Trends
Sci.
2026;
23(8):
13191
Chlorogenic Acid Enhanced Testosterone-Mediated Libido Through Improved Glycemic Control and Redox Homeostasis in Diabetic Male Rats
Eko
Naning Sofyanita1,2,*,
![]() ,
Ari Suwondo3,
Heri Nugroho4,
,
Ari Suwondo3,
Heri Nugroho4,
Diana Nur Afifah5, Suhartono Suhartono6 and Achmad Zulfa Juniarto7
1Doctoral Study Program of Medical and Health Science, Universitas Diponegoro, Jawa Tengah, Indonesia
2Bachelor of Applied Medical Laboratory Technology, Poltekkes Kemenkes Semarang, Jawa Tengah, Indonesia
3Postgraduate Program, Poltekkes Kemenkes Semarang, Jawa Tengah, Indonesia
4Division of Endocrinology, Metabolism and Diabetes, Department of Internal Medicine, Faculty of Medicine, Universitas Diponegoro/Dr.Kariadi General Hospital, Jawa Tengah, Indonesia
5Department of Nutrition Science, Faculty of Medicine, Universitas Diponegoro, Jawa Tengah, Indonesia
6Department of Environmental Health, Faculty of Public Health, Universitas Diponegoro, Jawa Tengah, Indonesia
7Department of Medical Biology, Faculty of Medicine, Universitas Diponegoro, Jawa Tengah, Indonesia
(*Corresponding author’s e-mail: [email protected])
Received: 29 December 2025, Revised: 5 February 2026, Accepted: 12 February 2026, Published: 25 March 2026
Abstract
Type 2 diabetes mellitus (T2DM) induces chronic metabolic disruption characterized by sustained hyperglycemia, oxidative–antioxidant imbalance, and impaired steroidogenesis, resulting in progressive deterioration of male reproductive and sexual function. Chlorogenic acid (CGA) is a polyphenolic compound with metabolic and antioxidant activities, and has shown potential benefits in mitigating diabetes associated reproductive dysfunction. This study evaluated the effects of CGA on glycemic control, oxidative stress, testosterone levels, and libido parameters in male Wistar rats with T2DM. Thirty rats were divided into six groups (n = 5 per group): healthy controls, diabetic controls, diabetic + metformin, and diabetic treated with CGA at 6.25, 12.5, and 25 mg/kg BW. Glycated hemoglobin A1c (HbA1c), malondialdehyde (MDA), superoxide dismutase (SOD), and testosterone were measured on day 32. Sexual behavior was recorded over three consecutive days. CGA demonstrated a dose-dependent improvement across all biological parameters. The highest CGA dose is 25 mg/kg BW and can reduce HbA1c levels, decrease MDA, and increase SOD, testosterone levels and libido activity close to those of normal controls. Correlation analyses revealed strong relationships between glicemic control, oxidative, hormonal, and libido variables. In conclusion, CGA enhances glycemic control and redox balance, restores testosterone, and improves libido-related behavior in diabetic rats, with broader restorative effects compared to metformin. CGA represents a promising adjunctive strategy for addressing diabetes-related reproductive dysfunction.
Keywords: Type 2 diabetes mellitus, Chlorogenic acid, HbA1c, Oxidative stress, SOD, Testosterone, Libido
Introduction
Diabetes mellitus (DM) is a chronic metabolic disorder characterized by persistent hyperglycemia resulting from impaired insulin secretion, insulin action, or both. According to the International Diabetes Federation, the global prevalence of diabetes reached 537 million adults in 2021, with type 2 diabetes mellitus
(T2DM) accounting for approximately 90% of all cases worldwide [1]. The burden of T2DM continues to increase globally, including in developing countries, and is associated with a wide range of chronic complications that significantly impair quality of life and increase morbidity and mortality [2,3].
The pathogenesis of T2DM involves insulin resistance in peripheral tissues combined with progressive pancreatic β-cell dysfunction. Prolonged hyperglycemia activates multiple intracellular pathways that disrupt vascular integrity, neural function, and endocrine regulation, leading to systemic complications affecting multiple organs [4,5]. Among these complications, reproductive and sexual disorders in men have gained increasing attention due to their substantial impact on physical, psychological, and social well-being [6].
Male sexual dysfunction is a common yet underrecognized complication of diabetes. Epidemiological studies report that approximately 40% - 50% of men with diabetes experience sexual dysfunction, including erectile dysfunction, ejaculatory disorders, and reduced libido, with prevalence increasing with age and duration of disease [7,8]. Libido, defined as sexual motivation or desire, represents a complex neuroendocrine process regulated by the interaction of hormonal balance, neural signaling, vascular function, and central nervous system activity. In men with T2DM, reduced libido is frequently associated with impaired testicular function, disruption of the hypothalamic–pituitary–gonadal (HPG) axis, and decreased testosterone synthesis [9].
Experimental studies using diabetic rodent models consistently demonstrate reductions in sexual motivation, reflected by decreased frequencies of sexual behaviors such as anogenital investigation, mounting or climbing, and intromission or coitus [10]. These behaviors are commonly assessed using validated parameters of male sexual activity and are considered sensitive indicators of libido and reproductive competence [11,12]. Behavioral impairment in diabetic animals is thought to result from the combined effects of metabolic dysregulation, vascular insufficiency, neuropathy, and endocrine disturbances that collectively impair both central and peripheral mechanisms of sexual motivation [13].
Metformin remains the first-line pharmacological therapy for T2DM and has demonstrated efficacy in improving insulin sensitivity and metabolic control. Beyond its antidiabetic action, metformin has been reported to exert beneficial effects on male reproductive health in experimental studies, including improvement of spermatogenesis, testicular morphology, and gonadotropin signaling [14]. The beneficial effects of metformin on male reproductive parameters appear to be largely mediated by activation of AMP-activated protein kinase (AMPK) and modulation of mitochondrial metabolism, which enhance metabolic control and steroidogenic signaling. However, long-term pharmacological therapy may be limited by adverse effects such as gastrointestinal intolerance and nutrient malabsorption, and may not fully restore reproductive and sexual function, prompting interest in complementary or alternative bioactive compounds [15].
Chlorogenic acid (CGA) is a naturally occurring polyphenolic compound abundantly found in coffee and various medicinal plants. Increasing evidence suggests that CGA exerts broad biological activities related to metabolic regulation, cellular signaling, and tissue protection [16]. In experimental diabetic models, CGA administration has been shown to improve metabolic disturbances and preserve reproductive tissue integrity, with reported benefits on testicular function and hormone synthesis [17]. Compared with metformin, CGA exhibits multi-target metabolic actions, including improvement of glucose tolerance, insulin sensitivity, modulation of AMPK signaling, and anti-inflammatory effects, while also demonstrating potential ameliorative impacts on diabetes-associated reproductive dysfunction in male animal models. These properties make CGA a promising adjunctive or alternative strategy for managing reproductive complications of T2DM [18].
Despite growing evidence supporting the metabolic and reproductive benefits of CGA, important research gaps remain. Most existing studies focus primarily on biochemical or histological outcomes, while comprehensive evaluation of libido-related behavioral parameters in diabetic conditions is limited. Moreover, direct comparison between CGA and established antidiabetic therapy, such as metformin, in the context of male sexual motivation is scarce. In addition, dose-response effects and longer treatment durations using validated behavioral indicators have not been adequately explored. Therefore, further investigation is warranted to evaluate the effectiveness of CGA in improving male libido in a T2DM rat model and to clarify its potential role relative to conventional therapy.
Materials and methods
Drug and chemicals
CGA with a purity of ≥95% was obtained from Sigma-Aldrich (St. Louis, MO, USA; product number C3878; batch WXBF3738V), streptozotocin (STZ) (Sigma-Aldrich, USA), nicotinamide (NA) (Solarbio N8070, China), administered estradiol benzoate (EB) (Sigma-Aldrich, St. Louis, MO, USA) and progesterone (Sigma-Aldrich, St. Louis, MO, USA).
Experimental Animal
This experimental study employed male Wistar rats weighing 150 - 200 g that were confirmed to be in good health. All animals were housed in standard conditions of temperature 23 ± 2 °C, relative humidity of 55 ± 5% and a controlled environment with a 12 h light/dark cycle. They were fed a standard pellet diet with water available ad libitum. Before the experimental procedures, the rats were acclimated to the laboratory environment for 7 days. The experimental workflow followed a defined timeline. Rats were acclimated for 7 days before receiving a high-fat diet for 14 days. Type 2 diabetes was induced with streptozotocin–nicotinamide, and hyperglycemia was confirmed. Metformin or chlorogenic acid was administered daily for 31 consecutive days. Sexual behavior assessments were conducted on days 29 - 31, and biochemical analyses were performed on day 32. A schematic overview of the experimental workflow is presented in the graphical abstract. The experimental protocol was approved by the Health Research Ethics Committee of Universitas Diponegoro (No. 031/EC/KEPK/KF-UNDIP/II/2024).
Animal groups
A total of 30 rats were used in this study, randomly divided into six groups with five rats each. Group I (K0) included healthy control rats, Group II (KN) consisted of T2DM rats without any treatment, Group III (KP) involved T2DM rats treated with 500 mg/kg body weight (BW) of metformin, Group IV (P1) comprised T2DM rats given chlorogenic acid (CGA) at 6.25 mg/kg BW, Group V (P2) included T2DM rats receiving 12.5 mg/kg BW of CGA, and Group VI (P3) consisted of T2DM rats treated with 25 mg/kg BW of CGA. CGA solutions were freshly prepared by dissolving the compound in phosphate-buffered saline (PBS, pH 7.4) before each use. The CGA was administered intraperitoneally once daily for 31 consecutive days. The intraperitoneal route was chosen to ensure accurate dosing and consistent systemic absorption, reducing variability from gastrointestinal absorption and first-pass hepatic metabolism. This method is commonly employed in experimental studies assessing the biological effects of polyphenolic compounds like CGA in models of metabolic and male reproductive health disorders [19].
Baseline measurement of sex hormone levels and sexual behavior was not performed prior to the intervention to prevent alterations in behavior due to stress, habituation, or experience-dependent changes that can influence libido and endocrine responses in male rats. Repeated testing for sexual behavior can alter motivation and copulatory responses. Therefore, a post-test-only design using different treatment groups, a standard approach in research on male sexual behavior and reproductive endocrine responses system [12].
To induce type 2 diabetes, rats were fed a high-fat diet (HFD) for two weeks, at a rate of 20 g per rat each day. The HFD consisted of Comfeed Par S (60%; Japfa Comfeed Indonesia), wheat flour (27.8%), cholesterol (2%), folic acid (0.2%), and pork fat (10%). After the dietary period, the rats fasted overnight for 12 h before receiving a single intraperitoneal injection of STZ at 45 mg/kg BW, dissolved in 0.1 M citrate buffer (pH 4.5), along with NA at 110 mg/kg BW. Seven days post-STZ–NA injection, blood glucose levels (BGLs) were measured using a glucometer (EasyTouch®, GCU Model: ET-301, Bioptik Technology, Taiwan) to verify stable diabetic conditions. Rats with fasting blood glucose levels exceeding 200 mg/dL were classified as diabetic and included in the study.
Analysis of HbA1c, MDA, SOD, and testosterone levels
Blood samples were collected via the retro-orbital sinus into non-EDTA tubes. They were kept at room temperature for about 30 min to allow clotting, then centrifuged at 3000 rpm for 15 min to obtain serum. On day 32, levels of HbA1c, malondialdehyde (MDA), superoxide dismutase (SOD), and testosterone were measured using commercial ELISA kits (FineTest®, Wuhan National Bio-Industry Base, China). The kits used included HbA1c (ER1030), MDA (ER1878), SOD (ER1950), and Testosterone (EU0400). All procedures followed the manufacturer’s instructions. After the assays, absorbance was read with a Medical Pro Reader-96 microplate reader (Germany), and biomarker levels were calculated from the standard curves.
Test for libido
Before testing sexual behavior, the estrous cycles of female rats were monitored to confirm their reproductive status and synchronize their estrus phase readiness. Estrus synchronization was performed to obtain sexually receptive females and to minimize variability related to spontaneous estrous cycling. Female rats were administered estradiol benzoate at a dose of 20 µg/kg body weight, 48 h before behavioral testing. This EB priming protocol has been widely used to induce estrus and enhance sexual receptivity in female rodents without causing excessive estrogenic stimulation [20,21]. To further optimize receptivity and facilitate copulatory behavior, progesterone was administered at a dose of 0.5 mg/kg body weight, 4 h before the behavioral observation. The combined use of estradiol benzoate followed by progesterone is a well-established method for synchronizing estrus and ensuring lordosis behavior in female rats during sexual behavior testing [22]. Estrus receptivity was confirmed before pairing based on behavioral acceptance of the male.
Libido was assessed by pairing each male rat with an estrus-synchronized female rat. The experimental duration was set at 31 days, based on previous evidence indicating that a significant decline in circulating testosterone levels occurs approximately 14 days after diabetes induction in male rat models, which may adversely affect sexual motivation and performance [23]. Libido observations were conducted on days 29 to 31 at 18:00 (6:00 PM), corresponding to the animals’ active (dark) phase, under dim light conditions to minimize stress-related behavioral interference. During each observation session, a female rat was first placed into the observation cage and allowed to acclimatize for 10 min. Subsequently, the male rat was introduced, and sexual behavior was recorded continuously for a 30-minute period. Sexual behaviors were directly observed and quantified using standardized behavioral parameters commonly employed in rodent sexual behavior studies [24,25]. Libido-related behavior was evaluated using the following parameters: Introduction as an anogenital investigation, defined as the male rat approaching and investigating the female’s anogenital region, including sniffing and licking, which reflects sexual motivation and appetitive behavior. Climbing or mount frequency is defined as the number of times the male rat mounts the female without penile insertion, representing consummatory sexual behavior and sexual performance. Coitus frequency defined intromission as successful penile insertion during mounting, characterized by rhythmic pelvic thrusting and a stereotypical copulatory posture, serving as a key indicator of effective copulatory behavior [26-28].
Libido was recorded for 30 min per session over three consecutive days to improve the reliability and reproducibility of behavioral measurements. Each occurrence of anogenital investigation and mount behavior was assigned a score of one, allowing quantitative comparison of libido-related activity among experimental groups. This behavioral assessment framework is widely recognized as a sensitive and valid method for evaluating male sexual motivation and copulatory performance in experimental models of metabolic and endocrine disorders.
The selection of day 32 as the study endpoint was based on previous evidence indicating that testosterone reduction and oxidative stress become apparent as early as 14 days after STZ–NA induction, while behavioral and endocrine disturbances reach a more stable state after approximately 3 - 4 weeks. Although a complete spermatogenic cycle in rats requires approximately 48 - 52 days [27], the present timeframe was sufficient to evaluate functional hormonal alterations and libido-related behavioral outcomes, which were the primary objectives of this study.
Statistical analysis
Data are expressed as mean ± standard deviation (SD). All statistical analyses were conducted using SPSS software version 31 (IBM Corp., USA). Comparisons among experimental groups were performed using one-way analysis of variance (ANOVA) when the data were normally distributed and homogeneous, followed by Tukey’s honestly significant difference (HSD) post hoc test to identify pairwise differences. A p-value of less than 0.05 was considered statistically significant. Associations between variables were evaluated using Pearson’s correlation analysis, and linear regression was applied to further assess the strength and direction of these relationships. Graphical representations were generated using GraphPad Prism version 10.6.1.
Results and discussion
HbA1c, SOD, MDA, and testosterone levels in diabetic rats given CGA
In six groups of rats induced with DM and treated with either CGA or metformin, there were significant differences in HbA1c levels among the groups (p = 0.000). The K0 group had the lowest average HbA1c level at 19.50 ± 1.90 ng/mL, while the KN group showed the highest HbA1c level at 60.02 ± 1.30 ng/mL. The KP group experienced a decrease in HbA1c to 36.50 ± 1.10 ng/mL compared to the KN group. The P1 treatment group had an HbA1c level of 41.00 ± 0.90 ng/mL. A more noticeable decrease in HbA1c was observed in groups P2 and P3, with values of 30.60 ± 2.03 ng/mL and 24.00 ± 0.80 ng/mL, respectively. Overall, HbA1c levels decreased with increasing doses of chlorogenic acid (Figure 1).
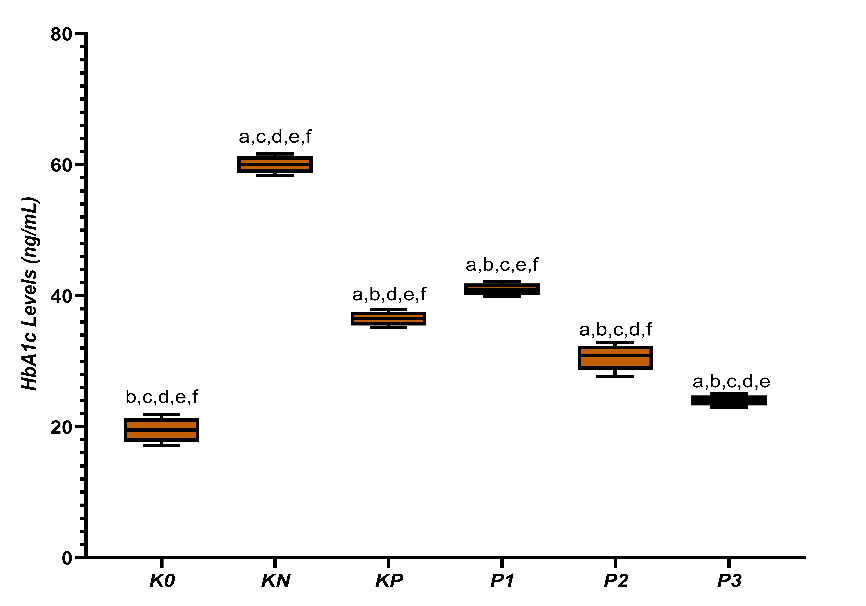
Figure 1 Box-plot distribution of HbA1c levels (ng/mL) across experimental groups (K0 = normal control; KN = T2DM rats; KP = metformin treatment; P1–P3 = CGA treatment doses). a, b, c, d, e, f denote statistical differences between indicated groups, p < 0.05).
Low HbA1c levels characterize good glycemic control and play a crucial role in preventing long-term complications. HbA1c reflects the average blood glucose over 2 - 3 months and serves as a key indicator in diabetes management. The reduction in HbA1c levels after administration of CGA in this study indicates dose-dependent improvement in glycemic control [29]. This decrease suggests reduced glucotoxicity, which contributes to the activation of inflammatory pathways and oxidative stress. CGA has been reported to increase insulin sensitivity and glucose tolerance by enhancing peripheral glucose utilization.
Activation of AMPK by CGA suppresses hepatic glucose production, increases fatty acid oxidation, and supports glucose uptake by muscle tissues. In skeletal muscle, AMPK activation increases the expression of gamma coactivator 1-alpha (PGC-1α) and peroxisome proliferator-activated receptor alpha (PPARα), thereby supporting energy metabolism [30]. CGA also enhances Akt phosphorylation and facilitates Glucose transporter type 4 (GLUT4) translocation to the cell membrane, accelerating glucose utilization. These metabolic improvements reduce chronic glucose exposure to proteins, thereby inhibiting the formation of damaging advanced glycation end-products (AGEs) [31]. Thus, the HbA1c improvements observed in this study reflect an integrated enhancement of insulin response, reprogramming of energy metabolism, and reduction of metabolic stress related to hyperglycemia. Zalewska et al. [32] Further demonstrated improved glucose handling and insulin-related pathways mediated by CGA.
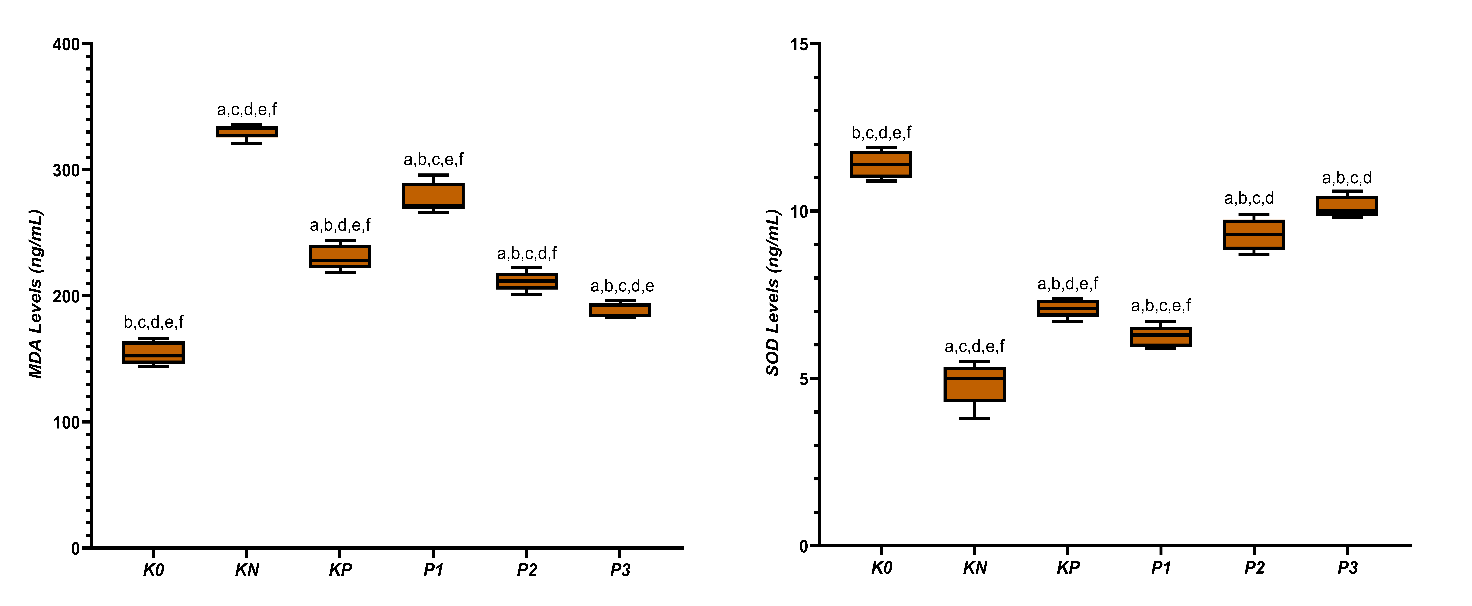
Figure 2 Averages of MDA and SOD levels in diabetic rats given CGA. a,b,c,d,e,f Superscript in the row signify statistically meaningful differences (p < 0.05).
The average MDA levels (Figure 2) showed that Group K0 had the lowest MDA level at 154.60 ± 9.23 ng/mL, followed by Group P3 at 183.70 ± 5.98 ng/mL, Group P2 at 211.62 ± 7.70 ng/mL, Group KP at 230.60 ± 9.97 ng/mL, Group P1 at 277.82 ± 12.00 ng/mL, and Group KN showed the highest increase in MDA levels at 330.62 ± 5.80 ng/mL. The decreasing trend of MDA appears to be in line with increasing doses of chlorogenic acid, indicating a significant difference between groups based on the ANOVA test (p < 0.05). These results suggest a clear difference in lipid peroxidation levels among the groups The increase in SOD levels and testosterone is shown in Table 1. The treatment groups gradually increased in tandem with the rising doses of chlorogenic acid. The SOD activity, which was initially very low in the KN group (4.86 ± 0.65 ng/mL), increased in the KP group (7.10 ± 0.27 ng/mL), then continued to P1 (6.26 ± 0.32 ng/mL), P2 (9.30 ± 0.47 ng/mL), and reached a high value in P3 (10.12 ± 0.33 ng/mL), approaching the normal control group K0 (11.40 ± 0.41 ng/mL).
High levels of MDA in the diabetic group reflect increased lipid peroxidation due to excess reactive oxygen species (ROS). The testes are susceptible to peroxidation because of their high content of polyunsaturated fatty acids in cell membranes. The decrease in MDA after administering CGA indicates a reduction in oxidative damage associated with hyperglycemia. Hyperglycemia increases ROS through disrupted mitochondrial electron transport and activation of advanced glycation end-products (AGE) receptor for AGE (RAGE) [33]. Reactive aldehyde products like MDA can bind to proteins and DNA, altering their structure and function. The greater reduction in MDA at higher doses of CGA suggests a stronger antioxidant activity. This improvement is related to decreased lipid peroxidation, which can disrupt membrane homeostasis and cell function. By suppressing MDA formation, CGA helps maintain membrane integrity and stabilize cellular signaling systems. The reduction in MDA aligns with the recovery of mitochondrial function, supporting energy utilization and cell survival. Through controlling lipid peroxidation, CGA contributes to preserving the structure and function of tissues vulnerable to chronic oxidative stress. Maske et al. [34] also reported increased lipid peroxidation in the testes of STZ-NA induced diabetic rats. The strongest decrease in MDA at higher CGA doses confirms its potent antioxidant activity.
The decrease in SOD in diabetic conditions reflects weakened endogenous antioxidant defenses due to increased ROS. Reduced SOD activity increases cellular exposure to superoxide and accelerates the formation of peroxynitrite. The recovery of SOD after CGA administration indicates an enhanced capacity to neutralize reactive oxygen species. Activation of the Nrf2 pathway by CGA has been reported to increase the expression of antioxidant enzymes, including SOD. The increase in SOD helps maintain redox balance by reducing the flow of radicals that trigger lipid peroxidation. Improved antioxidant capacity supports mitochondrial membrane stability and maintains membrane potential. SOD recovery inhibits oxidative damage that can disrupt metabolic enzymes and structural proteins [35]. Nguyen et al. [30] further highlight the role of CGA in strengthening the antioxidant system in diabetes. The parallel increase in SOD and decrease in MDA observed here support coordinated redox protection. The balance between the decrease in MDA and the increase in SOD reflects the recovery of redox homeostasis. Oxidative imbalance is a primary mediator of diabetic testicular damage. Aitken et al. [36] identified oxidative stress as a main driver of reproductive dysfunction in diabetes. By restoring redox balance, CGA limits cellular injury and maintains tissue function. Dose-dependent redox improvement aligns with hormonal recovery, supporting causality. Redox homeostasis thus represents a mechanistic bridge between metabolic control and endocrine recovery [37].
A similar pattern of increase was observed in testosterone levels (Figure 3), where the KN group showed the lowest level (812.59 ± 54.90 pg/mL), then increased in the KP group (1202.96 ± 55.67 pg/mL), and gradually rose in P1 (996.31 ± 13.32 pg/mL), P2 (1560.39 ± 21.45 pg/mL), reaching a value close to the normal control in P3 (1644.35 ± 34.71 pg/mL) compared to K0 (1765.23 ± 24.95 pg/mL). ANOVA tests showed that the increases in SOD and testosterone among groups were significant (p < 0.05), and the Tukey test results confirmed that these differences were consistent at each dose level.
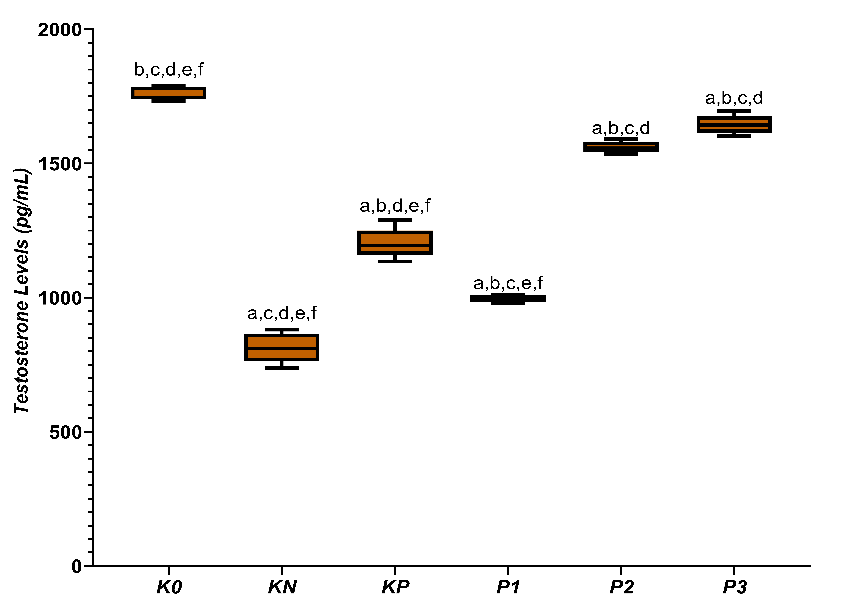
Figure 3 Average testosterone levels in diabetic rats treated with CGA. a,b,c,d,e,f superscripts in the row indicate statistically significant differences (p < 0.05) among groups K0, KN, KP, P1, P2, and P3.
The decrease in testosterone in diabetes control reflects Leydig cell steroidogenesis disorder. Hyperglycemia and oxidative stress disrupt mitochondrial function and steroidogenic enzymes. Further oxidative damage promotes Leydig cell apoptosis [38]. Sari et al. [39] reported a strong relationship between oxidative stress and androgen deficiency in men with diabetes. CGA significantly restores testosterone levels, especially at higher doses. This recovery indicates an increased steroidogenic capacity. Testosterone functions as a functional marker of testicular recovery. Steroidogenesis is highly sensitive to oxidative stress, making redox balance crucial for testosterone synthesis [40]. Reduction of ROS maintains the integrity of Leydig cell mitochondria and enzymatic activity.
Both CGA and metformin improve glycemic and oxidative parameters; however, CGA produces a stronger recovery of testosterone at higher doses. Metformin primarily targets liver glucose production and insulin sensitivity. Yan et al. [41] reported comparable hypoglycemic effects between CGA and metformin. However, reproductive outcomes with metformin vary depending on the metabolic context. Tseng et al. [42] emphasized that the endocrine effects of metformin depend on the model. As a polyphenolic antioxidant, CGA may offer additional tissue-specific protection in the testes. Therefore, CGA could provide a complementary advantage in maintaining reproductive endocrine function.
Libido assessment in T2DM rats given CGA
In Figure 4. Parameter introduction shows the variation in response among treatment groups. The highest mean value was found in group K0 (222.80 ± 7.19), followed by P3 (212.40 ± 6.58) and P2 (188.00 ± 7.13). Meanwhile, the KN group showed the lowest value (70.0 ± 3.61), which is much lower than the other groups, indicating a decrease in introduction activity under disease conditions. The KP group (140.0 ± 7.62) and P1 (94.2 ± 2.28) showed increased values compared to KN, but still below K0. The Tukey post-hoc test revealed that all pairwise group differences were significant, except for K0 compared to P3, which showed no significant difference. These findings indicate that gradually increasing the intervention dose results in an increased introduction response. In the highest dose group (P3), the response reaches a level comparable to the healthy group K0.
In climbing observations, group K0 showed the highest activity with an average of 55.00 ± 1.58. Conversely, the KN group exhibited a sharp decline to 5.00 ± 0.71, indicating a significant disturbance in sexual behavior due to the disease condition. The group receiving treatment showed a graded response: KP and P1 remained far below the healthy group, but further improvement was observed in P2, and the closest recovery to the normal group was achieved in P3 (39.80 ± 1.48). Overall, the Tukey test results confirm that the differences between groups are statistically significant (p < 0.05).
For the coitus parameter, the observed pattern differs from climbing. Although the K0 group still shows the highest value (12.80 ± 1.48), the variation between groups is wider, and the recovery trend is not as strong as that of other parameters. KN is at the lowest point (0.80 ± 0.84), while the treatment groups (KP, P1, P2, and P3) show an inconsistent range of increases, with P3 reaching the highest value among the treatment groups (10.60 ± 1.14). However, further analysis indicates that only the comparison between K0 and KN is statistically significant (p < 0.05). All other pairs do not show meaningful differences, indicating that coitus recovery remains partial and has not yet reached a level that can be statistically distinguished among the treatment groups.
The libido-based graphical analysis demonstrates consistent intergroup separation for introduction and climbing frequencies, with clear statistical discrimination among groups. The pattern indicates reduced values in diabetic controls and progressive increases across treated groups. Mean ± SD values were well distributed with minimal overlap in most comparisons. Significant differences were primarily observed in the introduction and climbing parameters. Coitus frequency showed greater variability and limited statistical separation. Consequently, only statistically robust parameters are emphasized in the interpretation of this figure.
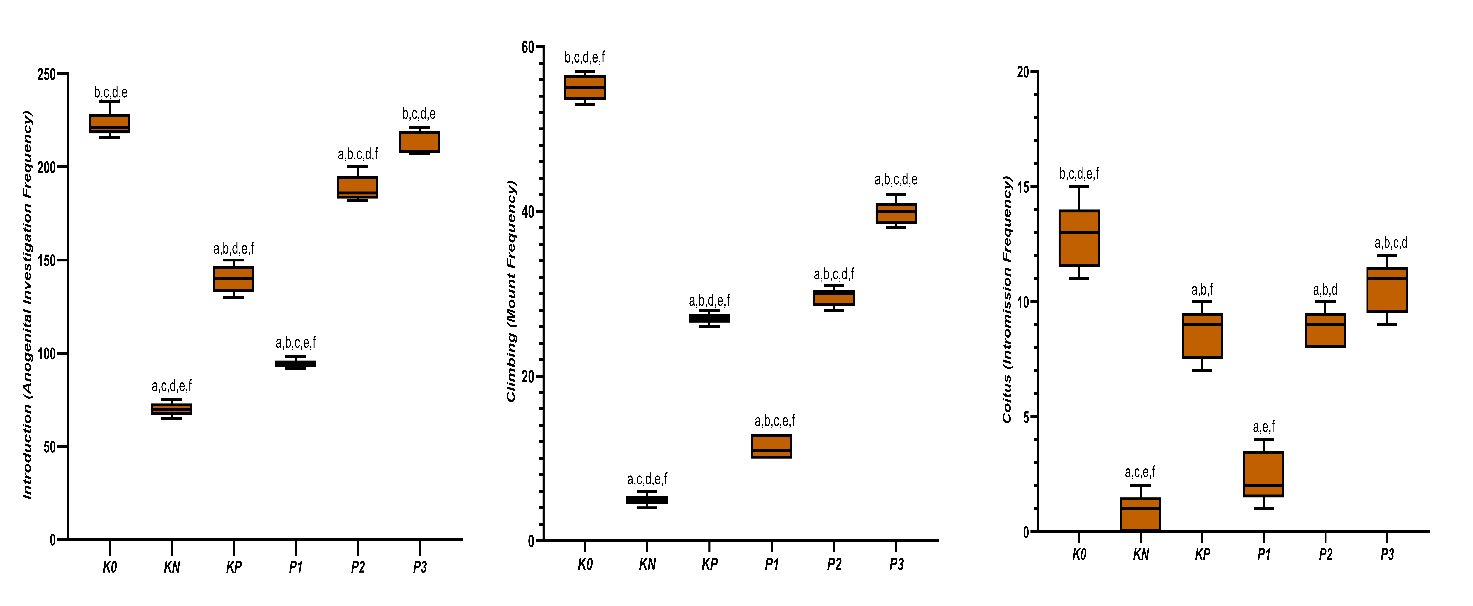
Figure 4 Intergroup comparison of introduction frequency, climbing (mount) frequency, and coitus (intromission) frequency among experimental groups. Data are presented as box-and-whisker plots with mean ± SD (n = 5 per group). Statistical analysis was performed using one-way ANOVA followed by Tukey’s post-hoc test. Different superscript letters indicate significant differences between groups (p < 0.05) (a,b,c,d,e,f= K0, KN, KP, P1, P2, P3) while “ns” denotes non-significant comparisons.
A significant decrease in introduction frequency in diabetic animals reflects disruptions in the central neural substrates of sexual motivation. The medial preoptic area (mPOA) and its dopaminergic inputs are essential for initiating male sexual behavior, as central dopamine release in the mPOA increases during precopulatory encounters and facilitates early motivational sequences such as approach and mounting. Chronic metabolic disturbances such as diabetes impair dopaminergic signaling and diminish sexual motivation, resulting in reduced frequencies of early behavioral components that are more sensitive to neuro-metabolic changes than later consummatory actions [12]. The increased introduction after the administration of CGA indicates a partial recovery in the central neuro-metabolic pathway. CGA is known to be able to cross the blood–brain barrier and reduce neuronal oxidative stress through activation of Nuclear factor erythroid 2-related factor 2 (Nrf2) - antioxidant response element (ARE) signaling and suppression of mitochondrial ROS [43, 44]. A study by Hada et al. [45] reported that CGA increases the expression of Nrf2 and heme oxygenase-1 (HO-1) in brain tissue, thereby protecting hypothalamic neurons from oxidative damage. Additionally, CGA has been reported to enhance neuronal AMPK phosphorylation, which plays a crucial role in energy sensing and motivation-based behaviors. The recovery of this pathway allows the central nervous system to reinitiate basic social behaviors such as introduction. The stronger effects observed at moderate–high doses suggest that the repair of this molecular pathway requires a certain antioxidant threshold. Therefore, the introduction becomes the primary parameter indicating a response to the improvement of internal conditions.
Climbing or mounting is a more complex behavior compared to introduction because it requires sustained motor activation, neuromuscular integrity, and endocrine support. Diabetes reduces climbing ability through a combination of neuromuscular junction (NMJ) dysfunction, decreased mitochondrial capacity in skeletal muscles, and peripheral glucose metabolism disturbances [46]. Shelbayeh et al. [47] research shows that diabetes decreases the expression of PGC-1α and mitochondrial respiratory complexes in muscles, thereby reducing endurance and motor performance. Additionally, peripheral oxidative stress also damages neuromuscular transmission by lowering acetylcholine availability and nicotinic receptor sensitivity [48]. This combination of impairments explains why climbing is more affected than the introduction. In other words, climbing represents a ‘second layer’ of function that depends on the readiness of both central and peripheral systems.
The increase in climbing in the CGA group indicates that this compound not only works in the central system but also restores peripheral function. CGA has been reported to activate AMPK–PGC-1α signaling in muscle tissue, which plays a crucial role in mitochondrial biogenesis and energy efficiency [49]. A study by Gu et al. [50] Showed that CGA increases the expression of PGC-1α and nuclear respiratory factor 1 (NRF-1) in the skeletal muscles of diabetic rats, thereby improving oxidative capacity and muscle endurance. This improvement allows the rats to maintain motor activity for a longer period, reflected in increased climbing frequency. However, because neuromuscular recovery requires time and the tissue’s adaptive capacity, the effects of CGA on climbing appear more slowly and are more dose-dependent compared to the introduction. This explains why climbing does not recover as quickly as the introduction. Therefore, climbing becomes a more ‘biologically costly’ indicator of peripheral function recovery.
The frequency of coitus is consistently lower and more variable compared to introduction and climbing because this behavior represents the most complex endpoint in the reproductive response hierarchy. Coitus requires precise synchronization between central dopaminergic signals, peripheral androgen levels, spinal reflex integrity, and sensorimotor coordination [51]. Unlike introduction and climbing, coitus is highly dependent on testosterone levels that surpass a certain physiological threshold. Hull & Dominguez’s [22] The study emphasizes that even if dopamine and motor activity have recovered, coitus does not occur if androgen stimulation is not optimal. Additionally, coitus is very sensitive to experimental stress and individual variations, so its response is often delayed. Therefore, the low occurrence of coitus in this study is not a failure of intervention but a reflection of more complex biological needs. This parameter is indeed known to be the most difficult to restore.
Although CGA improves the metabolic and oxidative environment, recovery of coitus requires longer hormonal and neural reprogramming. Testosterone, although increased in the CGA group, may not have yet reached or maintained the optimal threshold throughout the behavioral observation period. That recovery of copulatory behavior lags behind improvements in metabolic and motor parameters because it requires stabilization of the HPG axis [52]. Additionally, CGA primarily works through antioxidant and metabolic pathways, rather than as a direct androgen agonist. This causes its effects on coitus to be indirect and gradual. Therefore, coitus is biologically at the ‘end’ of the function recovery cascade. This explanation aligns with the literature on experimental reproductive behavior.
Metformin and CGA show differences in behavioral recovery patterns due to their distinct primary molecular targets. Metformin primarily works through the activation of hepatic AMPK and the reduction of gluconeogenesis, with indirect effects on the nervous system and muscles. In contrast, CGA has multi-target effects: antioxidant, neuroprotective, and mitochondrial [53]. Gao et al. [54] Indicates that metformin improves metabolic parameters without significantly increasing the expression of neuromuscular or neurobehavioral genes. This explains why improvements in introduction and climbing are more limited with metformin. CGA, through the simultaneous modulation of Nrf2, AMPK, and PGC-1α, provides broader systemic benefits. Therefore, the behavioral differences between CGA and metformin are mechanistic in nature, not coincidental.
In the context of behavioral hierarchy, metformin tends to improve initial parameters that depend on glycemic status, while CGA reaches a higher level of function through redox and mitochondrial stabilization. Zhang et al. [55] shows that polyphenol-based interventions result in broader behavioral recovery compared to conventional antidiabetic therapy. These findings are consistent with this study’s results, where moderate to high doses of CGA demonstrate clearer recovery in introduction and climbing behaviors. However, both CGA and metformin show limitations in short-term coitus recovery. This emphasizes that copulatory behavior is an endpoint that requires more time and hormonal adaptation. Therefore, the difference between CGA and metformin lies in the depth of the affected biological pathways, not just metabolic effectiveness.
The sexual behavior response shows different recovery levels among libido components [56]. Climbing activity increases consistently with treatment dose, which may reflect improvements in hormonal function and mid-stage sexual motivation [26,57]. In contrast, coitus exhibits a slower and more variable recovery, so differences between groups are not always significant. This mechanism aligns with the nature of intromission, which requires integrated coordination of motor skills, hormonal stimulation, and the readiness of the female partner, making the recovery effects more complex and gradual.
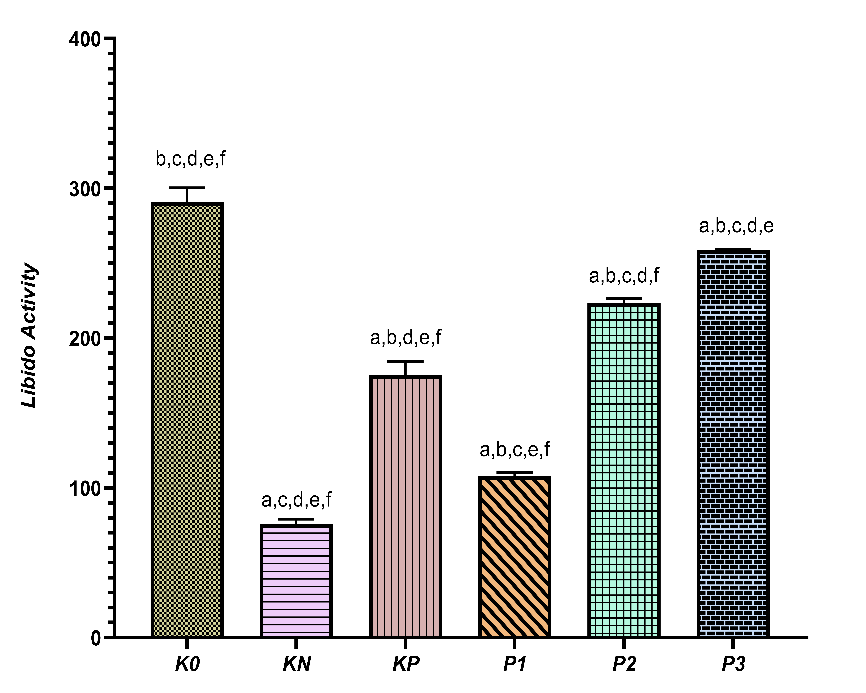
Figure 5 Total libido activity in each treatment group. Data are presented as mean ± standard deviation (SD) for independent experimental groups (n = 5 per group). Statistical analysis was performed using one-way ANOVA followed by Tukey’s test. Different superscript letters (a-f) indicate significant differences between groups (p < 0.05).
Figure 5 shows clear response patterns in libido activity across the groups. The highest value was observed in K0 (290.60 ± 9.84), indicating optimal sexual activity under healthy physiological conditions. KN showed a marked decrease (75.80 ± 3.27), reflecting impaired sexual behavior due to the disease condition. The treatment group exhibited gradual changes. KP showed an increase in sexual behavior activity to 175.60 ± 8.76, while P1 experienced a slight relative decrease compared to KP (108.00 ± 2.35). However, an increase was again observed in P2 (223.20 ± 3.27), and P3 reached a value close to the healthy group (258.80 ± 0.84). The 95% confidence interval range indicates non-overlapping estimates between the healthy and sick groups, with the graded treatment groups falling in between. According to the superscript notation on the graph, all groups differed significantly from each other (p < 0.05), indicating that each treatment produced a level of sexual behavior activity that can be statistically distinguished.
The response pattern indicates that sexual behavior activity is highly sensitive to physiological status and recovery triggered by interventions. The sharp decline in KN reflects libido disturbances closely related to dysfunctional metabolic conditions. The recovery observed in KP and continuing through P3 suggests that the intervention can progressively improve sexual behavior. However, the increase is not always linear, as evidenced by a temporary decrease at P1 before rising again at P2 and P3. The value at P3, which nearly matches K0, indicates that the highest dose of treatment has the potential to restore sexual activity close to normal conditions, consistent with previous findings on libido parameters, especially introduction and climbing. Overall, this combined variable confirms that sexual behavior activity can serve as a comprehensive indicator of libido recovery, as it is sensitive to changes in both pathophysiological conditions and the effects of tiered interventions.
Samples were divided into three categories—low, medium, and high libido—each representing 33.3% based on percentile values. In the DM rat group and the group receiving 6.25 mg/kg chlorogenic acid, all rats fell into the low libido category, suggesting that diabetes reduces libido and that this low dose of chlorogenic acid is insufficient to significantly boost libido. Rats treated with metformin (KP) and those given 12.5 mg/kgBB chlorogenic acid were in the medium libido group, showing improved libido compared to the DM group, yet not reaching the levels seen in healthy rats (K0). This phase indicates a transitional stage where better glycemic control and enhanced antioxidant activity begin to influence testosterone levels but are not yet optimal. The most notable effects appeared in rats receiving 25 mg/kgBB chlorogenic acid, which fell into the high libido category, approaching the values of healthy rats (K0). This demonstrates a progressive increase in libido with higher doses of chlorogenic acid.
The decrease in overall sexual behavior activity in diabetic rats reflects that diabetes mellitus has a broad impact on reproductive behavior expression, not just on specific physiological aspects [58]. Sexual behavior in rats results from the integration of metabolic conditions, hormonal balance, central nervous system function, and overall physical condition. In diabetic conditions, chronic metabolic disturbances and prolonged hyperglycemia are known to reduce motivation and behavioral sexual responses [59]. This aligns with the report by Furman et al. [60] which shows that diabetic rats experience a significant decrease in overall sexual activity due to systemic disturbances affecting behavior regulation. This decline is not merely caused by a single factor but is the result of various internal disruptions. Therefore, total sexual behavior becomes a relevant indicator for assessing the impact of diabetes on reproductive function comprehensively. This parameter reflects the final functional condition experienced by the experimental animals.
The increase in total sexual behavior activity after the administration of CGA indicates an improvement in internal conditions that support the expression of such behavior. Several studies have reported that CGA has antioxidant and metabolic activities that can improve the overall condition of diabetic animals, allowing sexual behavior to gradually recover [61]. A study by Al-Megrin et al. [62] Reported that administering CGA to diabetic rats improved reproductive behavior that had declined due to oxidative stress and metabolic disturbances. The polyphenol content in CGA is known to play a role in reducing oxidative stress and restoring internal balance, which indirectly supports the recovery of sexual behavior. The more pronounced improvements at medium and high doses suggest that the effects of CGA are dose-dependent. Therefore, the increase in total sexual behavior in the CGA group reflects a recovery of a more conducive biological environment for the expression of normal behavior. These results are consistent with previous findings in experimental diabetes models.
Although metformin can improve sexual behavior activity compared to the diabetes group, the level of recovery achieved is relatively lower than that of high-dose CGA. Metformin is known to be effective in controlling blood glucose levels, but its effects on sexual behavior are often indirect. Several studies indicate that improvements in sexual behavior after metformin administration are related to overall metabolic improvements, rather than specific effects on behavior regulation. Tseng et al. [42] Reported that metformin could improve some parameters of sexual behavior in diabetic rats, but full recovery is often not achieved within a short observation period. This suggests that glycemic control alone may not be sufficient to restore sexual behavior optimally. Therefore, the differences in response between CGA and metformin can be explained by differences in the nature of the compounds and their scope of biological effects. CGA tends to provide broader support for internal conditions that influence behavior.
In general, the improvement in total sexual behavior activity after administration of CGA and metformin is related to the ability of these compounds to repair internal conditions that are disrupted due to diabetes. CGA, which is rich in phenolic compounds, plays a role in reducing oxidative stress and improving overall physiological conditions, thereby supporting the recovery of sexual behavior. Meanwhile, metformin primarily works through improving glucose metabolism, which gradually also impacts behavior. A study by Rahman et al. [63] Showed that interventions capable of simultaneously improving metabolic and oxidative conditions yield better results in the sexual behavior of diabetic rats. Therefore, the changes in sexual behavior observed in this study can be understood as a result of improved systemic conditions, rather than direct stimulation of behavior. This approach positions sexual behavior as a reflection of the overall health status of the animal.
Association of HbA1c, MDA, SOD, testosterone, and libido in male diabetic rats given CGA
Table 1 shows strong and highly significant correlations between metabolic parameters, oxidative stress, hormonal status, and sexual behavior in male rats. HbA1c levels exhibit a very strong negative correlation with SOD and testosterone, while showing a strong positive correlation with MDA. In relation to libido activity, HbA1c also demonstrates significant negative associations with the frequency of introduction, climbing, coitus, and overall sexual activity or libido (p < 0.01). These findings indicate that poor glycemic control is closely linked to increased oxidative stress, reduced testosterone levels, and impaired sexual behavior parameters.
Conversely,
SOD activity and testosterone levels show strong positive
correlations with sexual performance.
SOD is positively
associated with introduction, climbing, coitus, and libido (p
< 0.01). Testosterone demonstrates the highest positive
correlations among biological parameters, showing strong
associations with introduction, climbing, coitus and libido (p
< 0.01). Meanwhile, MDA displays a strong negative correlation
with all sexual behavior variables, including libido (p
< 0.01), highlighting the detrimental role of oxidative stress on
libido.
Table 1 Pearson correlation between HbA1c, MDA, SOD, Testosterone, and Libido in diabetic rats given CGA
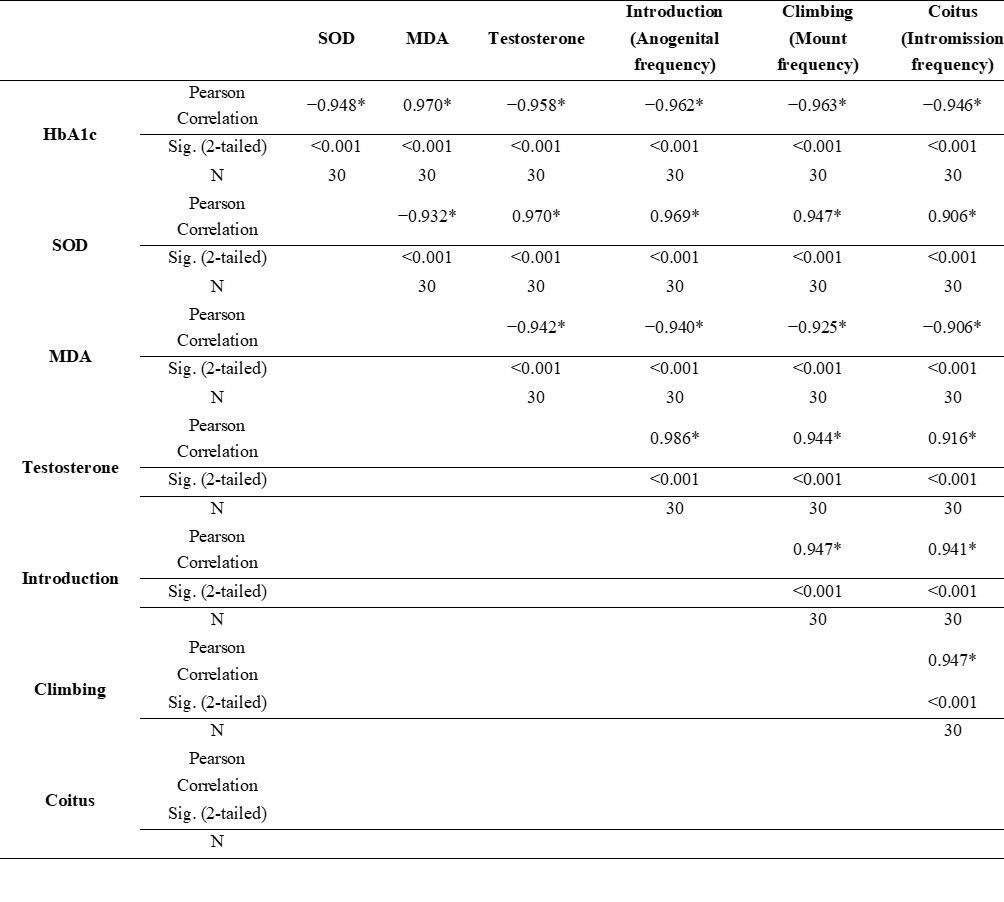
* Correlation is significant at the 0.01 level (2-tailed).
A very strong negative correlation between HbA1c and testosterone, SOD, and all libido components confirms that chronic hyperglycemia is a primary factor in the decline of reproductive function and sexual motivation, likely through oxidative stress mechanisms and endocrine disturbances. Conversely, high SOD is closely correlated with increased testosterone and libido parameters, indicating that redox balance plays an important role in maintaining male sexual function [64]. The strong correlation between testosterone and the total sexual behavior score reinforces the concept that testosterone is the main mediator of libido expression, especially during motivational behavior phases such as introduction and climbing, before transitioning to consummatory behaviors like coitus [65]. Strong correlations between the early motivational components of sexual behavior, particularly introduction and climbing, with overall libido suggest that the initial investigatory phases are the most sensitive indicators of metabolic hormonal disturbances. In contrast, consummatory phases such as coitus show slightly more variability.
Diabetes mellitus in male rats in this study shows effects that are not solely isolated but spread across various interconnected biological systems. The persistent hyperglycemia creates an unstable internal environment and gradually affects physiological functions. These changes are not immediately visible in behavior but develop over time following earlier metabolic and hormonal disturbances [59]. Sexual behavior is a complex output that reflects the overall internal condition of the animal. Therefore, the decrease in sexual activity observed in the diabetic group can be understood as a reflection of prior biological disturbances. This approach helps view behavior not as an isolated variable but as part of a systemic response. Animal models allow these relationships to be observed more clearly. With this framework, all variables analyzed can be positioned within a continuous biological process.
Chronic hyperglycemia, reflected by increased HbA1c, becomes the starting point for various subsequent changes. Long-term glucose exposure causes metabolic stress that impacts sensitive cells and tissues. Improving glycemic control through interventions is an important initial step in the recovery process [66]. Chlorogenic acid and metformin show the ability to lower the glycemic burden, although through different mechanisms. A reduction in HbA1c indicates decreased chronic glucose exposure, which has been a source of metabolic stress. This condition provides space for biological systems to begin readjusting. These glycemic changes do not immediately alter behavior but lay a more stable foundation [67]. This stage becomes a prerequisite for improvements at the next level. This relationship explains why glycemic control is closely related to other observed parameters.
An imbalance between the oxidant and antioxidant status is an almost inseparable consequence of prolonged hyperglycemia. Free radical production increases while the endogenous antioxidant system’s capacity decreases [68]. The reduction in SOD activity reflects a weakening of cellular protective mechanisms. This condition promotes oxidative stress that damages cellular structure and function. The increase in SOD activity after intervention indicates an improvement in cellular defense capacity. CGA is known as a phenolic compound that supports the antioxidant system through various pathways. Metformin also contributes indirectly by improving metabolic conditions [69]. These changes in oxidant status are important in maintaining the integrity of tissues involved in reproductive and nervous functions. The impact is not limited to a single organ. Therefore, the oxidant status becomes an important link between metabolism and behavioral function.
The activation of a low-grade inflammatory response often follows prolonged oxidative stress. This chronic inflammation worsens tissue damage and hampers the healing process. In diabetes conditions, oxidative stress and inflammation reinforce each other. A more controlled inflammatory internal environment follows a reduction in oxidative stress after intervention. CGA has anti-inflammatory properties that support this process. Metformin is also known to suppress inflammation by improving insulin sensitivity [70,71]. An environment with lower levels of inflammation allows tissues to function more efficiently. This is relevant for tissues involved in regulating sexual behavior. The improvement in inflammation does not work in isolation but is integrated with metabolic and oxidative improvements. This relationship demonstrates a close connection between biological systems.
Metabolic disturbances in diabetes also impact mitochondrial function. Mitochondria play a central role in energy supply and oxidative stress regulation [72]. A decrease in mitochondrial efficiency limits the availability of energy for tissues with high demands, such as the brain and muscles. This condition affects physical activity and nerve response. Improvement in mitochondrial function after intervention supports an overall increase in physiological capacity. CGA is reported to play a role in enhancing mitochondrial biogenesis and efficiency [73]. Metformin also influences energy metabolism, although its effects are more dominant in certain tissues. Better energy availability supports activities that require coordination and stamina. These changes occur gradually following tissue adaptation. This stage strengthens the relationship between energy metabolism and behavioral function.
Hormonal regulation, particularly testosterone, plays an important role in male sexual behavior. Diabetes is often associated with decreased testosterone levels due to impaired Leydig cell function [74]. Oxidative stress and inflammation contribute to inhibiting the steroidogenesis process. An increase in testosterone levels after intervention reflects improved endocrine function. CGA shows the ability to support hormonal recovery through improving the internal environment. Metformin also contributes through metabolic improvements that affect the hormonal axis [75]. Testosterone plays a role not only in peripheral tissues but also in the regulation of the central nervous system. This relationship explains the connection between hormonal changes and behavior. However, hormonal changes require time before they are reflected in behavioral responses. This stage serves as a link between biochemical improvements and functional outward responses.
Hyperglycemia and chronic oxidative stress have the potential to cause DNA damage. High oxidative stress increases the risk of changes in genome structure and stability. DNA damage in testicular and nerve tissues can have long-term effects on cell function. Interventions that reduce oxidative stress play a role in decreasing these genotoxic pressures. CGA has protective potential against DNA through free radical scavenging activity. Metformin also shows indirect protective effects by improving metabolic conditions [76,77]. DNA stability is important for maintaining tissue function in the long term. Its impact on behavior is not direct but contributes gradually. This pathway indicates that behavioral changes involve deep molecular levels. The process takes time before resulting in significant functional changes.
Libido and sexual behavior emerge as a result of the integration of the entire system that has been discussed. Disorders in one component can affect the overall expression of behavior. Improvements after intervention are cumulative and mutually supportive [59]. Biochemical and hormonal changes often appear earlier than behavioral changes. This explains why recovery of sexual behavior does not always align with a specific parameter. CGA shows broader effects because it works on several pathways simultaneously. Metformin provides more limited improvements in behavioral aspects. These differences are reflected in the overall pattern of recovery of sexual activity. An integrative approach helps to understand the variation in responses among groups.
Sexual behavior has a different time dynamic compared to biochemical parameters. Recovery of behavior requires a longer adaptation period. Testing over a short time frame may only capture the initial phase of changes. Individual variability is also greater in behavioral parameters. This condition makes the interpretation of results require caution. Several studies indicate that the effects of interventions on libido become more apparent in long-term observations [78]. A broader testing time frame allows for the stabilization of biological systems. This aspect is important for designing further research. A longitudinal approach will provide a more comprehensive picture. Time dynamics are a crucial factor in understanding changes in sexual behavior.
This study has several limitations that should be noted. The duration of the intervention and behavioral observations were relatively short, so they do not fully represent long-term hormonal conditions and sexual behavior, especially during more complex copulation stages. Additionally, the sample size in each group was still limited, as is common in experimental research on animals to balance statistical analysis power with ethical considerations. Libido improvement, particularly copulatory behavior, is known to require more time because it involves hormone stabilization and nervous system adaptation. Therefore, future research with longer intervention periods, gradual behavioral observations, and larger sample sizes is necessary to better understand the long-term effects of chlorogenic acid on male reproductive and sexual functions.
Conclusions
This study shows that diabetes mellitus in male rats diminishes sexual activity due to various interconnected biological disturbances, including poor glycemic control, oxidative–antioxidant imbalance, increased oxidative stress, and reduced testosterone levels. improves glycemic control, restores redox status, and reduces oxidative stress, which aligns with increased testosterone and libido levels, with the restorative effects of chlorogenic acid being broader compared to metformin. These results confirm that sexual behavior, especially libido, is influenced by the complex interaction of multiple biological systems rather than a single factor. Nonetheless, the study has limitations such as a short intervention period, indirect measures of inflammation and DNA damage, and individual variations in sexual responses. Consequently, future research should include longer observation periods and larger sample sizes to conduct more comprehensive libido assessments and molecular analyses like gene and protein expression related to oxidative stress, inflammation, and hormonal regulation to better understand the mechanisms behind sexual behavior changes in diabetic conditions.
Acknowledgements
The author gratefully acknowledges the Faculty of Medicine, Diponegoro University, Indonesia for providing institutional support throughout the planning and implementation of this study. The author further thanks colleagues from the Doctoral Program in Medical and Health Sciences for their valuable discussions and scholarly input, which contributed substantially to the refinement of ideas and development of this work. Although this study did not receive external funding, the collaborative academic environment and sustained institutional support were essential to its completion.
Declaration of Generative AI in Scientific Writing
During the preparation of this manuscript, the authors utilized (Grammarly) to improve English grammar and language. Following the application of this tool/service, the authors thoroughly reviewed and edited the content as necessary and take full responsibility for the final published version.
CRediT Author Statement
Eko Naning Sofyanita: Conceptualization, Methodology, Software, Data curation, Writing - original draft, Project administration, and Funding acquisition. Ari Suwondo: Data curation, Writing - original draft, Formal analysis, Visualization, and Validation. Heri Nugroho: Data curation, Writing - original draft, Formal analysis, Visualization, Investigation, and Supervision. Diana Nur Afifah: Data Curation, Formal analysis, Investigation, and Validation. Suhartono Suhartono: Data Curation, Formal analysis, Investigation, and Validation. Achmad Zulfa Juniarto: Conceptualization; Methodology, Supervision, Validation, and Writing - original draft.
References
[1] EN Sofyanita, A Suwondo, H Nugroho and AZ Juniarto. Review articles open access global research trends on the role of chlorogenic acid on antidiabetic mechanisms and reproductive hormone regulation (2015-2025): A bibliometric analysis. Media Publikasi Promosi Kesehatan Indonesia 2025; 8(12), 1617-1632.
[2] Y Zheng, SH Ley and FB Hu. Global aetiology and epidemiology of type 2 diabetes mellitus and its complications. Nature Reviews Endocrinology 2018; 14(2), 88-98.
[3] B Siudek, O Dróżdż, W Bińczyk, K Lissak, B Nowińska, P Brzozowska and A Wiśniewska. Re-diagnosis of diabetes mellitus type 1 to maturity-onset diabetes of the young type 2 (mody 2) in a 26-year-old male: A case report. Quality in Sport 2024; 18, 53394.
[4] BAR Mesa and WD Cardona-Maya. Effect of spent coffee grounds on sperm biology: In vitro approach. Revista Chilena de Obstetricia y Ginecologia 2020; 85(4), 312-323.
[5] EN Sofyanita, SP Syanubari and R Afriansya. The effect of Robusta coffee brew (Coffea canephora) administration on the number of pancreatic beta cells (experimental study on white rats (Rattus Norvegicus) induced with streptozotocin). Jurnal Analis Laboratorium Medik 2024; 9(1), 7-13.
[6] P Capogrosso, M Albersen, AL Burnett, OO Cakir, F Dehó, LA Morgado, J Mulhall, A Salonia, AF Sarikaya, M Satchi, EC Serefoglu, M Shabbir, I Sokolakis, T Yap and F Castiglione. Erectile dysfunction: Update on clinical management. European Urology 2025; 88(4), 388-399.
[7] R Bebb, A Millar and G Brock. Sexual dysfunction and hypogonadism in men with diabetes. Canadian Journal of Diabetes 2018; 42(1), S228-S233.
[8] T Mostafa and IA Abdel-Hamid. Ejaculatory dysfunction in men with diabetes mellitus. World Journal of Diabetes 2021; 12(7), 954-974.
[9] S Ray and S Pramanik. Reproductive dysfunctions in males with type 2 diabetes mellitus: An updated review. European Medical Journal Diabetes 2020; 8(1), 79-89.
[10] AC Huang, TC Yeh, NC Wu, PH Lin and KY Yeh. Protective effects of epigallocatechin gallate for male sexual dysfunction in streptozotocin-induced diabetic rats. International Journal of Molecular Sciences 2022; 23(17), 9759.
[11] KN Killari, H Polimati, DSNBK Prasanth, G Singh, SP Panda, GS Vedula and VB Tatipamula. Salazinic acid attenuates male sexual dysfunction and testicular oxidative damage in streptozotocin-induced diabetic albino rats. RSC Advances 2023; 13(19), 12991-13005.
[12] M Bialy, W Bogacki-Rychlik, J Przybylski and T Zera. The sexual motivation of male rats as a tool in animal models of human health disorders. Frontiers in Behavioral Neuroscience 2019; 13, 1-10.
[13] G Rastrelli, L Vignozzi and M Maggi. Testosterone therapy: A friend or a foe for the aging men with benign prostatic hyperplasia? Asian Journal of Andrology 2020; 22(3), 233-235.
[14] KV Derkach, AA Bakhtyukov, IV Romanova, II Zorina, LV Bayunova, VM Bondareva, IY Morina, VK Roy and AO Shpakov. The effect of metformin treatment on the basal and gonadotropin-stimulated steroidogenesis in male rats with type 2 diabetes mellitus. Andrologia 2020; 52(11), 1-13.
[15] Y Du, YJ Zhu, YX Zhou, J Ding and JY Liu. Metformin in therapeutic applications in human diseases: Its mechanism of action and clinical study. Molecular Biomedicine 2022; 3(1), 41.
[16] EN Sofyanita, AZ Juniarto, A Suwondo and H Nugroho. The effect of chlorogenic acid content in coffee can reduce malondialdehyde (MDA) and increase testosterone hormone levels in DM conditions. Gema Lingkungan Kesehatan 2025; 23(1), 20-26.
[17] WA Al-megrin, MF El-khadragy, MH Hussein, S Mahgoub, DM Abdel-Mohsen, H Taha, AAA Bakkar, AE Abdel Moneim and HK Amin. Green coffea arabica extract ameliorates testicular injury in high-fat diet/streptozotocin-induced diabetes in rats. Journal of Diabetes Research 2020; 2020, 6762709.
[18] Y Yan, Q Li, L Shen L, K Guo and X Zhou. Chlorogenic acid improves glucose tolerance, lipid metabolism, inflammation and microbiota composition in diabetic db/db mice. Frontiers in Endocrinology 2022; 13, 1042044.
[19] OO Oguntibeju, Y Aboua and P Kachepe. Possible therapeutic effects of vindoline on testicular and epididymal function in diabetes-induced oxidative stress male Wistar rats. Heliyon 2020; 6(4), e03817.
[20] P Arundani, R I’Tishom and B Purwanto. Administration of kebar grass extract (Biophytum petersianum Klotszch) the viability of spermatozoa in mice (Mus musculus) with diabetes melitus. Oceana Biomedicina Journal 2021; 4(1), 26-37.
[21] AZ Simatauw and AJA Unitly. Description of the estrus cycle of Rattus norvegicus rats exposed to cigarette smoke after being treated with ethanol extract of Kebar grass (Biophytum petersianum Klotzsch). Rumphius Pattimura Biological Journal 2019; 1(1), 1-7.
[22] EM Hull and JM Dominguez. Sexual behavior in male rodents. Hormones and Behavior 2007; 52(1), 45-55.
[23] B Wahono. Effects of peaberry coffee on the sexual behavior and the blood testosterone levels of the male mouse (Mus musculus). In: Proceedings of the 3rd International Conference on Research, Implementation and Education of Mathematics and Science Yogyakarta, Yogyakarta, Indonesia. 2016, p. B21-B26.
[24] N Rahmawati. Test of the aphrodisiac effects of five jamu herbal remedies on male rat libido. Badan Penelitian dan Pengembangan Kesehatan 2012; 1(1), 1-35.
[25] N Rahmawati, A Prichatin and K Dewi. Aphrodisiac effect of lunasia amara blanco, centella asiatica and curcuma domestica combination infusion on male rat libido. In: Proceedings of the International Conference: Research and Application on Traditional Complemantary and Alternative Medicine in Health Care, Surakarta, Indonesia. 2012, p. 44-47.
[26] R Rollando, A Ardanareswari, FH Susanto and E Monica. Aphrodisiac effect of bajakah kalalawit stem extract (Uncaria gambir Roxb.) on male wistar rats (Rattus norvegicus). Jurnal Pharmascience 2022; 9(2), 213.
[27] A Yakubu, MA Akanji and AT Oladiji. Male sexual dysfunction and methods used in assessing medicinal plants with aphrodisiac potentials. Pharmacognosy Reviews 2007; 1(1), 49-56.
[28] LM Shulman and MD Spritzer. Changes in the sexual behavior and testosterone levels of male rats. Physiology and Behavior 2014; 133(1), 8-13.
[29] S Jin, C Chang, L Zhang, Y Liu, X Huang and Z Chen. Chlorogenic acid improves late diabetes through adiponectin receptor signaling pathways in db/db mice. PLoS One 2015; 10(4), e0120842.
[30] V Nguyen, EG Taine, D Meng, T Cui and W Tan. Chlorogenic acid: A systematic review on the biological functions, mechanistic actions, and therapeutic potentials. Nutrients 2024; 16(7), 924.
[31] LY Zuñiga, MC Aceves-de Aceves-de la Mora, M González-Ortiz, JL Ramos-Núñez and E Martínez-Abundis. Effect of chlorogenic acid administration on glycemic control, insulin secretion, and insulin sensitivity in patients with impaired glucose tolerance. Journal of Medicinal Food 2018; 21(5), 469-473.
[32] K Zalewska, M Kulawik, J Gierszewska, A Nowak, P Zielinski and B Kowalski. Chlorogenic acid’s role in metabolic health: Mechanisms and therapeutic potential. Nutrients 2025; 17(20), 3303.
[33] GA Soliman, RF Abdel-Rahman, HA Ogaly, MS El-Naggar, AM Hassan and SA El-Azab. Momordica charantia extract protects against diabetes-related spermatogenic dysfunction in male rats: molecular and biochemical study. Molecules 2020; 25(22), 5255.
[34] PP Maske, PS Kumbhar, AG Wali, JI Disouza and M Sharma. Antioxidant, antidiabetic and lipid profiling of Spermadicyton suaveolens in streptozotocin (STZ) induced diabetic rats. Brazilian Journal of Pharmaceutical Sciences 2023; 59, e21820.
[35] RF Alenezi, A Abdelkhalek, G El-Sayed, I Pet, M Ahmadi, ESE Sherbini, D Pușcașiu and AH Arisha. A natural polyphenol, chlorogenic acid, attenuates obesity-related metabolic disorders in male rats via miR-146a-IRAK1-TRAF6 and NRF2-mediated antioxidant pathways. Biomolecules 2025; 15(8), 1086.
[36] RJ Aitken, EG Bromfield and Z Gibb. Oxidative stress and reproductive function: The impact of oxidative stress on reproduction: A focus on gametogenesis and fertilization. Reproduction 2022; 164(6), F79-F94.
[37] L Palomba, C Silvestri and E Silvestri. Editorial: Redox signaling and homeostasis in the control of metabolism: A systemic interplay between central and peripheral effectors. Frontiers in Endocrinology 2024; 15, 1522613.
[38] Y Tang, H Li, Y Zeng, C Yang, R Zhang, A K Lund and M Zhang. Spermidine as a potential protective agents against Poly(I:C)-induced immune response, oxidative stress, apoptosis, and testosterone decrease in yak Leydig cells. International Journal of Molecular Sciences 2025; 26(6), 2753.
[39] I Sari, DM Rizal and RA Syarif. The effect of chlorogenic acid on endoplasmic reticulum stress and steroidogenesis in the testes of diabetic rats: Study of mRNA expressions of GRP78, XBP1s, 3β-HSD, and 17β-HSD. BIO Web of Conferences 2022; 49, 01001.
[40] EN Sofyanita, T Nasihun and ST Zulaikhah. The influence of administering propolis CMCE extract on FSH, LH, and testosterone levels in MSG-induced male Wistar rat. Sains Medika: Jurnal Kedokteran dan Kesehatan 2019; 10(2), 51-56.
[41] Y Yan, X Zhou, K Guo, F Zhou and H Yang. Use of chlorogenic acid against diabetes mellitus and its complications. Journal of Immunology Research 2020; 2020, 9680508.
[42] CH Tseng. The effect of metformin on male reproductive function and prostate: An updated review. World Journal of Men’s Health 2022; 40(1), 11-29.
[43] W Zhang, L Tong, B Jin and D Sun. Diabetic testicular dysfunction and spermatogenesis impairment: Mechanisms and therapeutic prospects. Frontiers in Endocrinology 2025; 16, 1653975.
[44] N Abedpour, MZ Javanmard, M Karimipour and AP Liqvan. Effect of chlorogenic acid on follicular development, hormonal status and biomarkers of oxidative stress in rats with polycystic ovary syndrome. Veterinary Research Forum 2022; 13(4), 513-520.
[45] Y Hada, HA Uchida, N Otaka, Y Onishi, S Okamoto, M Nishiwaki, R Takemoto, H Takeuchi and J Wada. The protective effect of chlorogenic acid on vascular senescence via the Nrf2/HO-1 pathway. International Journal of Molecular Sciences 2020; 21(12), 4527.
[46] S Suresh and S Prakash. Effect of mucuna pruriens (Linn.) on sexual behavior and sperm parameters in streptozotocin‐induced diabetic male rat. The Journal of Sexual Medicine 2012; 9(12), 3066-3078.
[47] OA Shelbayeh, T Arroum, S Morris and KB Busch. PGC-1α Is a master regulator of mitochondrial lifecycle and ROS stress response. Antioxidants 2023, 12(5), 1075.
[48] FA Espinoza-Hernández and A Andrade-Cetto. Chronic antihyperglycemic effect exerted by traditional extracts of three Mexican medicinal plants. Evidence-Based Complementary and Alternative Medicine 2022; 2022, 5970358.
[49] G Vial, D Detaille and B Guigas. Role of mitochondria in the mechanism(s) of action of metformin. Frontiers in Endocrinology 2019; 10, 294.
[50] T Gu, Z Zhang, J Liu, L Chen, Y Tian, W Xu, T Zeng, W Wu and L Lu. Chlorogenic acid alleviates LPS-induced inflammation and oxidative stress by modulating CD36/AMPK/PGC-1α in RAW264.7 macrophages. International Journal of Molecular Sciences 2023; 24(17), 13516.
[51] KI Astuti, AN Putri, R Saputri, IP Sari and TNS Sulaiman. Activity of kersen (Muntingia calabura L.) and kasturi (Mangifera casturi Kosterm.) extract as afrodisiac. Indonesian Journal of Pharmaceutical Science and Technology 2023; 10(2), 75.
[52] MZUH Shah, VK Shrivastava, S Sofi, YF Jamous, MF Khan, FK Alkholifi, W Ahmad and MA Mir. Chlorogenic acid restores ovarian functions in mice with letrozole-induced polycystic ovarian syndrome via modulation of adiponectin receptor. Biomedicines 2023; 11(3), 900.
[53] A Hasanvand. The role of AMPK-dependent pathways in cellular and molecular mechanisms of metformin: A new perspective for treatment and prevention of diseases. Inflammopharmacology 2022; 30(3), 775-788.
[54] S Gao, G Hu, D Li, M Sun and D Mou. Anti-hyperlipidemia effect of sea buckthorn fruit oil extract through the AMPK and Akt signaling pathway in hamsters. Journal of Functional Foods 2020; 66, 103837.
[55] Z Zhang, C Shi and Z Wang. Therapeutic effects and molecular mechanism of chlorogenic acid on polycystic ovarian syndrome: Role of HIF-1α. Nutrients 2023; 15(13), 2833.
[56] A Budiawan. Test of the aphrodisiac activity of watermelon fruit skin infusion (Citrullus lanatus). Pharmed: Journal of Pharmaceutical Science and Medical Research 2021; 4(1), 1-10.
[57] AQAL Sahribulan, I Mawarni, IN Amar and DI Sulaeman. Study of plant extract effects related to libido in male mice (Mus musculus L.). Biotech: Biology and Biology Education Journal 2025; 2(1), 129.
[58] M Kaneedi, Sutyarso, H Busman, CI Kesuma, Y Intan and ML Lande. Ameliorative effect of plant extracts of suruhan (Peperomia pellucida) on blood glucose and libido of male mice injected with alloxan. European Journal of Biomedical and Pharmaceutical Sciences 2019; 6(2), 18-21.
[59] DM Malini, S Wulandari, E Maulani and N Ratningsih. Mating behavior in male rat (Rattus Norvegicus) diabetic that given ethanol extract of jengkol (Archidendron Pauciflorum) fruit peel. Metamorfosa: Journal of Biological Sciences 2020; 7, 133.
[60] BL Furman. Streptozotocin-Induced diabetic models in mice and rats. Current Protocols 2021; 1(4), e78.
[61] GAMP Isvari, E Hadibrata, AE Yunianto and S Susianti. The effects of Piper Nigrum L. (black pepper extract) on libido (sexual behavior) in a male white rat model (Rattus Norvegicus) with diabetes mellitus. 2024; 14(9), 1707-1712.
[62] WA Al-Megrin, MF El-Khadragy, MH Hussein, S Mahgoub, DM Abdel-Mohsen, H Taha, AAA Bakkar, AE Abdel Moneim and HK Amin. Green coffea arabica extract ameliorates testicular injury in high-fat diet/streptozotocin-induced diabetes in rats. Journal of Diabetes Research 2020; 2020, 6762709.
[63] HS Rahman, GA Ahmad, B Mustapha, HA Al-Rawi, RH Hussein, K Amin, HH Othman and R Abdullah. Wet cupping therapy ameliorates pain in patients with hyperlipidemia, hypertension, and diabetes: A controlled clinical study. International Journal of Surgery Open 2020; 26(10), 10-15.
[64] DA Abood, KS Mahdi KS and ZCH Hameed. The correlation study between testosterone levels and Hba1c in type 2 diabetic patients compare with healthy persons. Clinical Medicine and Health Research Journal 2024; 4(1), 765-769.
[65] J Rong, X Leng, K Jiang, J Tan and M Dong. Systemic impacts of diabetes on spermatogenesis and intervention strategies: Multilayered mechanism analysis and cutting-edge therapeutic approaches. Reproductive Biology and Endocrinology 2025; 23(1), 122.
[66] M Sianturi, N Susilaningsih, H Nugroho, N Suci, TN Kristina and M Suryani. Effect of lycopene and metformin combination on phagocytosis, glycemic control, and oxidative stress in rats with type 2 diabetes. Medical Journal of Indonesia 2023; 32(1), 123-128.
[67] FA Espinoza-Hernández and A Andrade-Cetto. Chronic antihyperglycemic effect exerted by traditional extracts of three mexican medicinal plants. Evidence-Based Complementary and Alternative Medicine 2022; 2022, 5970358.
[68] EN Sofyanita and AR Yunianti. Effectiveness of white guava leaves (Psidium Guajava Var. Pyrifera L.) in repair proximal tubule damage and glomerulus diameter in hyperglycemic mice. Journal of Engineering and Technology for Industrial Applications 2023; 9(41), 41-45.
[69] L Hussain, N Aamir, M Hussain, M Asif, Z Chauhdary, F Manzoor, R Siddique and M Riaz. Therapeutic investigation of standardized aqueous methanolic extract of bitter melon (Momordica charantia L.) for its potential against polycystic ovarian syndrome in experimental Animals’ model: In vitro and in vivo studies. Evidence-Based Complementary and Alternative Medicine 2022; 2022, 5143653.
[70] R Hasanuddin, J Jasmiadi and N Abdillah. The analysis of the chlorogenic acid in the ethanol fraction of robusta coffee beans and its effect on glucose levels in wistar rats. Disease Prevention and Public Health Journal 2021; 15(2), 118.
[71] N Apostolova, F Iannantuoni, A Gruevska, J Muntane, M Rocha and VM Victor. Mechanisms of action of metformin in type 2 diabetes: Effects on mitochondria and leukocyte-endothelium interactions. Redox Biology 2020; 34, 101517.
[72] L He. Metformin and systemic metabolism. Trends in Pharmacological Sciences 2020; 41(11), 868-881.
[73] AK Singh and AK Pandey. Alleviation of diabetes Mellitus-Induced reproductive dysfunction by chlorogenic acid in male rats via combating redox imbalance. Indian Journal of Clinical Biochemistry 2025. https://doi.org/10.1007/s12291-025-01301-8
[74] MZ Javanmard, A Zeinali and SG Ahari. Improvement of spermatogenesis and fertility in mice under ionizing radiation using chlorogenic acid. Journal of Isfahan Medical School 2023; 40, 981-990.
[75] MF El-Khadragy, WA Al-Megrin, S Alomar, AF Alkhuriji, DM Metwally, S Mahgoub, HK Amin, OA Habotta, AE Abdel Moneim and RS Albeltagy. Chlorogenic acid abates male reproductive dysfunction in arsenic-exposed mice via attenuation of testicular oxido-inflammatory stress and apoptotic responses. Chemico-Biological Interactions 2021; 333, 109333.
[76] J Santana-Gálvez, L Cisneros-Zevallos and DA Jacobo-Velázquez. Chlorogenic acid: Recent advances on its dual role as a food additive and a nutraceutical against metabolic syndrome. Molecules 2017; 22(3), 358.
[77] C Signorini, R Corsaro, G Collodel, R Maettner, K Sterzik, E Strehler, L Liguori and E Moretti. Addition of chlorogenic acid to human semen: Effects on sperm motility, DNA integrity, oxidative stress, and Nrf2 expression. Antioxidants 2025; 14(4), 382.
[78] H Ahmed and M Aslam. Evaluation of aphrodisiac activity of ethanol extract of Ganoderma lucidum in male Wistar rats. Clinical Phytoscience 2018; 4, 26.