2021 [54].
Drug-likeness and ADMET prediction
The drug-likeness properties of the selected compounds were evaluated using Lipinski’s rule of five through the SwissADME web server (www.swissadme.ch) [56,57]. Their pharmacokinetic properties, including absorption, distribution, metabolism, excretion, and toxicity (ADMET), were further assessed using the pkCSM platform (https://biosig.lab.uq.edu.au/pkcsm/) [58].
Results and discussion
Coumarins from Calophyllum species
The Calophyllum genus is known to contain various coumarin derivatives from its plant components, including stem bark, leaves, latex, and even fruits. From the species of this genus, many of these compounds have already been isolated, proving the inherent chemical diversity of Calophyllum. One of the most common coumarin compounds in C. recurvatum, C. andersonii, and C. inophyllum is Soulattrolide (1), which is found in stem bark and latex, making it more abundant in different species and parts of the plant. In addition, Inophyllum A (2), Inophyllum B (3), and Inophyllum P (5), which have been isolated in the leaves and fruit kernels of C. inophyllum, are the other most significant coumarin types. This suggests that some species of Calophyllum not only have multiple coumarins but also readily distribute them in various parts of the plant. Such distribution may be associated with the plant’s ecological adaptations or as a means of defense mechanism as coumarins have a vital role in protecting the plant against herbivore and pathogen invasion [59].
Calanolide A (6) and Calanolide B (7) are dominant in Calophyllum species. Such compounds were obtained from the latex, twigs, fruits, and stem bark of C. lanigerum and C. teysmannii. The fact that these calanolide types are frequently found in plant parts and in various studies highlights their significance for the genus. A pattern of chemical conservation among the many Calophyllum species is highlighted by this recurrent isolation, indicating that specific biosynthetic pathways are continuously active in several of the genus’s members [28].
In the leaves of C. lanigerum, other coumarins such as Cordatolide A (14) and Cordatolide B (15) have been identified, whereas the fruits and twigs of the same species also produced 12-Methoxycalanolide A (10) and 12-Methoxycalanolide B (11). Such intraspecific variation regarding the sources and types of coumarins suggests that species of Calophyllum have biosynthetic pathways that can synthesize diverse forms of coumarins in different tissues and organs of the plant. Table 1 provides a systematic summary of the coumarins extracted from Calophyllum species which illustrates a clear variety in both the types of compounds and parts of the plant. The widespread occurrence of some coumarins in many species and tissues indicates the great chemical potential of the genus. These results demonstrate that Calophyllum deserves intense attention and exploration for future research.
Table 1 Coumarins isolated from several Calophyllum species.
Compounds |
Species |
Part |
References |
Soulattrolide (1) |
C. recurvatum |
Stem bark |
[25] |
C. inophyllum |
Leaves |
[26] |
|
C. teysmannii |
Stem bark |
[27] |
|
C. teysmannii |
Latex |
[28] |
|
C. teysmannii |
Latex |
[29] |
|
Inophyllum A (2) |
C. inophyllum |
Leaves |
[26] |
C. inophyllum |
Fruit kernel |
[30] |
|
Inophyllum B (3) |
C. inophyllum |
Leaves |
[26] |
Inophyllum D (4) |
C. symingtonianum |
Bark & leaves |
[31] |
C. teysmannii |
Leaves & twigs |
[32] |
|
C. inophyllum |
Leaves |
[26] |
|
Inophyllum P (5) |
C. inophyllum |
Leaves |
[26] |
Calanolide A (6) |
C. lanigerum |
Fruit & twigs |
[7] |
C. lanigerum |
Stem bark |
[32] |
|
C. teysmannii |
Latex |
[29] |
|
Calanolide B (7)
|
C. lanigerum |
Fruit & twigs |
[7] |
C. teysmannii |
Latex |
[28] |
|
C. costatum |
Stem bark |
[33] |
|
C. brasiliense |
Leaves |
[34] |
|
C. teysmannii |
Stem bark |
[35] |
|
C. teysmannii |
Latex |
[29] |
|
Calanolide C (8) |
C. lanigerum |
Stem bark |
[32] |
12-Acetoxycalanolide A (9) |
C. lanigerum |
Fruit & twigs |
[7] |
12-Methoxycalanolide A (10) |
C. lanigerum |
Fruit & twigs |
[7] |
12-Methoxycalanolide B (11) |
C. lanigerum |
Fruit & twigs |
[7] |
Costatolide (12) |
C. teysmannii |
Stem bark |
[35] |
Calanolide F (13) |
C. teysmannii |
Leaves and twigs |
[32] |
Cordatolide A (14) |
C. lanigerum |
Leaves |
[32] |
Cordatolide B (15) |
C. lanigerum |
Leaves |
[32] |
Calanone (16) |
C. recurvatum |
Stem bark |
[25] |
C. lanigerum |
Stem bark |
[36] |
|
C. symingtonianum |
Bark & Leaves |
[31] |
|
C. teysmannii |
Stem bark |
[27] |
|
C. biflorum |
Latex |
[28] |
|
C. teysmannii |
Latex |
[28] |
|
C. teysmannii |
Latex |
[29] |
|
Calophyllolide (17) |
C. inophyllum |
Leaves |
[26] |
C. inophyllum |
Fruit kernel |
[30] |
|
Inophyllum C (18) |
C. inophyllum |
Leaves |
[26] |
C. teysmannii |
Stem bark |
[35] |
|
C. inophyllum |
Fruit kernel |
[30] |
|
C. inophyllum |
Leaves |
[37] |
|
C. inophyllum |
Seeds |
[38] |
|
C. apetalum |
Leaves |
[39] |
|
Inophyllum E (19) |
C. inophyllum |
Leaves |
[26] |
C. teysmannii |
Stem bark |
[35] |
|
C. inophyllum |
Fruit kernel |
[30] |
|
Soulattrolone (20) |
C. teysmannii |
Latex |
[28] |
Mammea A/BA cyclo F (21) |
C. dispar |
Stem bark |
[40] |
Mammea A/BB cyclo F (22) |
C. dispar |
Stem bark |
[40] |
Mammea A/BC cyclo F (23) |
C. dispar |
Fruit |
[40] |
Mammea A/AA cyclo F (24) |
C. dispar |
Stem bark |
[41] |
Mammea A/AB cyclo F (25) |
C. dispar |
Stem bark |
[41] |
Mammea A/AC cyclo F (26) |
C. dispar |
Fruit |
[41] |
Mammea A/AA methoxycyclo F (27) |
C. dispar |
Stem bark |
[41] |
Mammea A/AB dioxalanocyclo F (28) |
C. dispar |
Stem bark |
[41] |
Mammea A/AB cyclo E (29) |
C. dispar |
Stem bark |
[41] |
Calanolide E2 (30) |
C. depressinervosum |
Stem bark |
[42] |
Calopolyanolide A (31) |
C. depressinervosum |
Stem bark |
[42] |
C. sclerophyllum |
Stem bark |
[43] |
|
Calanolide D (32) |
C. lanigerum |
Fruit & twigs |
[7] |
Isodispar B (33) |
C. dispar |
Fruit |
[41] |
C. sclerophyllum |
Stem bark |
[44] |
|
C. sclerophyllum |
Stem bark |
[45] |
|
Calophyllic Acid (34) |
C. inophyllum |
Leaves |
[26] |
Isocalophyllic Acid (35) |
C. inophyllum |
Leaves |
[26] |
Teysmanone A (36) |
C. recurvatum |
Stem bark |
[25] |
C. teysmannii |
Stem bark |
[35] |
|
Cordatolide E (37) |
C. lanigerum |
Stem bark |
[32] |
Isocalanone (38) |
C. andersonii |
Stem bark |
[25] |
C. ferrugineum |
Stem bark |
[46] |
|
C. teysmannii |
Stem bark |
[27] |
|
Disparfuran B (39) |
C. dispar |
Stem bark |
[41] |
Disparacetylfuran A (40) |
C. dispar |
Stem bark |
[41] |
Inophyllum G-1 (41) |
C. inophyllum |
Leaves |
[26] |
Inophyllum G-2 (42) |
C. inophyllum |
Leaves |
[26] |
Inophyllum A 4-bromobenzoate (43) |
C. inophyllum |
Leaves |
[26] |
Inophyllum P acetate (44) |
C. inophyllum |
Leaves |
[26] |
Inophyllum B acetate (45) |
C. inophyllum |
Leaves |
[26] |
11,12-Anhydroinophyllum P (46) |
C. inophyllum |
Leaves |
[26] |
Isodisparfuran A (47) |
C. dispar |
Fruit |
[40] |
Mucigerin (48) |
C. mucigerum |
Stem bark |
[47] |
Incrassamarin A (49) |
C. incrassatum |
Stem bark |
[48] |
Incrassamarin B (50) |
C. incrassatum |
Stem bark |
[48] |
Incrassamarin C (51) |
C. incrassatum |
Stem bark |
[48] |
Incrassamarin D (52) |
C. incrassatum |
Stem bark |
[48] |
Soulamarin (53) |
C. soulattri |
Stem bark |
[49] |
Teysmanone B (54) |
C. teysmannii |
Stem bark |
[35] |
Wallimarin T (55) |
C. wallichianum |
Stem bark |
[50] |
Benjaminin (56) |
C. benjaminum |
Stem bark |
[51] |
Gracilenin A (57) |
C. gracilentum |
Stem bark |
[52] |
Gracilenin B (58) |
C. gracilentum |
Stem bark |
[52] |
Gracilenin C (59) |
C. gracilentum |
Stem bark |
[52] |
Hoseimarin (60) |
C. hosei |
Bark |
Biological activities of coumarins
Calanolide C (8), Calanolide F (13), along with Calanolide E2 (30), represent another group of Calophyllum coumarins with notable anti-HIV activity. Although their binding affinities are somewhat lower than those of Calanolide A (6) and B (7), these compounds still demonstrate meaningful inhibition of HIV-1 reverse transcriptase. Their inclusion in studies broadens the range of Calophyllum compounds that may be optimized and developed into effective antiviral agents. Moreover, their diverse structural characteristics offer valuable data for designing next-generation inhibitors [32-34]. The biological activities described in Table 2 include significant cytotoxic, anti-inflammatory, and antibacterial properties. Calophyllolide (17) and Inophyllum E (19), for instance, exhibit significant toxicity to human epidermoid cancer cells [60]. These results suggest that coumarins derived from Calophyllum may possess applications beyond antiviral therapy. The documented actions create avenues for further investigation into their roles as anticancer and antibacterial agents, along with their therapeutic efficacy in inflammatory conditions. In summary, the bioactivity highlights the exceptional potential of Calophyllum coumarins, particularly in the context of HIV treatment. Some of the compounds that have been proven to inhibit HIV-1 RT significantly are Soulattrolide (1), Calanolide A (6), and Calanolide B (7). These drugs have low IC50 and EC50 values [33,34,61,62]. Inophyllum derivatives also have a high potency and distinctive structure, further substantiating that Calophyllum is a valuable source of bioactive compounds. Studies to date cover a range of coumarins isolated from different Calophyllum species, underscoring how important this genus has become in natural-product chemistry and the ongoing search for new therapeutic agents.
Table 2 Biological activities of several Calophyllum phytochemicals.
Compounds |
Species |
Description |
Soulattrolide (1)
|
C. brasiliense |
Anti-HIV: Showed a potent inhibition against HIV-1 reverse transcriptase in human lymphatic MT2 cell with percent inhibition 77.7 ± 1.6% [34]. |
C. brasiliense |
Anti-HIV: Showed a potent inhibition against HIV-1 reverse transcriptase with percent inhibition 77.7 ± 1.6% [34]. |
|
C. recurvatum
|
Cytotoxic: Showed a notably activity against HepG2 cell lines with IC50 value 34.53 μg/mL while for HeLa Chang liver, and HL-7702 cell lines there are no activity found with their IC50 values > 100 µg/mL [25]. |
|
C. teysmannii |
Anti-HIV: Showed a potent inhibitor activity of HIV-1 reverse transcriptase with an IC50 of 0.34 µM [61]. |
|
Inophyllum B (3) |
C. inophyllum |
Anti-HIV: Showed the most potent activity with IC50 value 1.4 µM [37]. |
C. inophyllum |
Anti-HIV: Strongly inhibited HIV-1 reverse transcriptase cell culture with IC50 values of 1.4 µM [26]. |
|
Inophyllum D (4) |
C. symingtonianum |
Enzyme inhibitor (α-glucoside, AG): Showed potent activity inα-glucosidase (maltase) inhibitory with IC50 value 35.7 ± 1.1 µM [31]. |
C. inophyllum |
Anti-HIV: Displayed an active activity in inhibit HIV-1 reverse transcriptase cell culture with IC50 values of 11.0 µM [26]. |
|
Inophyllum P (5) |
C. inophyllum |
Anti-HIV: Highly potent in inhibit HIV-1 reverse transcriptase cell culture with IC50 values of 1.6 µM [26]. |
Calanolide A (6) |
C. lanigerum |
Anti-HIV: Display highly protective against HIV-1 replication and cytopathicity with EC50 values of 0.1 µM [62]. |
C. brasiliense |
Anti-HIV: Showed a potent inhibition against HIV-1 reverse transcriptase in human lymphatic MT2 cell with percent inhibition 81.5 ± 0.9% [34]. |
|
C. brasiliense |
Anti-tumor: Showed moderate inhibition of activity against vitro assay of TPA-induced EBV-EA activation in Raji cells with IC50 290 µg/mL [63]. |
|
C. brasiliense |
Antivirus: Completely protective against HIV-1 replication and cyctopathicity with EC50 values of 0.1 mM [33]. |
|
C. cordato-oblongum |
Anti-HIV: Showed highly remarkably activity inhibit HIV-1 reverse transcriptase assay compared to control with IC50 values of 0.32 µM [64]. |
|
C. lanigerum |
Anti-HIV: Showed highly potent to inhibit the replication of HIV-1 and protect cells from the cytopathic effects of the virus with EC50 values 0.1 µM [7]. |
|
Calanolide B (7) |
C. lanigerum |
Anti-HIV: Display highly protective against HIV-1 replication and cytopathicity with EC50 values of 0.4 µM [62]. |
C. brasiliense |
Anti-HIV: Showed a potent inhibition against HIV-1 reverse transcriptase in human lymphatic MT2 cell with percent inhibition 76.2 ± 2.2% [34]. |
|
C. brasiliense |
Anti-HIV: Showed a potent inhibition against HIV-1 reverse transcriptase with percent inhibition 76.2 ± 2.2% [34]. |
|
C. brasiliense |
Antivirus: Showed protective activity against HIV-1 replication and cyctopathicity with EC50 values of 0.4 mM [33]. |
|
C. lanigerum |
Anti-HIV: Showed highly potent to inhibit the replication of HIV-1 and protect cells from the cytopathic effects of the virus with EC50 values 0.4 µM [7]. |
|
C. brasiliense |
Anti-HIV: Showed a moderate inhibition against HIV-1 reverse transcriptase in human lymphatic MT2 cell with percent inhibition 50.7 ± 2.0% [34]. |
|
C. brasiliense |
Anti-tumor: Showed moderate inhibition of activity against vitro assay of TPA-induced EBV-EA activation in Raji cells with IC50 351 µg/mL [63]. |
|
Calanolide F (13) |
C. teysmannii |
Anti-HIV: Showed potent activity in the NCI’s primary assay with IC50 12.7 μM [32]. |
Cordatolide A (14) |
C. cordato-oblongum |
Anti-HIV: Displayed highly potent to inhibit HIV-1 reverse transcriptase assay with IC50 values of 12.3 µM [64]. |
Cordatolide B (15) |
C. cordato-oblongum |
Anti-HIV: Displayed highly potent to inhibit HIV-1 reverse transcriptase assay with IC50 values of 19.0 µM [64]. |
C. lanigerum |
Anti-HIV: Showed potent activity in the NCI’s primary assay with IC50 14.0 μM [32]. |
|
Calanone (16) |
C. recurvatum
|
Cytotoxic: Showed a moderate activity against HepG2 cell lines with IC50 value 75.25 μg/mL while for HeLa Chang liver, and HL-7702 cell lines there are no activity found with their IC50 values > 100 µg/mL [25]. |
C. brasiliense |
Anti-tumor: Showed moderate inhibition of activity against vitro assay of TPA-induced EBV-EA activation in Raji cells with IC50 347 µg/mL [63]. |
|
Calophyllolide (17) |
C. inophyllum |
Anti-inflammatory: Have a potent therapeutic for cutaneous wound healing treatment against HaCaT and RAW264.7 cell viability [65]. |
C. brasiliense |
Anti-tumor: Strongly induce apoptosis in HL-60 leukemia cells by activating a specific pathway to mitochondrial dysfunction with IC50 8.7 μM [66]. |
|
|
C. inophyllum |
Cytotoxicity: Display potent activity against human epidermoid carcinoma of the nasopharynx cell (KB) with IC50 value 3.5 μM [60]. Antimicrobial: Displayed potent against Staphylococcus aureus (ATCC6538) with IC50 value 16.0 μM [60]. |
C. inophyllum |
Anti-inflammatory: Displayed an effective in reducing the increased capillary permeability induced that involved in this process against, histamine (HA), 5-hydroxytryptamine (5-HT) and bradykinin (BK) with PD50 value 144.1, 250.0 and 133.5 mg/kg, respectively [67]. |
|
Inophyllum C (18) |
C. inophyllum |
Antimicrobial: Displayed potent against Staphylococcus aureus (ATCC6538) with IC50 value 10.0 μM [60]. |
C. inophyllum |
Anti-HIV: Showed significant in inhibit HIV-1 reverse transcriptase cell culture with IC50 values of 10.0 µM [26]. |
|
Inophyllum E (19) |
C. inophyllum |
Cytotoxicity: Display potent activity against human epidermoid carcinoma of the nasopharynx cell (KB) with IC50 value 36.1 μM [60]. Antimicrobial: Displayed potent against Staphylococcus aureus (ATCC6538) with IC50 value 13.0 μM [60]. |
Calanolide E2 (30) |
C. lanigerum |
Anti-HIV: Displayed a fit patterns activity in the NCI’s primary assay with IC50 2.5 μM [32]. |
Isodispar B (33) |
C. sclerophyllum |
Cytotoxicity: Exerted the highest activity against nasopharyngeal cancer cell lines (SUNE1, TW01, CNE1, HK1) with IC50 values ranging from 3.8 to 11.5 μM [44]. |
C. dispar |
Cytotoxic: Displayed significant activities against human KB cancer cell lines with ED50 value 8 µg/mL [40]. |
|
Calophyllic acid (34) |
C. inophyllum |
Antidiabetic: Highly can inhibits palmitate-induced, reactive oxygen species-associated MAPK kinase activation and restored insulin sensitivity through regulating IRS-1 function for insulin resistance and type 2 diabetes with a significant response observed at 5 µM concentration of this compound [68]. |
C. inophyllum |
Antidiabetic: Highly can activate glucose uptake through PI-3-K-dependent and extracellular signal-regulated kinase 1 and 2 (EKR 1/2) in skeletal muscle cells of type 2 diabetes by enhancing translocation of GLUT4 to plasma membrane with the phosphorylation of AKT levels from 1.0 - 2.0 AU [69]. |
|
Isocalophyllic acid (35) |
C. inophyllum |
Antidiabetic: Highly can inhibits palmitate-induced, reactive oxygen species-associated MAPK kinase activation and restored insulin sensitivity through regulating IRS-1 function for insulin resistance and type 2 diabetes with a significant response observed at 2.55 µM concentration of this compound [68]. |
C. inophyllum |
Antidiabetic: Highly can activate glucose uptake through PI-3-K-dependent and extracellular signal-regulated kinase 1 and 2 (EKR 1/2) in skeletal muscle cells of type 2 diabetes by enhancing translocation of GLUT4 to plasma membrane with the phosphorylation of AKT levels from 1.0 - 2.0 AU [69]. |
|
Teysmanone A (36) |
C. recurvatum
|
Cytotoxic: Showed a portrayed appreciable activity against HepG2 cell lines with IC50 value 42.57 μg/mL while for HeLa Chang liver, and HL-7702 cell lines there are no activity found with their IC50 values > 100 µg/mL [25]. |
Cordatolide E (37) |
C. lanigerum |
Anti-HIV: Showed potent activity in the NCI’s primary assay with IC50 value 14.0 μM [32]. |
Isocalanone (38) |
C. andersonii |
Cytotoxic: Showed a moderate activity against HepG2 cell lines with IC50 value 75.68 μg/mL while for HeLa Chang liver, and HL-7702 cell lines there are no activity found with their IC50 values > 100 µg/mL [25]. |
Incrassamarin A (49) |
C. incrassatum |
Antidiabetic: Shows strong inhibition against α-Glucosidase enzyme with IC₅₀ value 53.48% [48]. |
Incrassamarin C (51) |
C. incrassatum |
Cytotoxicity: Display moderate activity against cell cancer line MCF-7 and A-549 with IC₅₀ value 73.7 µg/mL [48]. |
Molecular docking
In this study, molecular docking was employed to explore whether coumarins from Calophyllum could play a part in suppressing HIV replication. The approach allowed a closer examination of the key protein-ligand contacts that define how these molecules interact with their targets. From such analyses, researchers can infer the chemical traits that appear to make certain coumarins more reactive or selective than others. During the viral life cycle, the enzymes RT and IN act in sequence, where RT converts the viral RNA into DNA, while IN inserts that DNA into the host genome. Interfering with either step effectively halts replication. The docking evaluation of 60, therefore, provides a useful starting point for designing anti-HIV candidates. Table 3 summarizes the binding affinities of the tested coumarin derivatives, and Figures 2 and 3 illustrate the corresponding 2D and 3D ligand-protein complexes.
Table 3 Binding energies (in kJ/mol) of coumarin derivatives against 1F9K and 3LPT.
Compounds / Ligands |
Binding Affinity (1F9K) |
Binding Affinity (3LPT) |
Efavirenz |
−27.20 |
- |
Raltegravir |
- |
−26.36 |
Soulattrolide (1) |
−26.78 |
−25.52 |
Inophyllum A (2) |
−29.29 |
−26.36 |
Inophyllum B (3) |
−28.45 |
−25.52 |
Inophyllum D (4) |
−27.20 |
−25.94 |
Inophyllum P (5) |
−26.78 |
−27.20 |
Calanolide A (6) |
−27.61 |
−24.69 |
Calanolide B (7) |
−27.61 |
−25.10 |
Calanolide C (8) |
−26.36 |
−26.78 |
12-Acetoxycalanolide A (9) |
−28.03 |
−23.43 |
12−Methoxycalanolide A (10) |
−24.69 |
−22.18 |
12−Methoxycalanolide B (11) |
−25.94 |
−22.59 |
Costatolide (12) |
−26.36 |
−23.43 |
Calanolide F (13) |
−26.36 |
−23.43 |
Cordatolide A (14) |
−28.87 |
−23.85 |
Cordatolide B (15) |
−27.20 |
−25.94 |
Calanone (16) |
−29.29 |
−25.94 |
Calophyllolide (17) |
−26.78 |
−24.69 |
Inophyllum C (18) |
−31.38 |
−26.36 |
Inophyllum E (19) |
−31.38 |
−28.87 |
Soulattrolone (20) |
−30.96 |
−29.29 |
Mammea A/BA cyclo F (21) |
−26.78 |
−24.69 |
Mammea A/BB cyclo F (22) |
−27.61 |
−23.85 |
Mammea A/BC cyclo F (23) |
−28.45 |
−22.59 |
Mammea A/AA cyclo F (24) |
−26.36 |
−23.85 |
Mammea A/AB cyclo F (25) |
−27.20 |
−24.27 |
Mammea A/AC cyclo F (26) |
−28.03 |
−23.85 |
Mammea A/AA methoxycyclo F (27) |
−25.94 |
−24.27 |
Mammea A/AB dioxalanocyclo F (28) |
−25.94 |
−28.87 |
Mammea A/AB cyclo E (29) |
−27.20 |
−25.52 |
Calanolide E2 (30) |
−24.27 |
−24.69 |
Calopolyanolide A (31) |
−26.36 |
−24.27 |
Calanolide D (32) |
−25.94 |
−25.52 |
Isodispar B (33) |
−28.03 |
−23.01 |
Calophyllic Acid (34) |
−26.36 |
−23.85 |
Isocalophyllic Acid (35) |
−25.94 |
−23.85 |
Teysmanone A (36) |
−33.05 |
−27.61 |
Cordatolide E (37) |
−27.20 |
−24.27 |
Isocalanone (38) |
−29.29 |
−28.45 |
Disparfuran B (39) |
−29.29 |
−26.36 |
Disparacetylfuran A (40) |
−28.03 |
−25.94 |
Inophyllum G−1 (41) |
−27.61 |
−26.36 |
Inophyllum G−2 (42) |
−31.38 |
−29.29 |
Inophyllum A 4−bromobenzoate (43) |
−29.71 |
−29.71 |
Inophyllum P acetate (44) |
−27.20 |
−25.94 |
Inophyllum B acetate (45) |
−29.29 |
−26.36 |
11,12−Anhydroinophyllum P (46) |
−28.45 |
−27.20 |
Isodisparfuran A (47) |
−28.45 |
−23.43 |
Mucigerin (48) |
−25.52 |
−22.18 |
Incrassamarin A (49) |
−28.03 |
−25.94 |
Incrassamarin B (50) |
−26.36 |
−21.76 |
Incrassamarin C (51) |
−26.36 |
−25.10 |
Incrassamarin D (52) |
−28.03 |
−23.01 |
Soulamarin (53) |
−28.03 |
−23.01 |
Teysmanone B (54) |
−29.71 |
−23.85 |
Wallimarin T (55) |
−23.85 |
−20.08 |
Benjaminin (56) |
−27.61 |
−20.08 |
Gracilenin A (57) |
−25.94 |
−19.66 |
Gracilenin B (58) |
−25.94 |
−20.08 |
Gracilenin C (59) |
−25.10 |
−22.18 |
Hoseimarin (60) |
−26.36 |
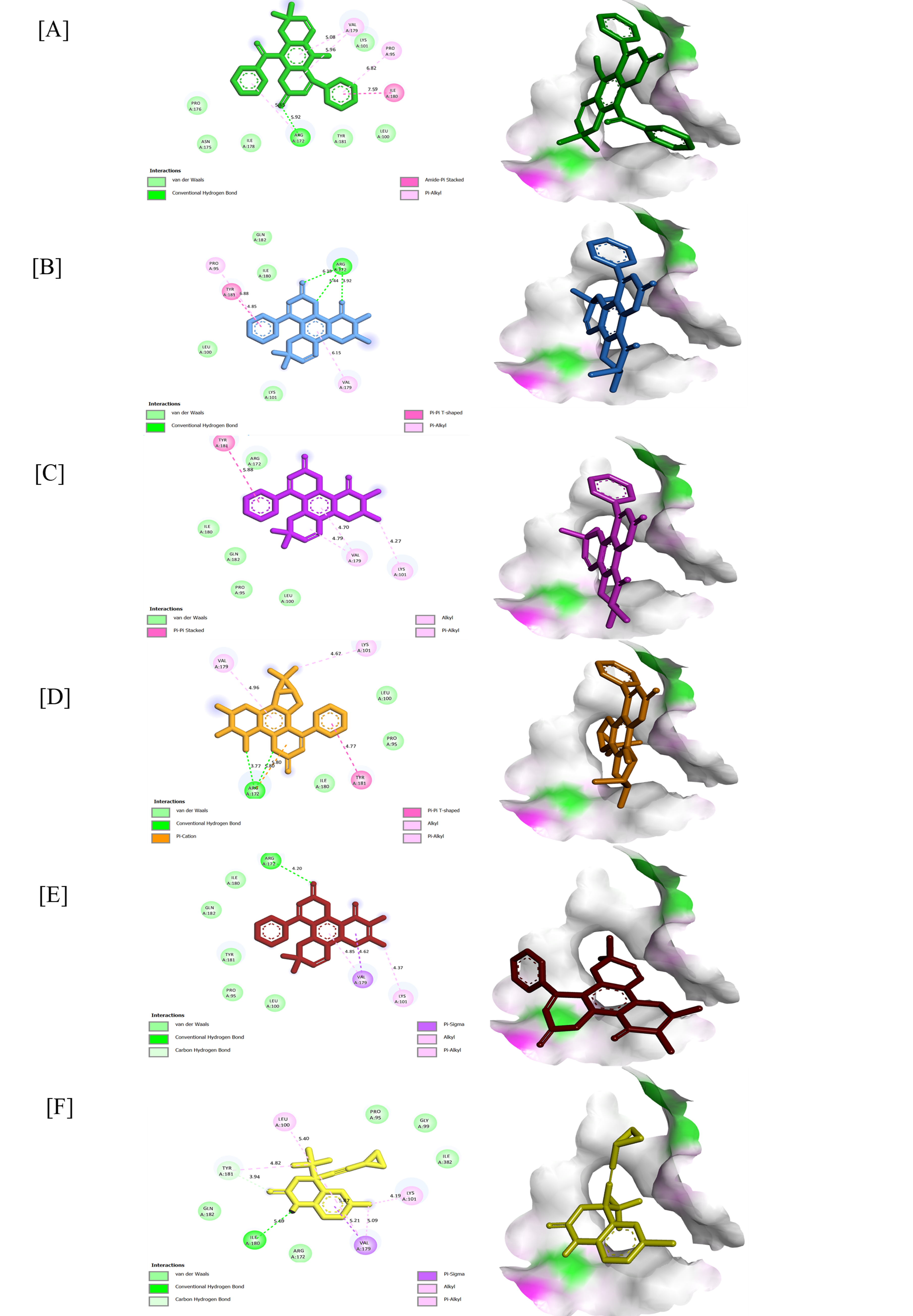
Figure 2 2D and 3D conformation view of (36); (A), (19) (B), (18) (C), (41) (D) (20) (E) and Efavirenz (F) against 1F9K.
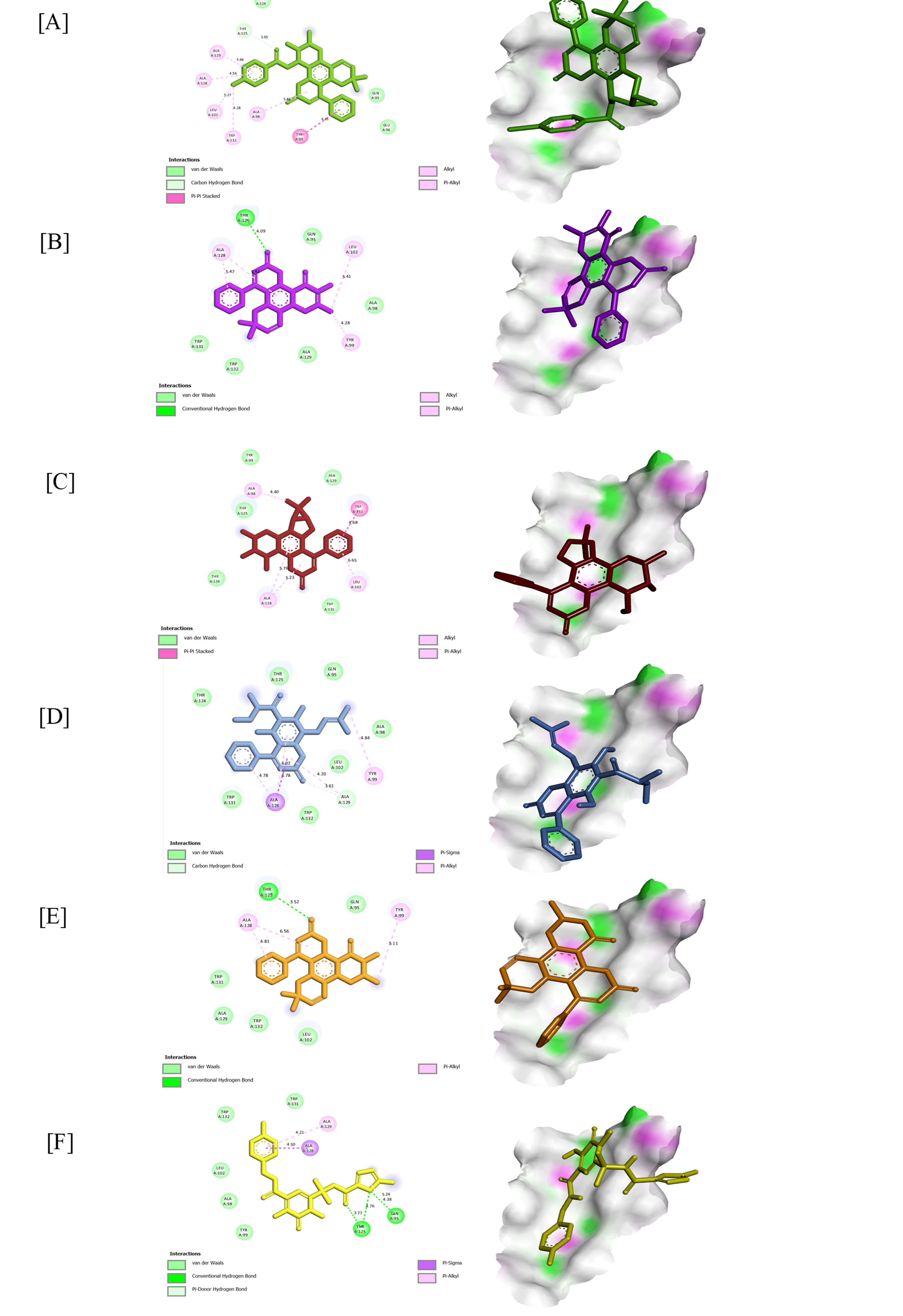
Figure 3 2D and 3D conformation view of (43) (A), (20) (B), (42) (C), (28) (D), (19) (E), and Raltegravir (F) against 3LPT.
HIV-1 RT target
Docking with RT demonstrated that Efavirenz (−27.20 kJ/mol) interacted mainly through π-π and hydrophobic contacts with residues Tyr181, Lys101, Val179, and Ile180, forming a representative non-nucleoside inhibitor (NNRTI) binding profile. In comparison, Calophyllum coumarins exhibited comparable with remarkable affinities.
Teysmanone A (36) (−33.05 kJ/mol) established conventional hydrogen bonds between its carbonyl group and Arg172 (5.65 Å) and van der Waals interactions with Tyr181 and Ile180, complemented by π-alkyl contacts with Val179 (5.08 Å) and Lys101 (5.96 Å). Its extended conjugation and prenylated ring contribute to amide π-stacking and van der Waals stabilization within the NNRTI pocket. Such contacts mediate the relevant conformational flexibility of the domain polymerase, allowing for the proper positioning of incoming nucleotides.
Inophyllum E (19) (−31.38 kJ/mol) donated hydrogen bonds to Arg172 (3.92 - 6.38 Å) through conventional hydrogen interaction. Tyr181 (4.85 Å) and formed π-π T-shaped stacking with the aromatic system, while Val179 (6.15 Å) anchors the coumarin core within the π-alkyl linking.
Inophyllum C (18) and Inophyllum G-1 (41) (−31.38 kJ/mol) engaged in π-π stacking with Tyr181 and hydrophobic contacts with Val179 and Lys101 (4.27 - 4.96 Å). These interactions are known to allosterically restrict the conformational flexibility of the thumb subdomain of HIV-1 RT, a region responsible for positioning the nucleic acid duplex during polymerization.
Soulattrolone (20) (−30.96 kJ/mol) displayed a bonding within the carbonyl of ligands and Arg172 (4.20 Å) through conventional hydrogen linkage and other interactions as well. These ligands collectively occupied the NNRTI allosteric pocket by exhibiting significant RT inhibitory activity through a combination of polar and hydrophobic interactions.
HIV-1 IN target
For HIV-1 IN, the control ligand raltegravir (−26.36 kJ/mol) interacted through conventional hydrogen bonding with Thr125 (3.77 - 4.76 Å) and Gln95 (4.38 Å), alongside π-alkyl and π-sigma, which interacted with Ala129 (4.21 Å) and Ala128 (4.50 Å), respectively. Typically, it is inhibitors that chelate or block the DDE catalytic triad that is responsible for DNA-strand transfer. In contrast, several coumarin analogues displayed stronger affinities.
Inophyllum A 4-bromobenzoate (43) (−29.71 kJ/mol) formed a carbon-hydrogen bond between carbonyl oxygen and Thr125 (3.65 Å), while π-π stacking with Tyr99 (4.61 Å). Several alkyl and π-alkyl interactions were observed within the range 4.28 - 5.66 Å. While the bromophenyl substituent enhanced π-electron delocalization, strengthening hydrophobic and van der Waals interactions that could hinder access to the viral DNA-binding groove.
Soulattrolone (20) (−29.29 kJ/mol) formed a conventional hydrogen bond with Thr125 (4.09 Å) and π-alkyl contacts with Tyr99 and Leu102 (4.28 - 5.41 Å), occupying the catalytic loop that initiates strand transfer. At the same time, Ala128 formed an alkyl interaction within the active site cleft of the aromatic system of the coumarin core.
Inophyllum G-2 (42) (−29.29 kJ/mol) showed π-π stacking with Trp132 and π-alkyl contacts with Ala98, including alkyl interaction with Leu102 and Ala128. These interactions suggest that its prenylated side chain fits into the hydrophobic DNA-binding channel, potentially disrupting the coordination geometry of the Mg²⁺ cofactors required for catalytic activity.
Mammea A/AB dioxalanocyclo F (28) (−28.87 kJ/mol) displayed a carbon-hydrogen bond with Ala129 (3.61 Å) and π-alkyl interactions with Tyr99, Ala129, and Ala128 (4.20 - 4.84 Å), indicating stabilization of the dioxolane ring system through this interaction.
Inophyllum E (19) (−28.87 kJ/mol) exhibited a conventional hydrogen bond with Thr125 and several π-alkyl interactions with Ala128 and Tyr99 together with van der Waals contacts, suggesting a close aromatic fit within the catalytic pocket of integrase.
In addition to this study, calanolide A (6), a known anti-HIV coumarin from C. lanigerum, exhibited moderate binding toward both HIV-1 RT (−6.6) and IN (−5.9) compared to the newly evaluated analogues. Its lower affinity is likely due to a limited hydrophobic surface and a lack of prenyl or brominated substituents that enhance π-stacking stabilization. Nevertheless, its conserved binding orientation supports its role as a reliable benchmark for validating the docking outcomes of Calophyllum coumarins. Overall, the docking analysis of Calophyllum-derived compounds shows significant affinity for both the HIV-1 reverse-transcriptase (RT) and integrase (IN) binding pockets, in several instances performing even better than the reference drugs Efavirenz and Raltegravir. Interaction maps reveal a mix of π–π stacking, π–alkyl, and hydrogen-bond contacts. Residues which include Tyr181, Lys101, and Arg172 in RT, as well as Thr125, Ala128, and Trp132 in IN, seem to be crucial for keeping these complexes stable. Molecules with larger conjugated aromatic systems or prenyl-type side groups tend to bind better.
Drug-likeness and ADMET prediction
The drug-likeness of the selected 12 phytocompounds, chosen based on their docking scores, was evaluated using Lipinski’s rule of five (RO5) to estimate their potential suitability as orally active drug candidates. The calculated physicochemical parameters, including molecular weight (MW), lipophilicity (LogP), number of rotatable bonds (NORTB), hydrogen bond acceptors (HBA), hydrogen bond donors (HBD), and Lipinski’s rule violations, are summarized in Table 4. Efavirenz and Raltegravir were included as reference drugs for comparison. Several of the investigated phytocompounds showed drug-likeness profiles comparable to those of these reference compounds.
Table 4 Predicted drug-likeness ability for some chosen compounds.
Compound |
MWa |
LogPb |
NORTBc |
HBAd |
HBDe |
Lipinski’s violationf |
Calanolide A (6) |
370.44 |
3.80 |
2 |
5 |
1 |
0 |
Inophyllum C (18) |
402.44 |
4.34 |
1 |
5 |
0 |
0 |
Inophyllum E (19) |
402.44 |
4.33 |
1 |
5 |
0 |
0 |
Soulattrolone (20) |
402.44 |
4.33 |
1 |
5 |
0 |
0 |
Mammea A/AB dioxalanocyclo F(28) |
436.45 |
3.97 |
4 |
7 |
1 |
0 |
Teysmanone A (36) |
424.44 |
4.57 |
3 |
5 |
1 |
0 |
Isocalanone (38) |
424.44 |
4.70 |
3 |
5 |
1 |
0 |
Inophyllum G-1 (41) |
404.46 |
4.08 |
1 |
5 |
1 |
0 |
Inophyllum G-2 (42) |
404.46 |
4.12 |
1 |
5 |
1 |
0 |
Inophyllum A 4-bromobenzoate (43) |
587.46 |
6.23 |
4 |
6 |
0 |
2 |
Efavirenz |
315.67 |
3.80 |
1 |
5 |
1 |
0 |
Raltegravir |
444.42 |
1.46 |
8 |
9 |
3 |
1 |
aMW: Molecular weight, bLogP: Partition coefficient, cNORTB: Rotatable bonds, dHBA: H-bond acceptors, eHBD: H-bond donors, fLipinski’s violation: 0 = good
Most of the studied phytocompounds, Calanolide A (6), Inophyllum C (18), Inophyllum E (19), Soulattrolone (20), Mammea A/AB dioxalanocyclo F (28), Teysmanone A (36), Isocalanone (38), Inophyllum G-1 (41), and Inophyllum G-2 (42), exhibited favorable drug-likeness characteristics and satisfied the RO5 criteria, including LogP ≤ 5, MW ≤ 500 Da, HBAs ≤ 10, and HBDs ≤ 5. However, Inophyllum A 4-bromobenzoate (43) showed 2 violations of Lipinski’s rule, mainly due to its higher MW and increased lipophilicity (LogP = 6.23). These features may limit its oral bioavailability. However, such deviations do not necessarily exclude its potential as a bioactive molecule, particularly if alternative formulation or delivery approaches are considered. Overall, the results suggest that most of the selected phytocompounds possess physicochemical properties consistent with drug-like molecules, supporting their further investigation in pharmacokinetic and biological studies.
The ADMET properties of the selected 12 phytocompounds were further evaluated to gain further insight into their predicted pharmacokinetic behavior and safety profiles. Parameters related to absorption, distribution, metabolism, excretion, and toxicity are summarized in Table 5. Efavirenz and Raltegravir were included as reference compounds to facilitate comparison. All phytocompounds exhibited relatively low predicted aqueous solubility, with water solubility values ranging from −3.01 to −5.35 log mol/L. Despite this, the compounds showed consistently high predicted human intestinal absorption, with most values exceeding 95%. This suggests that, although solubility may be limited, efficient oral absorption is still likely. Low aqueous solubility may therefore be addressed through appropriate formulation strategies to support oral bioavailability [58,70]. The detailed distribution indicated that most phytocompounds showed negative log BB values, indicating limited ability to cross the blood-brain barrier. This was supported by low predicted CNS permeability values, suggesting minimal central nervous system exposure. Note that, Efavirenz exhibited higher BBB permeability, consistent with its known pharmacological profile.
Table 5 Predicted ADMET characteristics for some selected compounds.
No |
Compounds |
Adsorption |
Distribution |
Metabolism |
Excretion |
Toxicity |
||||||
Water solubility |
Intestinal absorption (human) |
BBB permeability |
CNS permeability |
CYP |
Renal OCT2 substrate |
Total Clear anc |
AMES toxicity |
|||||
2D6 |
3A4 |
2D6 |
3A4 |
|||||||||
Substate |
Inhibitor |
|||||||||||
Numeric (log mol/L) |
Numeric (% Absorbed) |
Numeric (log BB) |
Numeric (log PS) |
Categorical (Yes/No) |
Categorical (Yes/No) |
Numeric (log ml/min/kg) |
Categorical (Yes/No) |
|||||
1 |
Calanolide A (6) |
−4.982 |
94.94 |
−0.322 |
−1.820 |
No |
Yes |
Yes |
Yes |
No |
0.506 |
No |
2 |
Inophyllum C (18) |
−4.952 |
100 |
−0.453 |
−1.606 |
No |
Yes |
Yes |
Yes |
No |
0.589 |
No |
3 |
Inophyllum E (19) |
−4.952 |
100 |
−0.453 |
−1.606 |
No |
Yes |
Yes |
Yes |
No |
0.589 |
No |
4 |
Soulattrolone (20) |
−4.952 |
100 |
−0.453 |
−1.606 |
No |
Yes |
Yes |
Yes |
No |
0.589 |
No |
5 |
Mammea A/AB dioxalanocyclo F (28) |
−5.352 |
99.946 |
−0.39 |
−2.966 |
No |
Yes |
Yes |
Yes |
No |
0.28 |
No |
6 |
Teysmanone A (36) |
−4.075 |
98.584 |
−0.35 |
−1.700 |
No |
Yes |
Yes |
Yes |
No |
0.633 |
No |
7 |
Isocalanone (38) |
−4.456 |
97.985 |
-0.27 |
−1.736 |
No |
Yes |
Yes |
No |
No |
0.645 |
No |
8 |
Inophyllum G-1 (41) |
−4.807 |
97.869 |
0.094 |
−1.710 |
No |
Yes |
Yes |
Yes |
No |
0.052 |
No |
9 |
Inophyllum G-2 (42) |
−4.807 |
97.869 |
0.094 |
−1.710 |
No |
Yes |
Yes |
Yes |
No |
0.052 |
No |
10 |
Inophyllum A 4-bromobenzoate (43) |
−4.668 |
95.669 |
−0.838 |
−1.353
|
No |
Yes |
Yes |
No |
No |
−0.115 |
No |
11 |
Efavirenz |
−5.017 |
88.476 |
0.397 |
−1.977 |
No |
Yes |
No |
No |
No |
−0.141 |
No |
12 |
Raltegravir |
−3.012 |
67.455 |
−1.377 |
−4.129 |
No |
No |
No |
No |
No |
0.225 |
|
Metabolic predictions suggested that many of the phytocompounds may interact with cytochrome P450 enzymes, particularly CYP3A4 and CYP2D6, either as substrates or inhibitors. These results indicate that hepatic metabolism is likely to play an important role in their biotransformation. Notably, compounds such as Isocalanone (38) and Inophyllum A 4-bromobenzoate (43) showed fewer predicted CYP interactions, which may be advantageous in terms of metabolic liability. For excretion, none of the phytocompounds were predicted to be substrates of the renal organic cation transporter OCT2, suggesting that transporter-mediated renal clearance is unlikely to be a major elimination pathway. Predicted total clearance values were generally moderate, indicating balanced elimination behavior. Notably, none of the phytocompounds investigated were predicted to be AMES toxic, suggesting a low risk of mutagenicity at this preliminary stage. This toxicity profile was comparable to that observed for the reference drugs. Overall, the ADMET predictions suggest that, although limited aqueous solubility may present a challenge, the selected phytocompounds generally exhibit favorable absorption, acceptable distribution characteristics, manageable metabolic profiles, and low predicted toxicity. These findings support their continued evaluation as potential drug candidates, particularly with consideration of formulation approaches to improve solubility.
Conclusions
In summary, studying how coumarins interact at a molecular level helps to explain why these compounds continue to draw attention as possible sources of new medicines. Early study on the genus revealed a striking range of coumarin frameworks, and many of them remain under investigation for potential therapeutic use. During the docking stage, selection of protein targets and adjustment of ligand configurations proved essential for achieving significant binding predictions and pinpointing which parts of each molecule appear to drive activity. The results from docking give a better understanding of how ligands and proteins interact and help to choose the best candidates. The 7 compounds with the highest results also have good pharmacokinetic and safety features, according to drug-likeness and ADMET tests. This shows how beneficial they could be as starting points for future improvements. These results generally assist in further discovering Calophyllum coumarins, as they could serve as templates for developing antiviral and other effective medications.
Acknowledgements
This research was supported by the Geran Penyelidikan Universiti (Kecemerlangan@UPSI) (Grant No. 2025-0012-103-01), funded by Universiti Pendidikan Sultan Idris (UPSI), Malaysia. The authors also acknowledge the Department of Chemistry, Faculty of Science and Mathematics, UPSI, for providing the necessary research facilities.
Declaration of generative AI in scientific writing
This manuscript used generative artificial intelligence (AI) tools, namely ChatGPT for grammar checks. All scientific content, analysis, and conclusion were developed by authors.
CRediT author statement
Nur Nabilah Mohd Zaini: Conceptualization, investigation, formal analysis, methodology, writing original draft, visualization. Wan Mohd Nuzul Hakimi Wan Salleh: Supervision, resources, writing, review and editing, funding acquisition. Abubakar Siddiq Salihu: Investigation, validation, formal analysis. Nadtanet Nunthaboot: Resoursces, investigation, validation, formal analysis. Nurunajah Ab Ghani: Validation, formal analysis, review and editing. Farkhod Eshboev, Alfred Ngenge Tamfu: Investigation, validation, formal analysis, review and editing. All authors have read and agreed to the published version of the manuscript.
References
[1] S Ferdosh. The extraction of bioactive agents from Calophyllum inophyllum L. and their pharmacological properties. Scientia Pharmaceutica 2024; 92(1), 6.
[2] A Atabani and AD César. Calophyllum inophyllum L. A prospective non-edible biodiesel feedstock. Study of biodiesel production, properties, fatty acid composition, blending and engine performance. Renewable and Sustainable Energy Reviews 2014; 37, 644-655.
[3] HM Alshibl, ES Al-Abdullah, ME Haiba, HM Alkahtani, GEA Awad, AH Mahmoud, BMM Ibrahim, A Bari and A Villinger. Synthesis and evaluation of new coumarin derivatives as antioxidant, antimicrobial, and anti-inflammatory agents, Molecules 2020; 25(14), 3251.
[4] SJ Hamid and T Salih. Design, synthesis, and anti-inflammatory activity of some coumarin Schiff base derivatives: In silico and in vitro study. Drug Design, Development and Therapy 2022; 16, 2275-2288.
[5] KV Sashidhara, M Kumar, RK Modukuri, R Sonkar, G Bhatia, A Khanna, S Rai and R Shukla. Synthesis and anti-inflammatory activity of novel biscoumarin-chalcone hybrids. Bioorganic & Medicinal Chemistry Letters 2011; 21(15), 4480-4484.
[6] A Pribowo, J Girish, M Gustiananda, RG Nandhira and P Hartrianti. Potential of tamanu (Calophyllum inophyllum) oil for atopic dermatitis treatment. Evidence-Based Complementary and Alternative Medicine 2021; 2021(1), 6332867.
[7] Y Kashman, KR Gustafson, RW Fuller, JH Cardellina, JB McMahon, MJ Currens, RW Buckheit, SH Hughes, GM Cragg and MR Boyd. The calanolides, a novel HIV-inhibitory class of coumarin derivatives from the tropical rainforest tree Calophyllum lanigerum. Journal of Medicinal Chemistry 1992; 35(15), 2735-2743.
[8] MP de Béthune. Non-nucleoside reverse transcriptase inhibitors (NNRTIs), their discovery, development, and use in the treatment of HIV-1 infection: A review of the last 20 years (1989-2009). Antiviral Research 2010; 85(1), 75-90.
[9] RW Buckheit, EL White, V Fliakas-Boltz, J Russell, TL Stup, TL Kinjerski, MC Osterling, A Weigand and JP Bader. Unique anti-human immunodeficiency virus activities of the nonnucleoside reverse transcriptase inhibitors calanolide A, costatolide, and dihydrocostatolide. Antimicrobial Agents and Chemotherapy 1999; 43(8), 1827-1834.
[10] NLDL Mata, N Kumarasamy, V Khol, OT Ng, KV Nguyen, TP Merati, TT Pham, MP Lee, N Durier and M Law. Improved survival in HIV treatment programmes in Asia. Antiviral Therapy 2016; 21(6), 517-527.
[11] M Li, F. Yu, B Zhu, J Xiao, C Yan, X Yang, X Liang, F Wang, H Zhang and F Zhang. Interactions between human immunodeficiency virus and human endogenous retroviruses. Journal of Virology 2025; 99(3), e02319.
[12] Z Hosseini, A Ebadi, T Aghamolaei and S Nedjat. A model for explaining adherence to antiretroviral therapy in patients with HIV/AIDS: A grounded theory study. Health & Social Care in the Community 2022; 30(6), e5735.
[13] Q Xiao, D Guo and S Chen. Application of CRISPR/Cas9-based gene editing in HIV-1/AIDS therapy. Frontiers in Cellular and Infection Microbiology 2019; 9, 69.
[14] ND Kolanu. CRISPR-Cas9 gene editing: Curing genetic diseases by inherited epigenetic modifications. Global Medical Genetics 2024; 11(01), 113-122.
[15] H Lin, G Li, X Peng, A Deng, L Ye, L Shi, T Wang and J He. The use of CRISPR/Cas9 as a tool to study human infectious viruses. Frontiers in Cellular and Infection Microbiology 2021; 11, 590989.
[16] AAD Zailan, T Karunakaran, MH Abu Bakar and VJY Mian. The Malaysian genus Calophyllum (Calophyllaceae): A review on its phytochemistry and pharmacological activities. Natural Product Research 2021; 36(17), 4569-4579.
[17] V Flores-Morales, AP Villasana-Ruíz, I Garza-Veloz, S González-Delgado and ML Martinez-Fierro. Therapeutic effects of coumarins with different substitution patterns. Molecules 2023; 28(5), 2413.
[18] OM Tsivileva, OV Koftin and NV Evseeva. Coumarins as fungal metabolites with potential medicinal properties. Antibiotics 2022; 11(9), 1156.
[19] M Lončar, M Jakovljević, D Šubarić, M Pavlić, V Buzjak Služek, I Cindrić and M Molnar. Coumarins in food and methods of their determination. Foods 2020; 9(5), 645.
[20] SD Sarker and L Nahar. Progress in the chemistry of naturally occurring coumarins. Progress in the Chemistry of Organic Natural Products 2017; 106, 241-304.
[21] S Gupta and P Gupta. The genus Calophyllum: Review of ethnomedicinal uses, phytochemistry and pharmacology. In: J Singh, V Meshram and M Gupta (Eds.). Bioactive Natural Products in Drug Discovery. Springer, Singapore, 2020.
[22] T Ma, P Zheng, X Li, X Hong and G Liu. Function, pharmaceutical, and pharmacological research and development of natural tetracyclic dipyranocoumarin (+)-calanolide A and its analogs. In: G Liu (Ed.). Medicinal chemistry in drug development. Elsevier, Amsterdam, Netherland, 2025, p. 651-687.
[23] RK Arora, N Kaur, Y Bansal and G Bansal. Novel coumarin-benzimidazole derivatives as antioxidants and safer anti-inflammatory agents. Acta Pharmaceutica Sinica B 2014; 4(5), 368-375.
[24] A Stefanachi, F Leonetti, L Pisani, M Catto and A Carotti. Coumarin: a natural, privileged and versatile scaffold for bioactive compounds. Molecules 2018; 23(2), 250.
[25] NS Firouza, T Karunakaran, N Mokhtar, R Santhanam, VJY Mian and MH Abu Bakar. Chemical constituents from the stem barks of Calophyllum recurvatum P.F. Stevens and Calophyllum andersonii P.F. Stevens and their in vitro hepatotoxic activity. Natural Product Research 2024; 39(9), 2587-2593.
[26] AD Patil, AJ Freyer, DS Eggleston, RC Haltiwanger, MF Bean, PB Taylor, MJ Caranfa, AL Breen, HR Bartus and RK Johnson. The inophyllums, novel inhibitors of HIV-1 reverse transcriptase isolated from the Malaysian tree Calophyllum inophyllum Linn. Journal of Medicinal Chemistry 1993; 36(26), 4131-4138.
[27] S Cao, X Wu, K Sim, BH Tan, JJ Vittal, JT Pereira and S Goh. Minor coumarins from Calophyllum teysmannii var. inophylloide and synthesis of cytotoxic calanone derivatives. Helvetica Chimica Acta 1998; 81(5-8), 1404-1416.
[28] TC McKee, CD Covington, RW Fuller, HR Bokesch, S Young, JH Cardellina, MR Kadushin, DD Soejarto, PF Stevens, GM Cragg and MR Boyd. Pyranocoumarins from tropical species of the genus Calophyllum: a chemotaxonomic study of extracts in the National Cancer Institute collection. Journal of Natural Products 1998; 61(10), 1252-1256.
[29] KR Gustafson, HR Bokesch, RW Fuller, JH Cardellina, MR Kadushin, DD Soejarto and MR Boyd. Calanone, a novel coumarin from Calophyllum teysmannii. Tetrahedron Letters 1994; 35(32), 5821-5824.
[30] MB Zakaria, Vijayasekaran, Z Ilham and NA Muhamad. Anti-inflammatory activity of Calophyllum inophyllum fruit extracts. Procedia Chemistry 2014; 13, 218-220.
[31] NI Aminudin, F Ahmad, M Taher and RM Zulkifli. α-Glucosidase and 15-lipoxygenase inhibitory activities of phytochemicals from Calophyllum symingtonianum. Natural Product Communications 2015; 10(9), 1585-1587.
[32] TC McKee, RW Fuller, CD Covington, JH Cardellina, RJ Gulakowski, BL Krepps and JB McMahon, MR Boyd. New pyranocoumarins isolated from Calophyllum lanigerum and Calophyllum teysmannii. Journal of Natural Products 1996; 59(8), 754-758.
[33] T Ishikawa. Anti-HIV-1 active Calophyllum coumarins: distribution, chemistry, and activity. Heterocycles 2000; 53(2), 453-474.
[34] M Huerta-Reyes, MC Basualdo, F Abe, M Jimenez-Estrada, C Soler and R Reyes-Chilpa. HIV-1 inhibitory compounds from Calophyllum brasiliense leaves. Biological and Pharmaceutical Bulletin 2004; 27(9), 1471-1475.
[35] S Cao, K Sim, J Pereira and S Goh. Coumarins from Calophyllum teysmannii (Guttiferae). Phytochemistry 1998; 47(5), 1051-1055.
[36] N Mokhtar, T Karunakaran, R Santhanam, MH Abu Bakar and VJY Mian. Phenolics and triterpenoids from stem bark of Calophyllum lanigerum var. austrocoriaceum (Whitmore) P.F. Stevens and their cytotoxic activities. Natural Product Research 2024; 38(5), 873-878.
[37] F Laure, P Raharivelomanana, J Butaud, J Bianchini and EM Gaydou. Screening of anti-HIV-1 inophyllums by HPLC-DAD of Calophyllum inophyllum leaf extracts from French Polynesia islands. Analytica Chimica Acta 2008; 624(1), 147-153.
[38] SP Joshi, VB Deodhar and UD Phalgune. ChemInform abstract: a new coumarin from the seeds of Calophyllum inophyllum Linn. Indian Journal of Chemistry 2000; 39, 560-561.
[39] SP Joshi, SR Kulkarni, UD Phalgune and VG Puranik. New dipyranocoumarin from the leaves of Calophyllum apetalum Willd. Natural Product Research 2013; 27, 1896-1901.
[40] D Guilet, JJ Hélesbeux, D Séraphin, T Sévenet, P Richomme and J Bruneton. Novel cytotoxic 4-phenylfuranocoumarins from Calophyllum dispar. Journal of Natural Products 2001; 64(5), 563-568.
[41] D Guilet, D Séraphin, D Rondeau, P Richomme and J Bruneton. Cytotoxic coumarins from Calophyllum dispar. Phytochemistry 2001; 58(4), 571-575.
[42] NH Zamakshshari, GCL Ee, SS Teh, SK Daud and I Safinar. Natural product compounds from Calophyllum depressinervosum. Pertanika Journal of Tropical Agricultural Science 2016; 39(2), 249-255.
[43] SB Daud, GC Ee, EA Malek, SS The and I See. A new coumarin from Calophyllum hosei. Natural Product Research 2014; 28(19), 1534-1538.
[44] CK Lim, S Hemaroopini, SY Gan, SM Loo, JR Low, VY Jong, HC Soo, CO Leong, CW Mai and CF Chee. In vitro cytotoxic activity of isolated compounds from Malaysian Calophyllum species. Medicinal Chemistry Research 2016; 25(8), 1686-1694.
[45] FL Yong. 2015, Phytochemical and antioxidant studies of Calophyllum sclerophyllum. Bachelor’s Thesis. Universiti Tunku Abdul Rahman, Perak, Malaysia.
[46] IA Noh, VJY Mian. Phytochemicals, antimicrobials and antioxidants studies of the stem bark extract from Calophyllum ferrugineum. Scientific Research Journal 2020; 17(2), 1-12.
[47] G Ee, K Ng, Y Taufiq-Yap, M Rahmani, A Ali and R Muse. Mucigerin, a new coumarin from Calophyllum mucigerum (Guttiferae). Natural Product Research 2004; 18(2), 123-128.
[48] NI Aminudin, F Ahmad, M Taher and RM Zulkifli. Incrassamarin A-D: Four new 4-substituted coumarins from Calophyllum incrassatum and their biological activities. Phytochemistry Letters 2016; 16, 287-293.
[49] GC Ee, SH Mah, SS Teh, M Rahmani, R Go and YH Taufiq-Yap. Soulamarin, a new coumarin from stem bark of Calophyllum soulattri. Molecules 2011; 16(11), 9721-9727.
[50] KH Tee, GC Ee, IS Ismail, T Karunakaran, SS Teh, VY Jong and SM Mohd Nor. A new coumarin from stem bark of Calophyllum wallichianum. Natural Product Research 2018; 32(21), 2565-2570.
[51] MSM Sahimi, GCL Ee, SS Teh, AAF Ismail and MA Sukari. Chemical constituents of Calophyllum benjaminum and Calophyllum javanicum and their bioactivities. Open Conference Proceedings Journal 2013; 4, 127.
[52] CK Lim, YP Ham, LQ Lim and VY Jong. 4-Alkylcoumarins and a phloroglucinol from the stem bark of Calophyllum gracilentum. Phytochemistry Letters 2019; 30, 99-102.
[53] AS Salihu, WMNHW Salleh and T Ogunwa. Computational exploration of flavonoids from the genus Knema with anti-inflammatory potential. Journal of the Serbian Chemical Society 2024; 89(7-8), 1039-1051.
[54] PC Agu, CA Afiukwa, OU Orji, EM Ezeh, IH Ofoke, CO Ogbu, EI Ugwuja and PM Aja. Molecular docking as a tool for the discovery of molecular targets of nutraceuticals in diseases management. Scientific Reports 2023; 13(1), 40160.
[55] MS Bilal, SA Ejaz, S Naseem, PA Channar, A Saeed, S Zargar, R Ujan, R Sahito, Q Abbas and TA Wani. Synthesis, in vitro evaluation and computational modelling of benzene sulfonamide derivatives as Dickkopf-1 inhibitors for anticancer drug development. Scientific Reports 2025; 15(1), 68901.
[56] CA Lipinski. Lead- and drug-like compounds: The rule-of-five revolution. Drug Discovery Today: Technologies 2004; 1(4), 337-341.
[57] A Daina, O Michielin and V Zoete. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Scientific Reports 2017; 7(1), 42717.
[58] DE Pires, TL Blundell and DB Ascher. pkCSM: Predicting small-molecule pharmacokinetic and toxicity properties using graph-based signatures. Journal of Medicinal Chemistry 2015; 58(9), 4066-4072.
[59] HR Dharmaratne, WM Wanigasekera, E Mata-Greenwood and JM Pezzuto. Inhibition of human immunodeficiency virus type-1 reverse transcriptase activity by cordatolides isolated from Calophyllum cordato-oblongum. Planta Medica 1998; 64(05), 460-461.
[60] MC Yimdjo, AG Azebaze, AE Nkengfack, AM Meyer, B Bodo and ZT Fomum. Antimicrobial and cytotoxic agents from Calophyllum inophyllum. Phytochemistry 2004; 65(20), 2789-2795.
[61] T Pengsuparp, M Serit, SH Hughes, DD Soejarto and JM Pezzuto. Specific inhibition of human immunodeficiency virus type-1 reverse transcriptase mediated by soulattrolide, a coumarin isolated from the latex of Calophyllum teysmannii. Journal of Natural Products 1996; 59(9), 839-842.
[62] L Nahar, AD Talukdar, D Nath, S Nath, A Mehan, FMD Ismail and SD Sarker. Naturally occurring calanolides: Occurrence, biosynthesis, and pharmacological properties including therapeutic potential. Molecules 2020; 25(21), 4983.
[63] C Ito, M Itoigawa, Y Mishina, VC Filho, F Enjo, H Tokuda, H Nishino and H Furukawa. Chemical constituents of Calophyllum brasiliense. 2. Structure of three new coumarins and cancer chemopreventive activity of 4-substituted coumarins. Journal of Natural Products 2003; 66(3), 368-371.
[64] HR Dharmaratne, JR Mayuri Sajeevani, GP Marasinghe and EMS Ekanayake. Distribution of pyranocoumarins in Calophyllum cordato-oblongum. Phytochemistry 1998; 49(4), 995-998.
[65] V Nguyen, C Truong, BC Nguyen, TV Vo, T Dao, V Nguyen, DT Trinh, HK Huynh and C Bui. Anti-inflammatory and wound healing activities of calophyllolide isolated from Calophyllum inophyllum Linn. PloS One 2017; 12(10), e0185674.
[66] C Ito, T Murata, M Itoigawa, K Nakao, N Kaneda and H Furukawa. Apoptosis-inducing activity of 4-substituted coumarins from Calophyllum brasiliense in human leukaemia HL-60 cells. Journal of Pharmacy and Pharmacology 2006; 58(7), 975-980.
[67] RC Saxena, R Nath, G Palit, SK Nigam and KP Bhargava. Effect of calophyllolide, a nonsteroidal anti-inflammatory agent, on capillary permeability. Planta Medica 1982; 44(04), 246-248.
[68] N Jaiswal, N Gunaganti, CK Maurya, T Narender and AK Tamrakar. Free fatty acid-induced impairment of insulin signaling is prevented by the diastereomeric mixture of calophyllic acid and isocalophyllic acid in skeletal muscle cells. European Journal of Pharmacology 2015; 746, 70-77.
[69] J Prasad, A Shrivastava, AK Khanna, G Bhatia, SK Awasthi and T Narender. Antidyslipidemic and antioxidant activity of the constituents isolated from the leaves of Calophyllum inophyllum. Phytomedicine 2012; 19(14), 1245-1249.
[70] F Zafar, A Gupta, K Thangavel, K Khatana, AA Sani, A Ghosal, P Tandon and N Nishat. Physicochemical and pharmacokinetic analysis of anacardic acid derivatives. ACS Omega 2020; 5(11), 6021-6030.