Trends
Sci.
2026; 23(8): 12886
Fermentation Associated Changes in Phenolic Profile, Antioxidant Capacity, and In Vitro Apoptotic Activity of Soursop Leaf Kombucha Against T47D Breast Cancer Cells
Elok Zubaidah1,*, Maryam Tsaqifah Muwahhidah1, Hidayat Sujuti2,
Nur Kusmiyati1, Viera Nuriza Pratiwi3 and Eirene Charista Dea1
1Department of Food Science and Biotechnology, Faculty of Agricultural Technology, Universitas Brawijaya,
East Java 65145, Indonesia
2Department of Biomedical, Faculty of Medicine, Universitas Brawijaya, East Java 65145, Indonesia
3Department of Nutrition, Faculty of Health, Universitas Nahdlatul Ulama Surabaya, Surabaya 60237, Indonesia
(*Corresponding author’s e-mail: [email protected])
Received: 4 December 2025, Revised: 6 February 2026, Accepted: 13 February 2026, Published: 25 March 2026
Abstract
Kombucha fermentation of soursop leaves is associated with changes in functional properties through post-fermentation physicochemical and metabolite profile modifications. This study was evaluated for its effects on physicochemical properties, qualitative LC-MS metabolite profiles, antioxidant capacity, and in vitro anticancer activity against T47D breast cancer cells. Fermentation was carried out for 12 days using leaf concentrations of 0.6% - 2.0%, followed by measurements of pH, total microbial counts, total phenolic and flavonoid contents, antioxidant activity, and LC-MS metabolite profiling. Cytotoxicity and apoptosis induction were assessed using CCK-8 assays, morphological observations, fluorescence imaging (DAPI, Annexin V-FITC/PI), and flow cytometry. At a 1.4% leaf concentration, endpoint comparison between day 0 and day 12 showed an increase in total viable counts from 6.77 to 7.65 log CFU/mL and a marked increase in total phenolic content from 198.13 to 322.03 mg GAE/mL, accompanied by the greatest reduction in antioxidant IC₅₀ values (from 252.29 to 99.73 ppm). Qualitative LC-MS profiling revealed differences in detected compounds between fermented and unfermented samples, including the presence of aglycone flavonoids in fermented kombucha. Acidic soursop kombucha demonstrated the strongest cytotoxicity toward T47D cells (IC₅₀ = 2.35 mg/mL), followed by neutralized kombucha (IC₅₀ = 3.87 mg/mL) and soursop leaf drink (IC₅₀ = 7.37 mg/mL). Flow cytometry indicated a marked shift toward apoptotic populations in fermented samples, with combined early and late apoptosis reaching 83.56% in acidic kombucha treated cells. Kombucha fermentation under acidic conditions was associated with enhanced antioxidant capacity and in vitro anticancer related activity of soursop leaves.
Keywords: Soursop leaf kombucha, Fermentation, Antioxidant activity, Anticancer potential
Introduction
Kombucha is a traditional fermented beverage produced by inoculating sweetened tea with a symbiotic culture of bacteria and yeast (SCOBY). During fermentation, microbial metabolism leads to the production of organic acids, amino acids, and modified polyphenolic compounds that have been associated with changes in the nutritional and functional properties of the beverage [1]. Dominant microbes
Acetobacter, Lactobacillus, and yeasts secrete enzymes like β-glucosidase and tannase, which have been reported to modify complex phytochemicals into smaller or structurally altered forms, rather than directly increasing safety or efficacy. These fermentation associated processes have been linked to enhanced antioxidant capacity and changes in detectable metabolite profiles [2]. Soursop (Annona muricata L.) leaves contain phenolics, flavonoids, alkaloids, terpenoids, and acetogenins with antioxidant, anti inflammatory, and anticancer properties. However, many of these compounds occur in glycosylated or polymerized forms, which may limit solubility, cellular uptake, and biological activity [3]. In addition, certain acetogenins, such as annonacin and squamocin, have been reported to exhibit neurotoxic or cytotoxic effects at high concentrations, raising concerns regarding safety and dosage [4]. Although fermentation has been suggested to alter the bioactivity of plant substrates and may influence their functional profiles, direct evidence supporting toxicity reduction through fermentation remains limited and highly substrate dependent. Microbial enzymatic activity during fermentation has been reported to convert flavonoid glycosides into aglycone forms such as quercetin and kaempferol, which exhibit stronger antioxidant and cytotoxic activities in vitro [5]. Studies on turmeric and ginger kombucha revealed increased phenolic content, antioxidant capacity, and the formation of novel metabolites after fermentation [6]. Collectively, these findings suggest that kombucha fermentation is associated with functional modifications of plant derived compounds rather than guaranteed improvements in safety. Previous studies have reported that fermentation of soursop (Annona muricata) products using lactic acid bacteria and yeasts is associated with increased soluble phenolic content and antioxidant capacity, suggesting improved bioavailability of bioactive compounds. Fermented soursop based products, including yogurt and kombucha, have also shown enhanced sensory and functional attributes, although mechanistic evidence remains limited and highly dependent on fermentation conditions [7,8].
Breast cancer is among the most common malignancies globally, accounting for 16.6% of new cancer cases and over 230,000 deaths annually [9]. Although chemotherapy is a cornerstone of breast cancer treatment, its adverse effects often compromise patient quality of life, highlighting the need for complementary strategies based on bioactive compounds that modulate oxidative stress and apoptosis pathways [10]. Extracts of soursop leaves have demonstrated in vitro cytotoxic activity against breast cancer cell lines, including MCF-7 and T47D cells, primarily through apoptosis induction. Nevertheless, their biological effectiveness may be constrained by limited solubility, stability, and bioavailability of the active constituents. Kombucha fermentation has therefore been proposed as a strategy to modify the physicochemical and functional properties of soursop leaves through fermentation associated microbial processes that may result in altered metabolite profiles. During fermentation, organic acids such as acetic, glucuronic, and lactic acids are produced, which have been linked to redox modulation and apoptosis related signaling in cell based models [11]. However, the relative contribution of acidity versus specific microbial metabolites remains incompletely understood. Accordingly, this study investigated fermentation associated changes in the bioactive composition of soursop leaf kombucha and evaluated their antioxidant capacity and in vitro anticancer related activity against T47D breast cancer cells. By integrating metabolomic profiling with cell based functional assays, this work aims to clarify associations between fermentation driven compositional changes and observed bioactivity, while acknowledging the limitations inherent to in vitro and endpoint based analyses. The findings provide insight into the functional potential of soursop leaf kombucha and support further targeted mechanistic and safety oriented investigations.
Materials and methods
Materials
Fresh soursop (Annona muricata L.) leaves of local varieties were harvested from the third leaf position of young to semi mature shoots at Hijo Farm, Malang, East Java, Indonesia. Harvesting was conducted in the morning between 07:00 and 08:00 a.m. to minimize photodegradation of phenolic compounds. The leaves were washed, air dried, and further dehydrated using a cabinet dryer (220 V, 1200 W) at 70 °C for 3 h. Dried leaves were ground into a fine powder using a stainless-steel chopper and stored in airtight containers at room temperature until use. The plant material was obtained from a local cultivation area and identified based on common vernacular name and morphological characteristics. No formal taxonomic authentication or voucher specimen was available for this study. Commercial black tea (Tong Tji), cane sugar (Gulaku), and a symbiotic culture of bacteria and yeast (SCOBY; Healthy Secret brand) were purchased from local suppliers in Malang, East Java. All reagents used were of analytical grade. The human breast cancer cell line T47D was obtained from the Biomedical Culture Collection, Faculty of Medicine, Universitas Brawijaya, Malang, Indonesia, and maintained according to standard cell culture protocols.
Preparation of soursop leaf kombucha
The soursop leaf kombucha was prepared following a standard protocol with slight modifications. Soursop leaf powder (0.6 - 2.0 %w/v) was packed into tea bags and infused in 500 mL boiling distilled water for 5 min. Subsequently, sucrose (10 %w/v) was added and dissolved, and the infusion was cooled to 25 - 28 °C before inoculation. Fermentations were conducted in sterile 650 mL glass jars. All jars and accessories were sanitized aseptically by hot-water sterilization (rinsed with boiling water prior to use) and handled using clean/aseptic procedures to minimize contamination. For each treatment, 500 mL of sweetened infusion was transferred into the jar, then inoculated with 50 mL (10 %v/v) SCOBY starter culture. Prior to inoculation, the starter culture was homogenized thoroughly to ensure uniform microbial distribution across replicates. Jars were covered with sterile muslin cloth to allow aerobic fermentation and incubated statically for 12 days at room temperature (25 - 28 °C). An unfermented control (soursop leaf drink) was prepared identically but without SCOBY. Each condition was prepared in three independent biological replicates (n = 3). To control batch to batch variation, all treatments were prepared using the same homogenized SCOBY starter batch, identical sucrose concentration, infusion time, inoculum ratio, container type, and incubation conditions. Fermentation was evaluated using an endpoint comparison design, with analyses conducted at day 0 and day 12 to represent the initial and final stages of the fermentation process. This approach was selected to assess compositional and functional differences between unfermented and fully fermented soursop leaf kombucha, rather than to characterize fermentation kinetics or intermediate stage dynamics. For in vitro assays, a neutralized kombucha sample (NSLK) was prepared from the selected optimal formulation (1.4 %w/v) by adding sterile 1 M NaOH dropwise with gentle mixing until the pH reached 6.5 - 7.0. The NSLK sample was included as a comparative treatment to partially assess the contribution of acidity to bioactivity, rather than to fully isolate or model fermentation derived metabolite kinetics. This sample was used for subsequent cytotoxicity and apoptosis analyses.
Fermentation monitoring and chemical analysis
pH was measured using a calibrated digital pH meter (Ohaus Starter 2100, USA). Total microbial counts were determined by the pour plate method on Plate Count Agar (PCA; Merck, Germany) using serial dilutions (10⁻¹ - 10⁻⁶) in 0.85% NaCl, incubated at 30 ± 2 °C for 48 h, and expressed as log CFU/mL. Total phenolic content (TPC) was quantified via the Folin Ciocalteu assay by mixing diluted sample with 10% Folin-Ciocalteu reagent and 7.5% Na₂CO₃, incubating for 30 min, and measuring absorbance at 765 nm. Results were expressed as mg GAE/mL [12]. Total flavonoid content (TFC) was determined using the AlCl₃ colorimetric method, with absorbance measured at 415 nm and results expressed as mg QAE/mL [10]. Antioxidant activity was assessed using the DPPH assay by mixing equal volumes of 0.1 mM DPPH and sample, incubating for 30 min, and reading absorbance at 517 nm. Radical scavenging (%) and IC₅₀ were calculated by nonlinear regression [13].
Identification of bioactive compounds using Liquid Chromatography-Mass Spectrometry (LC-MS)
Bioactive compounds in the selected soursop leaf kombucha and the unfermented infusion were profiled using LC–MS. Sample preparation followed a standardized dilution and clarification workflow. Briefly, each extract sample (powder/paste/liquid) was diluted using a polar solvent appropriate to the sample matrix to obtain a non-saturated solution for MS detection, with a final volume of 1500 µL. The diluted sample was vortex mixed at 2000 rpm for 2 min, followed by spindown/centrifugation at 6000 rpm for 2 min to sediment particulates. The supernatant was collected, filtered through a 0.22 µm syringe filter, and transferred into LC vials. Vials were placed in the autosampler prior to injection. Chromatographic separation was performed on a UPLC system equipped with a C18 column (1.7 µm) maintained at 50 °C. The mobile phase consisted of water with 5 mM ammonium formate (A) and 0.05% formic acid (B), delivered at 0.2 mL/min with a sample temperature of 28.1 °C and a 5 µL injection volume. The gradient program was: 0 - 2 min, 95% A/5% B; 2 - 3 min, 75% A/25% B; 3 - 14 min, 75% A/25% B; 14 - 15 min, 0% A/100% B; 15 - 19 min, 0% A/100% B; 19 - 23 min, 95% A/5% B. Mass analysis was conducted using a Xevo G2-S QTof in positive ESI mode over a 50 - 1300 m/z scan range. Data acquisition and processing were performed using MassLynx (Waters). Metabolite identification (annotation) criteria were based on accurate mass (m/z), isotopic pattern, and retention behavior, followed by database matching against ChemSpider, HMDB, and PubChem.
Cell culture, cytotoxicity, and fluorescence analysis
T47D human breast cancer cell line was obtained from the Biomedical Culture Collection, Universitas Brawijaya. Cells were cultured in RPMI-1640 containing 10% FBS and 1.5% penicillin–streptomycin at 37 °C and 5% CO₂. Cytotoxicity was assessed using the WST-8 (CCK-8) assay. Cells (1×10⁴/well) were seeded in 96-well plates, pre-incubated for 24 h, and treated for 48 h with unfermented soursop leaf drink, acidic kombucha (pH 3.0), or neutralized kombucha (pH 7.0) at 1,500 - 6,000 µg/mL. After treatment, 10 µL CCK-8 reagent was added, incubated for 3 h, and absorbance was measured at 450 nm. Cell viability was normalized to untreated controls, and IC₅₀ values were calculated by nonlinear regression. Morphological changes after 48 h were observed under an inverted microscope (Olympus, Japan). Apoptosis was quantified by flow cytometry (BD Accuri C6, USA) using Annexin V-FITC/PI double staining, with results expressed as the percentage of Annexin V-positive cells. For fluorescence imaging, cells were assigned to four groups: control (K), unfermented drink (MDS, 1.4%), acidic kombucha (KDS, 1.4%, pH 3.0), and neutralized kombucha (NKDS, 1.4%, pH 7.0). After 48 h incubation, cells were stained with DAPI and Annexin V-FITC/PI. DAPI emitted blue fluorescence, while Annexin V and PI emitted red and yellow signals indicating early and late apoptosis. Images were captured using a fluorescence microscope at 40× magnification.
Statistical analysis
Data were analyzed using one-way analysis of variance (ANOVA), followed by Tukey’s HSD post hoc test to identify significant differences among treatments. Statistical significance was set at p < 0.05, and all analyses were conducted using Minitab version 19 (Minitab Inc., USA).
Results and discussion
Total microbial cells of soursop leaf kombucha
Table 1 presents total viable counts from day 0 to day 12 of kombucha fermentation at different soursop leaf concentrations. The largest endpoint increases were observed at 1.4% concentrations. These results indicate that moderate soursop leaf supplementation supported higher overall viable growth under the tested conditions in this endpoint comparison (day 0 vs day 12). Because PCA is non selective, the enumeration represents combined viable populations and does not differentiate yeasts, acetic acid bacteria, or lactic acid bacteria; therefore, group-specific dynamics and microbial interactions were not assessed in this study. Nevertheless, the observed increase in total viable counts is consistent with reports that kombucha fermentations involve cooperative carbohydrate utilization, where sucrose is hydrolyzed and subsequently converted into organic acids and other metabolites by the mixed microbial consortium[14], [15]. Ethanol is subsequently oxidized by acetic acid bacteria (Acetobacter, Komagataeibacter, Gluconobacter) to form acetic acid, while part of the glucose is metabolized into additional organic acids. In parallel, lactic acid bacteria (Lactobacillus, Oenococcus, Leuconostoc), though present in lower abundance, enhance acidity, flavor complexity, and functional properties through the synthesis of lactic acid and secondary bioactive metabolites.
At higher soursop leaf concentrations (1.8% - 2.0%), microbial growth declined markedly (111.25% and 60.50%, respectively), indicating an inhibitory effect at elevated levels. Moderate supplementation (1.2% - 1.4%) supported optimal microbial proliferation by balancing fermentable substrates and bioactive compounds. The reduced growth at higher concentrations likely stems from the antimicrobial activity of phenolics and annonaceous acetogenins, which can inhibit microbial metabolism when present in excess [16]. Similar trends have been reported in other polyphenol-rich fermentations [17]. Compounds such as anonaine, reticuline, caryophyllene, annonacin, and squamocin possess antimicrobial potential and may disrupt microbial homeostasis or induce metabolic stress, thereby suppressing growth and fermentation efficiency [18].
Table 1 Microbial counts of kombucha soursop leaves before and after fermentation.
Soursop leaf concentration (%) |
Total microbial cells (cfu/mL) |
Percentage Increase (%) |
|
Day 0 |
Day 12 |
||
0.6 |
0.19×105 |
0.86×107 |
360.71b |
0.8 |
0.26×105 |
0.96×107 |
264.56bc |
1.0 |
0.34×105 |
1.65×107 |
390.10b |
1.2 |
0.49×105 |
3.54×107 |
627.40a |
1.4 |
0.59×105 |
4.51×107 |
664.97a |
1.6 |
0.71×105 |
1.85×107 |
160.56cd |
1.8 |
0.80×105 |
1.69×107 |
111.25d |
2.0 |
0.90×105 |
1.47×107 |
|
Values represent total viable microbial counts (cfu/mL). Percentage increase was calculated relative to Day 0. Different superscript letters indicate significant differences (p < 0.05).
pH of soursop leaf kombucha
The pH change between day 0 and day 12 reflects acid production by the kombucha microbial consortium. As shown in Figure 1(A), pH decreased from the initial range of 3.8 - 4.6 to 3.0 - 3.3 after 12 days of fermentation. This endpoint decrease is consistent with previous reports in white turmeric and temulawak kombucha, where pH reached around 3.1 - 3.3 after fermentation [19]. To compare treatments more clearly, we evaluated the percentage decrease in pH from day 0 to day 12; the greatest reduction was observed at 1.4% soursop leaf concentration (30.32%), indicating the most pronounced endpoint acidification.
The pH reduction is generally attributed to yeast-mediated sucrose hydrolysis followed by ethanol and CO₂ formation, and subsequent oxidation of ethanol and sugars by acetic acid bacteria into organic acids such as acetic, gluconic, and glucuronic acids [20]. The presence of soursop leaves likely enhances this acidification by providing fermentable substrates and micronutrients that stimulate microbial activity. The pH drop was most evident at moderate soursop leaf concentrations (1.2% - 1.6%), indicating optimal conditions for microbial acid production. At higher concentrations (1.8% - 2.0%), the decline was less pronounced, likely due to inhibitory effects of soursop leaf bioactives. Excess phenolics and acetogenins can suppress lactic and acetic acid bacteria by reducing enzyme activity and hydrogen ion availability [5,21]. The steady pH reduction confirms active fermentation, while variations among treatments highlight how soursop leaf levels influence microbial metabolism and acid formation.
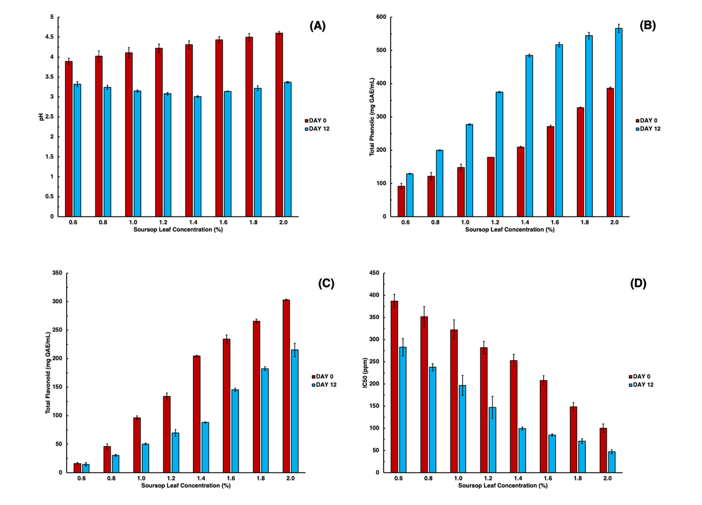
Figure 1 (A) pH values; (B) total phenolic; and (C) total flavonoid; (D) antioxidant activity of kombucha with different concentration of soursop leaves before and after fermentation. Data are presented as mean ± standard deviation (SD) of three independent replicates (n = 3). Error bars indicate SD.
Total phenolic content in soursop leaf kombucha
As shown in Figure 1(B), TPC increased with increasing soursop leaf concentration at both time points (day 0 and day 12), so higher concentrations naturally produced higher absolute TPC values. However, to identify which concentration was most affected by fermentation, we evaluated the percentage increase from day 0 to day 12. The highest relative improvement occurred at 1.4%, with a 62.54% increase (from 198.13 to 322.03 mgGAE/mL), indicating the most efficient phenolic release under the tested conditions. In contrast, although the final TPC continued to rise at higher concentrations, the percentage gains declined (39.54% at 1.6%, 28.93% at 1.8%, and 20.86% at 2.0%), consistent with diminishing returns likely due to reduced mass transfer and extraction efficiency at higher solids loading (increase viscosity and compound aggregation) [22,23]. The observed endpoint increase in TPC is consistent with fermentation associated microbial activity. Although specific enzymatic contributions were not directly measured in this study, bacteria and yeasts commonly associated with kombucha fermentation (e.g., Acetobacter pasteurianus, Zygosaccharomyces bailii, and Debaryomyces hansenii) have been reported to produce enzymes such as glycosidases, cellulases, polyphenol oxidases, and laccases, which may facilitate phenolic release and structural modification. Although microbial activities during fermentation have been associated with polyphenol modifications, the increase in measurable TPC observed here represents an endpoint effect, as individual phenolic transformations were not directly quantified [24]. Overall, while absolute TPC increases with leaf concentration, the percentage increase highlights 1.4% as the optimal concentration for maximizing fermentation associated enhancement of phenolic content in this study.
Total flavonoid content in soursop leaf kombucha
As shown in Figure 1(C), total flavonoid content (TFC) increased with higher soursop leaf concentrations at day 0 but was lower at day 12 compared with day 0 across treatments. Initially, TFC ranged from 16.22 - 302.96 mgQE/mL, while after 12 days values decreased to 14.56 - 215.17 mgQE/mL. To compare fermentation-associated changes among concentrations, we evaluated the percentage decrease from day 0 to day 12. The highest decrease at 1.4% (56.9%), indicating the most pronounced endpoint decline in the colorimetric TFC response under the tested conditions. This reduction may reflect multiple processes, including oxidation/degradation, polymerization, and/or fermentation associated shifts in flavonoid forms that respond differently in the colorimetric assay. Kombucha associated microorganisms have been reported to produce glycosidases and other hydrolases that may promote deglycosylation and generate aglycones (e.g., quercetin and kaempferol); however, these transformations were not directly quantified in this study. Notably, qualitative LC-MS detected aglycones such as quercetin and kaempferol in the fermented sample, which is consistent with a possible profile shift; however, glycoside to aglycone conversion versus degradation cannot be confirmed without targeted quantitative analysis of the corresponding glycosides/aglycones. Despite the overall decrease, final TFC remained relatively high at 1.6% - 2.0%, indicating that higher substrate loading retained a substantial flavonoid pool at the endpoint. Similar decreases in total flavonoids after kombucha fermentation have been reported, for example in white turmeric kombucha, dropping from 85.13 to 56.52 mg QE/mL after 8 days of fermentation [19]. Although total flavonoid levels declined, kombucha fermentation is widely reported to reshape flavonoid profiles through SCOBY driven processes that may influence the functional potential of specific metabolites [25]. The resulting bioactive composition is influenced by substrate concentration and the microbial consortium [26]. Soursop leaves, rich in flavonoids and acetogenins, represent a valuable substrate for the development of functional beverages [27]. For example, β-glucosidase–producing fermentative microbes (e.g., Lactobacillus plantarum in other fermentation systems) have been reported to promote aglycone formation and enhance antioxidant-related propertie [28].
Antioxidant activity of soursop leaf kombucha
As shown in Figure 1(D), antioxidant activity increased after fermentation, as indicated by lower IC₅₀ values on day 12 compared with day 0 across all soursop leaf concentrations. IC₅₀ decreased from 387.05 - 100.40 ppm (day 0) to 283.10 - 47.11 ppm (day 12). To compare fermentation-associated improvements among concentrations, we evaluated the percentage reduction in IC₅₀ from day 0 to day 12, and the greatest decrease was observed at 1.4%, showing a 60.45% reduction, indicating the most pronounced endpoint enhancement in antioxidant activity under the tested conditions.This improvement is consistent with compositional changes occurring as a result of fermentation in the phytochemical profile of soursop leaves, particularly phenolics and flavonoids, which may be converted into smaller or more reactive antioxidant active metabolites. Fermentation by SCOBY members has been reported to promote the release/formation of phenolic forms, including aglycones such as quercetin and kaempferol, which are known for strong radical-scavenging activity. The concurrent increase in measurable total phenolics and decrease in IC₅₀ suggests a relationship between phenolic enrichment and antioxidant capacity in this system, in line with previous reports on polyphenol-rich fermentations [29]. Microbial metabolism within the SCOBY has been reported to facilitate the release and modification of bioactive compounds, potentially generating secondary metabolites with stronger antioxidant-related properties [28]. In addition, substrate characteristics (e.g., phenolic richness and available sugars) can influence the extent of these changes and may contribute to higher antioxidant activity after fermentation [30]. Such effects are commonly attributed to microbial enzymes (e.g., β-glucosidases and other hydrolases) that can hydrolyze complex phenolic conjugates into simpler forms that may exhibit higher radical-scavenging capacity [20]. For example, β-glucosidase activity in fermented systems has been shown to promote aglycone formation from glycosylated flavonoids (e.g., rutin-related conversions), which can enhance antioxidant potential [31]. Previous studies have reported increased antioxidant potential in other kombucha matrices (e.g., turmeric kombucha) following fermentation, which has been associated with changes in phenolic composition [5,20].
Correlation between total flavonoid content, total phenolic content, and antioxidant activity
Pearson correlation analysis demonstrated strong associations among TFC, TPC, and antioxidant activity (Table 2). TFC showed a very strong positive correlation with TPC (r = 0.915, p < 0.01), indicating that flavonoids constitute a substantial component of the phenolic fraction and tend to vary concurrently within the samples. Both TFC and TPC exhibited strong negative correlations with antioxidant IC₅₀ values (r = −0.911 and r = −0.986, respectively; p < 0.01), consistent with the inverse relationship between phenolic content and IC₅₀, where lower IC₅₀ reflects higher antioxidant potency. This pattern aligns with previous reports attributing radical-scavenging capacity to phenolic hydroxyl groups involved in electron or hydrogen donation [32]. The stronger association observed between antioxidant activity and TPC, relative to TFC, suggests that non flavonoid phenolics may contribute substantially to the antioxidant response in this matrix, as also reported in studies on fruit vinegar and fermented plant extracts [33]. These correlations describe statistical associations and should not be interpreted as predictive or causal relationships.
Table 2 Pearson correlation matrix among TFC, TPC, and Antioxidant activity.
Variables |
TFC |
TPC |
Antioxidant activity |
TFC |
1 |
0.915** |
−0.911** |
TPC |
0.915** |
1 |
−0.986** |
Antioxidant activity |
−0.911** |
−0.986** |
1 |
Data were obtained from triplicate analyses (n = 3). Values indicate Pearson’s correlation coefficients (r). **Correlation is significant at p < 0.01 (two-tailed). Antioxidant activity is expressed as IC₅₀; therefore, negative correlations indicate stronger antioxidant capacity.
Comparative chemical and bioactive properties of soursop leaf drink and soursop leaf kombucha
Figure 2 shows a significant difference between soursop leaf drink (SLD) and soursop leaf kombucha (SLK) at a concentration of 1.4%, which contained total microorganisms of 4.51×10⁷ CFU/mL, a pH of 3.01, titratable acidity of 0.81%, total phenolic content of 484.58 mg GAE/mL, total flavonoid content of 88.10 mg QAE/mL, and antioxidant activity with an IC₅₀ value of 99.73 ppm. Compared with its unfermented equivalent, soursop leaf kombucha exhibited decreased pH and higher levels of microbial count, acidity, phenolic and flavonoid contents, and antioxidant activity. The formulation containing 1.4% soursop leaves produced the most pronounced effects, with maximum microbial growth, acidity, phenolic content, and antioxidant strength. Thus, 1.4% soursop leaf supplementation can be regarded as the optimal concentration for kombucha fermentation.
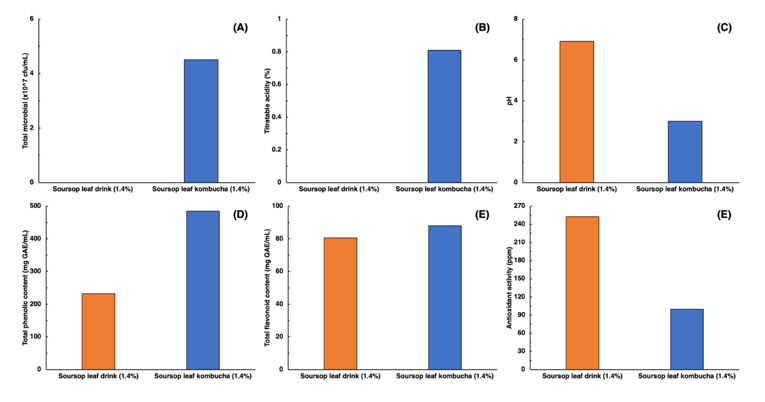
Figure 2 Comparative chemical and bioactive properties of soursop leaf drink and soursop leaf kombucha (1.4% concentration).
Fermentation of soursop (Annona muricata L.) leaves into kombucha enhances antioxidant capacity through synergistic interactions between plant derived bioactives and microbial metabolism within the SCOBY. Enzymes such as β-glucosidase and phenol oxidase, produced by lactic acid bacteria and yeasts, catalyze the release and structural modification of phenolic compounds into more bioactive forms. This enzymatic activities have been reported to be associated with changes in phenolic profiles, which may contribute to the observed increases in total phenolic and flavonoid contents, which contribute to the elevated antioxidant activity of the kombucha [34-36]. Enhancement of phenolic and flavonoid concentrations with fermentation progression underscores the close relationship between fermentation duration and the transformation of bioactive compounds [11,37,38]. Moreover, the rich phytochemical composition of soursop leaves, especially their antioxidant constituents, may act synergistically with microbial metabolites to produce a beverage with superior functional attributes and health-promoting potential [38,39].
Metabolite profiling of soursop leaf kombucha
LC-MS analysis provided a qualitative overview (presence/absence) of compounds detected in soursop leaf drink (SLD) and soursop leaf kombucha (SLK), including alkaloids, flavonoids, terpenoids, phenylpropanoids, amino acids, fatty acids, steroids, saponins, and aromatic compounds (Supplementary 1). Qualitative LC–MS profiling revealed differences in the detectable metabolite composition between soursop leaf drink (SLD) and soursop leaf kombucha (SLK), with a broader range of compound classes observed in fermented samples. Notably, aglycone flavonoids such as quercetin and kaempferol were detected in SLK but not in SLD. The presence of these features is consistent with fermentation-associated shifts in flavonoid profiles reported in previous studies; however, given the qualitative and endpoint nature of the analysis, definitive identification, quantification, or confirmation of glycoside to aglycone conversion cannot be established. Previous studies have reported that microbial activity during fermentation can influence flavonoid profiles through enzymatic and matrix mediated processes, including the potential release of aglycone-related features; these reports are provided here as contextual background rather than evidence of specific transformation pathways occurring in the present system [40,41]. In addition, SLK exhibited detectable features tentatively associated with fermentation-derived metabolites, including organic acid related signals, which have been discussed in the literature in relation to biological responses in fermented food systems. Because LC-MS analysis in this study was qualitative, these observations should be interpreted as compositional differences observed at the final fermentation stage rather than as evidence of increased metabolite abundance or specific enzymatic transformation pathways [42]. Similar fermentation associated modifications of detectable metabolite profiles have been reported in other kombucha substrates, including white turmeric and temulawak kombucha [5,19,43].
Cytotoxic effects of soursop leaf kombucha on T47D breast cancer cells
Cytotoxicity assay revealed a significant reduction in T47D cell viability across all treatments compared with the control (p < 0.05) (Table 3). A clear dose dependent decrease in cell viability was observed with increasing sample concentrations (1500 - 6000 µg/mL). Among the treatments, acidic soursop leaf kombucha showed the strongest cytotoxic effect, reducing cell viability to 13.32 ± 0.01% at 6000 µg/mL, followed by neutralized kombucha (47.61 ± 0.03%) and unfermented soursop leaf drink (61.56 ± 0.02%). Correspondingly, the IC₅₀ values were 2.35, 5.42, and 7.33 mg/mL, respectively, indicating that acidic kombucha exerted the highest cytotoxic potency. The increased cytotoxicity of kombucha is likely associated with fermentation related microbial processes that modify phenolic profiles, including the appearance of aglycone forms such as quercetin, kaempferol, and coumarin related compounds, which have been reported to induce apoptosis and suppress cancer cell proliferation through mitochondrial pathways and Bcl-2 downregulation [44]. Acidic kombucha exhibited higher efficacy than its neutralized counterpart, highlighting the crucial role of an acidic fermentation environment in maintaining the stability and enhancing the functionality of bioactive compounds, particularly flavonoid aglycones. During fermentation, β-glucosidase activity from lactic acid bacteria cleaves glycosidic bonds in polyphenolic compounds, releasing aglycones with improved bioavailability and stronger biological activity [45]. These results show that fermentation is associated with changes in antioxidant capacity and in vitro cytotoxic responses of soursop leaf beverages. Acidic soursop leaf kombucha demonstrates in vitro bioactivity that supports further mechanistic and in vivo investigation. Despite the enhanced cytotoxicity observed, the potential neurotoxicity associated with annonaceous acetogenins warrants careful safety evaluation in future in vivo and long-term studies.
Table 3 Viability of T47D cancer cells in all treatments.
Doses (mg/mL) |
Soursop leaf drink |
Soursop leaf kombucha |
Neutralized soursop leaf kombucha |
1.5 |
98.61 ± 0.05a |
59.51 ± 0.08b |
64.38 ± 0.04b |
3 |
85.28 ± 0.03a |
42.32 ± 0.03c |
51.88 ± 0.00b |
6 |
61.54 ± 0.02a |
13.32 ± 0.01c |
39.57 ± 0.03b |
IC50 (mg/mL) |
7.37 |
2.35 |
3.87 |
Data are presented as mean ± SD (n = 3). Within each dose (row), means with different superscript letters are significantly different among treatments (one-way ANOVA followed by Tukey’s HSD, p < 0.05).
Morphological alterations of T47D cells after treatment
Morphological observations revealed distinct structural changes among treatment groups after 48 h incubation (Figure 3). In the control group (A), T47D cells maintained their normal polygonal morphology with high confluency and intact cell adhesion, indicating healthy proliferating cells. Cells treated with the unfermented soursop leaf drink (SLD) (B) exhibited mild cytotoxic effects, showing slight cell rounding and detachment from the substrate, while most cells retained their regular shape and density. In contrast, cells exposed to acidic soursop leaf kombucha (SLK) (C) displayed the most pronounced morphological alterations, including cytoplasmic shrinkage, rounding, and detachment, accompanied by a marked reduction in cell density. These features are characteristic of apoptotic cell death, reflecting membrane integrity loss and cellular fragmentation. Neutralized soursop leaf kombucha (NKDS) group (D) showed moderate morphological changes, with an increased number of rounded cells compared to the SLD and control, but less severe than SLK. These findings are consistent with the cytotoxicity results (Table 3), where SLK exhibited the lowest IC₅₀ value, confirming its strongest antiproliferative activity. The observed morphological damage may reflect fermentation associated changes in phenolic and flavonoid composition; however, the involvement of specific aglycones such as quercetin, kaempferol, and isocoumarin in apoptosis induction was inferred from the literature rather than directly demonstrated in this study [46]. Fermentation process in kombucha production substantially increases the bioavailability of phenolic compounds, which are known for their potent antioxidant and anticancer properties [47,48]. These results suggest that fermentation significantly enhances the chemopreventive potential of soursop leaf kombucha by inducing morphological and apoptotic changes in cancer cells.

Figure 3 Morphological observation of T47D breast cancer cells after 48 h treatment (10× magnification). (A) Control (untreated cells showing normal polygonal morphology and high cell density); (B) Soursop leaf drink (SLD) showing low cytotoxicity with few detached or rounded cells; (C) Acidic soursop leaf kombucha (SLK) exhibiting marked cytotoxicity with increased cell rounding, shrinkage, and detachment; and (D) Neutralized soursop leaf kombucha (NSLK) showing moderate cytotoxicity with visible morphological alterations compared to SLD and control.
Apoptotic analysis of T47D cells by fluorescent staining
Fluorescent staining using DAPI, Annexin V, and propidium iodide (PI) was conducted to visualize nuclear alterations and apoptotic progression in T47D breast cancer cells after 48 h of treatment (Figure 4). Control cells (K) exhibited uniform blue DAPI fluorescence with intact nuclear morphology and no detectable Annexin V or PI signals, indicating minimal basal apoptosis. Cells treated with the unfermented soursop leaf drink (SLD) showed weak Annexin V and PI fluorescence, suggesting limited induction of early apoptotic events in a small proportion of cells, which may be associated with naturally occurring bioactive constituents present prior to fermentation. In contrast, treatment with acidic soursop leaf kombucha (SLK) resulted in pronounced PI and overlapping Annexin V/PI fluorescence, accompanied by nuclear condensation and fragmentation, indicative of advanced apoptosis and late-stage cell death. The neutralized soursop leaf kombucha (NSLK) group exhibited an intermediate response, with moderate PI fluorescence that was lower than SLK but higher than SLD. This pattern indicates that fermentation conditions, including acidity, may influence the magnitude of cytotoxic and apoptotic responses. Previous studies have reported that acidic fermentation environments and fermentation-associated organic acids can modulate the stability and bioactivity of phenolic compounds, including flavonoids [49,50]. Collectively, these findings underscore that the acidic fermentation environment is fundamental to enhancing the stability, bioavailability, and overall functionality of flavonoid aglycones in kombucha. However, the contribution of acidity to the observed cytotoxic response cannot be fully separated from fermentation associated compositional changes. Although a neutralized kombucha sample was included, the absence of a pH matched vehicle control limits isolation of the direct effect of low pH on T47D cell viability. Accordingly, the apoptotic response observed in this study should be interpreted as an in vitro outcome influenced by both acidity and overall fermentation related compositional changes, rather than attributed to the action of specific phytochemicals alone.
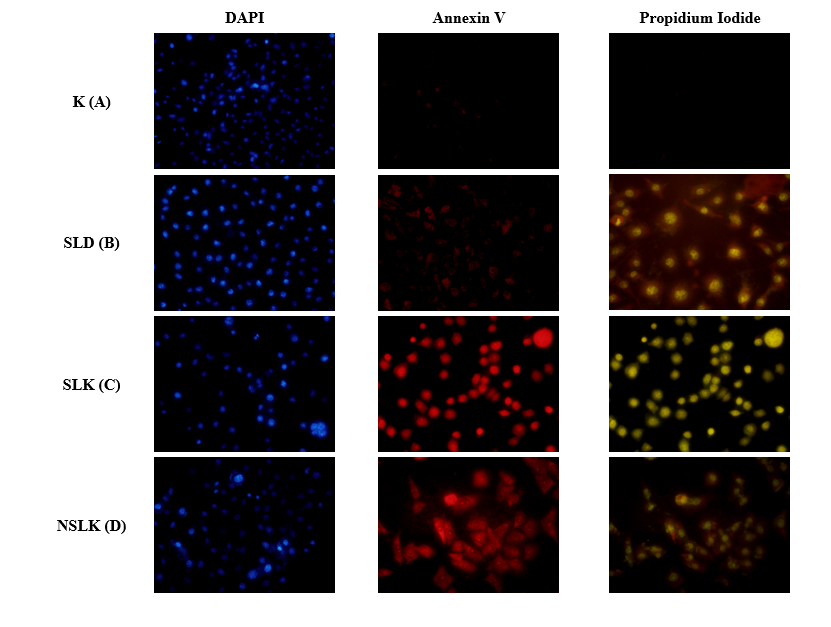
Figure 4 Fluorescent Staining of T47D cancer cells after treatment (40× magnification). (A) negative control (T47D cancer cells) (B) soursop leaf drink (SLD) (C) soursop leaf kombucha (SLK) (D) neutralized soursop leaf kombucha (NSLK). DAPI (4′,6-diamidino-2-phenylindole) is stained blue (nuclear staining); Annexin V-FITC is stained green (early apoptosis); Propidium iodide is stained red (late apoptosis/necrosis).
Apoptosis quantification by flow cytometry
Apoptosis in T47D breast cancer cells was evaluated using Annexin V-FITC/PI dual staining (Figure 5). The control group was predominantly composed of Annexin V⁻/PI⁻ cells, indicating minimal basal apoptosis. Treatment with the unfermented soursop leaf drink (SLD) induced a clear shift toward apoptotic populations, with increases in both early (Annexin V⁺/PI⁻) and late apoptosis (Annexin V⁺/PI⁺). This response is consistent with previous reports describing the pro-apoptotic activity of acetogenins and flavonoids present in soursop leaves, which have been associated with mitochondrial dysfunction, reactive oxygen species generation, and caspase activation [51]. In contrast, treatment with soursop leaf kombucha (SLK) resulted in a more moderate apoptotic response, characterized mainly by the accumulation of cells in the early apoptotic quadrant. This pattern suggests that fermentation associated compositional changes may preferentially promote apoptosis initiation without rapid progression to later stages. Fermentation has been reported to alter polyphenolic profiles, increase organic acid content, and generate additional microbial metabolites that can modulate cytotoxic intensity [52], which may contribute to the predominance of early apoptotic cells observed following SLK treatment. The neutralized soursop leaf kombucha (NSLK) exhibited the most pronounced pro-apoptotic profile, with a substantial proportion of cells located in the Annexin V⁺/PI⁺ quadrant, along with increased PI-positive populations indicative of late apoptosis and secondary necrosis. This response may reflect combined effects of soursop leaf constituents and fermentation-derived components under neutralized conditions. Previous studies have suggested that fermentation associated polyphenols and organic acids can influence apoptotic signaling pathways, including mitochondrial permeabilization, caspase-3 activation, and modulation of PI3K/Akt survival signaling [53,54]. Flow cytometry analysis demonstrated increased early and late apoptotic populations in T47D cells treated with fermented soursop leaf preparations compared with the unfermented control. However, this apoptotic response should be interpreted as being associated with overall fermentation related compositional changes rather than attributed to increased concentrations of specific metabolites, given the qualitative nature of the metabolite analysis.
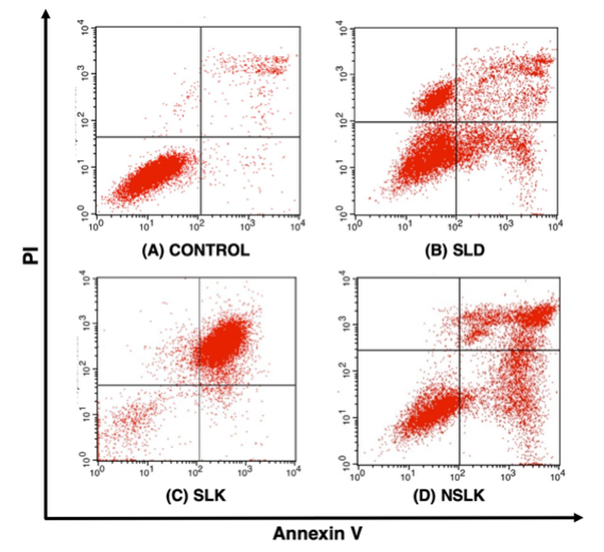
Figure 5 Apoptosis of T47D cancer cells using flow cytometry.
Table 4 Flow cytometry on T47D cells.
Quadrant |
Cell control (untreated) |
Soursop Leaf Drink (SLD) |
Soursop Leaf Kombucha (SLK) |
Neutralized soursop leaf kombucha (NSLK) |
Live cells (%) |
93.12 |
45.95 |
7.04 |
43.26 |
Early apoptosis(%) |
1.09 |
19.19 |
6.36 |
25.86 |
Late apoptosis (%) |
5.00 |
17.37 |
83.56 |
29.55 |
Necrosis (%) |
0.60 |
17.48 |
2.37 |
1.00 |
Data are presented as percentages of cell populations determined by Annexin V-FITC/PI staining and flow cytometry. Quadrant definitions were: live cells (Annexin V⁻/PI⁻), early apoptosis (Annexin V⁺/PI⁻), late apoptosis (Annexin V⁺/PI⁺), and necrosis (Annexin V⁻/PI⁺).
Limitations of the study
This study has several limitations that should be considered when interpreting the findings. Fermentation was assessed using an endpoint comparison design, which captures differences between the initial and final stages but does not resolve intermediate microbial or chemical dynamics. Metabolite profiling by LC-MS was qualitative, providing information on detectable features rather than absolute concentrations or relative abundances, thereby limiting quantitative interpretation of individual compounds. In addition, the contribution of acidity to the observed cytotoxic and apoptotic responses could not be fully isolated, as a pH matched vehicle control was not included. Anticancer related activity was evaluated using a single hormone dependent breast cancer cell line, and selectivity toward non cancerous cells was not assessed. Safety related endpoints, including targeted quantification of potentially neurotoxic acetogenins and toxicity evaluation, were not examined. These constraints indicate that the present results represent preliminary in vitro observations rather than definitive evidence of efficacy or safety.
Conclusions
This study shows that fermentation of soursop leaves into kombucha is associated with changes in physicochemical properties and enhanced in vitro bioactivity compared with the unfermented leaf drink. The fermented product exhibited higher total phenolic and flavonoid contents together with lower antioxidant IC₅₀ values, indicating improved radical scavenging capacity. In cell based assays, acidic soursop leaf kombucha demonstrated the strongest antiproliferative and apoptosis associated responses against T47D breast cancer cells. Overall, these findings suggest that kombucha fermentation is associated with compositional differences that coincide with enhanced antioxidant and apoptosis related in vitro responses. However, given the qualitative metabolite analysis, endpoint fermentation design, and absence of targeted safety assessment, the results should be interpreted as exploratory. Further studies employing quantitative metabolomics, pH controlled experimental designs, multiple cell models, and in vivo safety and efficacy evaluation are required before any health related or chemopreventive applications can be substantiated.
Acknowledgements
This research was supported by the Professor Grant Program of Universitas Brawijaya, Indonesia, under Grant No. 01218/UN/F1001/B/PT0103.2/2024. The authors also express their gratitude to the Faculty of Agricultural Technology for laboratory facilities and technical assistance provided throughout the study.
Declaration of Generative AI in Scientific Writing
The authors declare that generative artificial intelligence (AI) tools were used to assist in language refinement and structural editing of this manuscript. All content, including the scientific concepts, data interpretation, and conclusions, was developed, verified, and approved by the authors. The authors take full responsibility for the integrity and accuracy of the manuscript.
CRediT Author Statement
Elok
Zubaidah:
Conceptualization; Supervision; Resources; Funding Acquisition;
Writing-Review & Editing.
Maryam
Tsaqifah Muwahhidah:
Methodology; Investigation; Project Administration; Formal Analysis;
Data Curation; Visualization; Writing-Original Draft. Hidayat
Sujuti:
Supervision; Validation; Writing-Review & Editing. Nur
Kusmiyati:
Supervision; Validation; Writing-Review & Editing. Viera
Nu’riza Pratiwi:
Formal Analysis; Data Curation; Visualization; Writing-Original
Draft. Eirene
Charista Dea:
Formal Analysis; Data Curation; Visualization; Writing-Original
Draft.
References
[1] M Praveen and S Brogi. Microbial fermentation in food and beverage industries: Innovations, challenges, and opportunities. Foods 2025; 14(1), 114.
[2] RH Abdallah, R Al-Sayed Al-Attar, YM Shehata, DM Abdel-Fattah, RM Atta, OI Fantoukh and AM Mustafa. Comprehensive chemical profiling and mechanistic insight into anticancer activity of Annona muricata leaves extract. Pharmaceuticals 2024; 17(5), 614.
[3] MJ Rendón-Barrón, E Pérez-Arteaga, I Delgado-Waldo, J Coronel-Hernández, C Pérez-Plasencia, F Rodríguez-Izquierdo, R Linares, AR González-Esquinca, I Álvarez-González and NJ Jacobo-Herrera. Laherradurin inhibits tumor growth in an azoxymethane/dextran sulfate sodium colorectal cancer model in vivo. Cancers 2024; 16(3), 573.
[4] IS Sharafutdinov, GD Ozhegov, AE Sabirova, VV Novikova, SA Lisovskaya, AM Khabibrakhmanova, AR Kurbangalieva, MI Bogachev, AR Kayumov. Increasing susceptibility of drug-resistant candida albicans to fluconazole and terbinafine by 2(5H)-furanone derivative. Molecules 2020; 25(3), 642.
[5] E Zubaidah, EC Dea and H Sujuti. Physicochemical and microbiological characteristics of kombucha based on various concentration of Javanese turmeric (Curcuma xanthorrhiza). Biocatalysis and Agricultural Biotechnology 2022; 44, 102467.
[6] SF Tan, NL Chin, TP Tee and SK Chooi. Physico-chemical changes, microbiological properties, and storage shelf life of cow and goat milk from industrial high-pressure processing. Processes 2020; 8(6), 697.
[7] NTT Hạnh, NN Cham, NT Trang and CTN Anh. Effect of low temperature storage period on some quality attributes of soursop (Annona Muricata L.) juice fermented using lactobacillus plantarum and saccharomyces bayanus. Current Research in Nutrition and Food Science Journal 2025; 13(1), 243-257.
[8] J Nanyondo, S Byakika and IM Mukisa. Production of a probiotic Soy-Soursop yogurt containing lactobacillus rhamnosus yoba 2012. European Journal of Agriculture and Food Sciences 2023; 5(4), 60-64.
[9] F Bray, M Laversanne, H Sung, J Ferlay, RL Siegel, I Soerjomataram and A Jemal. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA: A Cancer Journal for Clinicians 2024; 74(3), 229-263.
[10] Herwin, Fitriana and S Nuryanti. Production of kombucha from Muntingia calabura L. leaves and evaluation of its antibacterial activity and total flavonoid content. Journal of Applied Pharmaceutical Science 2022; 12(8), 187-192.
[11] LT Phung, H Kitwetcharoen, N Chamnipa, N Boonchot, S Thanonkeo, P Tippayawat, P Klanrit, M Yamada and P Thanonkeo. Changes in the chemical compositions and biological properties of kombucha beverages made from black teas and pineapple peels and cores. Scientific Reports 2023; 13, 7859.
[12] JT Oliveira, FM da Costa, TG da Silva, GD Simões, EDS Pereira, PQ da Costa, R Andreazza, PC Schenkel and S Pieniz. Green tea and kombucha characterization: Phenolic composition, antioxidant capacity and enzymatic inhibition potential. Food Chemistry 2023; 408, 135206.
[13] AN Mfopa, R Kemzeu, R Fokom, LRT Yamthe, D Dize and FF Boyom. Phenolic compounds, antioxidant and antileishmanial activities of kombucha as affected by fermentation time. Heliyon 2024; 10(22), e40463.
[14] S Wang, L Zhang, L Qi, H Liang, X Lin, S Li, C Yu and C Ji. Effect of synthetic microbial community on nutraceutical and sensory qualities of kombucha. International Journal of Food Science & Technology 2020; 55(10), 33327-3333.
[15] KR Lee, K Jo, KS Ra, HJ Suh and KB Hong. Kombucha fermentation using commercial kombucha pellicle and culture broth as starter. Food Science and Technology 2022; 42, e70020.
[16] S Moghadamtousi, M Fadaeinasab, S Nikzad, G Mohan, H Ali and H Kadir. Annona muricata (Annonaceae): A review of its traditional uses, isolated acetogenins and biological activities. International Journal of Molecular Sciences 2015; 16(7), 15625-15658.
[17] P Mohitha and H Shanmugam. Chemometric analysis and quantification of major anti-cancer activity compounds from leaves of soursop (Annona muricata Linn.). Chemical Papers 2023; 77(10), 6069-6081.
[18] CC Egbo, DC Igboaka and PF Uzor. Antimicrobial assay and GC-MS profile of the extract of the endophytic fungus from Annona muricata (Annonaceae) leaf. Tropical Journal of Natural Product Research 2024; 8(4), 7030-7034.
[19] E Zubaidah and Y Hasfiani. Physicochemical properties and anti-cancer activity of javanese turmeric kombucha (Curcuma xanthorrhiza) against T47D cell line. Trends in Sciences 2025; 22(9), 10417.
[20] E Zubaidah, ZM Putri, H Sujuti, AP Rahayu and T Ardyati. Physicochemical characteristics of kombucha based on various concentration of white turmeric (Curcuma zedoaria (Berg.) Roscoe). Biocatalysis and Agricultural Biotechnology 2024; 56, 102998.
[21] S Khosravi, M Safari, Z Emam-Djomeh and MT Golmakani. Development of fermented date syrup using kombucha starter culture. Journal of Food Processing and Preservation 2019; 43(8), e13872.
[22] S Muhammad, R Attia, M Shehata, D Zaki and G Hussein. Synthesis, characterization and application of a novel extracting agent 2-(4-methylphenyl)-8,9-dihydro-8,8-dimethyl-7H-pyrazolo[1,5-a]quinazolin-6-one (MPHPQ) for U(VI) removal from sulfate aqueous solutions. Egyptian Journal of Chemistry 2023; 66(3), 197-207.
[23] J Jeremic, I Vongluanngam A Ricci, GP Parpinello and A Versari. The oxygen consumption kinetics of commercial oenological tannins in model wine solution and chianti red wine. Molecules 2020; 25(5), 1215.
[24] S Shi, Y Wei, X Lin, H Liang, S Zhang, Y Chen, L Dong and C Ji. Microbial metabolic transformation and antioxidant activity evaluation of polyphenols in kombucha. Food Bioscience 2023; 51, 102287.
[25] D Pebiyanti, R Widiantie and A Prianto. Analysis of chemical characteristics, flavonoids, and organoleptics on shallot skin (Allium cepa) kombucha. Jurnal Pendidikan Biologi 2021; 6(2), 166-177.
[26] J Hou, R Luo, H Ni, K Li, FC Mgomi, L Fan and L Yuan. Antimicrobial potential of kombucha against foodborne pathogens. Quality Assurance and Safety of Crops & Foods 2021; 13(3), 53-61.
[27] YN Rao. Variation of the content and antioxidant activity of active substances in the kombucha fermentation of mulberry leaves. Highlights in Science, Engineering and Technology 2024; 91, 396-401.
[28] AN Artanti, A Febriyanty, F Prihapsara and DE Ermawati. Formulation optimization and antioxidant test for self-nano emulsifying drug delivery system of soursop leaves (Annona muricata L.) chloroform extract using candlenut oil as oil phase. Pharmaciana 2021; 11(1), 143.
[29] K Jakubczyk, Ł Łopusiewicz, J Kika, K Janda and K Skonieczna‐Żydecka. Fermented tea as a food with functional value—its microbiological profile, antioxidant potential and phytochemical composition. Foods 2023; 13(1), 50.
[30] Y Liu, Y Zheng, W Wang, Z Wang, S Han and P Zhou. Kombucha enables to inhibit digestive enzymes activity and adipocyte differentiation of OP9 cells. Journal of Food Science 2024; 89(12), 10053-10063.
[31] CM Park, GM Kim and GS Cha. Biotransformation of flavonoids by newly isolated and characterized Lactobacillus pentosus NGI01 strain from kimchi. Microorganisms 2021; 9(5), 1075.
[32] Ż Arciszewska, S Gama, M Kalinowska, G Świderski, R Świsłocka, E Gołębiewska, M Naumowicz, M Worobiczuk, A Cudowski and B Godlewska-Żyłkiewicz. Caffeic Acid/Eu(III) complexes: Solution equilibrium studies, structure characterization and biological activity. International Journal of Molecular Sciences 2022; 23(2), 888.
[33] A Zeb. Concept, Mechanism, and Applications of phenolic antioxidants in foods. Journal of Food Biochemistry 2020; 44(9), e13394.
[34] H Kim, J Jeon, J Lee, C Song, B Gu, NM Kim, Y Tae-Hui, S Oh, S Park, K Pal, GJ Kim and D Kim. Utilizing kombucha culture for coffee fermentation and biochemical characteristic analysis. Current Research in Food Science 2025; 10, 100996.
[35] SA Villarreal-Soto, S Beaufort, J Bouajila, S Jean-Pierre, T Renard, S Rollan and P Taillandier. Impact of fermentation conditions on the production of bioactive compounds with anticancer, anti-inflammatory and antioxidant properties in kombucha tea extracts. Process Biochemistry 2019; 83, 44-54.
[36] H Kitwetcharoen, Y Phannarangsee, P Klanrit, S Thanonkeo, P Tippayawat, P Klanrit, P Klanrit, M Yamada and P Thanonkeo. Functional kombucha production from fusions of black tea and indian gooseberry (Phyllanthus emblica L.). Heliyon 2024; 10(24), e40939.
[37] S Selvaraj and K Gurumurthy. Metagenomic, organoleptic profiling, and nutritional properties of fermented kombucha tea substituted with recycled substrates. Frontiers in Microbiology 2024; 15, 1367697.
[38] WC Tan, BJ Muhialdin and ASM Hussin. Influence of storage conditions on the quality, metabolites, and biological activity of soursop (Annona muricata L.) kombucha. Frontiers in Microbiology 2020; 11, 603481.
[39] RI Limón, E Peñas, MI Torino, C Martínez‐Villaluenga, M Dueñas and J Frı́as. Fermentation enhances the content of bioactive compounds in kidney bean extracts. Food Chemistry 2015; 172, 343-352.
[40] DM Kopustinskiene, V Jakstas, A Savickas and J Bernatoniene. Flavonoids as anticancer agents. Nutrients 2020; 12(2), 457.
[41] C George Vazhappilly, M Amararathna, AC Cyril, R Linger, R Matar, M Merheb, WS Ramadan, R Radhakrishnan and HPV Rupasinghe. Current methodologies to refine bioavailability, delivery, and therapeutic efficacy of plant flavonoids in cancer treatment. Journal of Nutritional Biochemistry 2021; 94, 108623.
[42] Z He, H Zhang, T Wang, R Wang and X Luo. Effects of five different lactic acid bacteria on bioactive components and volatile compounds of oat. Foods 2022; 11(20), 3230.
[43] C Bas-Bellver, C Barrera and L Seguí. Impact of thermophysical and biological pretreatments on antioxidant properties and phenolic profile of broccoli stem products. Foods 2024; 13(22), 3585.
[44] J Yang, H Lee, J Sung, Y Kim, HS Jeong and J Lee. Conversion of rutin to quercetin by acid treatment in relation to biological activities. Preventive Nutrition and Food Science 2019; 24(3), 313-320.
[45] TP Van, QK Phan, HP Quang, GB Pham, NHN Thi, HTT Thi and AD Do. Multi-strain probiotics enhance the bioactivity of cascara kombucha during microbial composition-controlled fermentation. Preventive Nutrition and Food Science 2023; 28(4), 502-513.
[46] DP Sinaga, DKA Tampubolon, RF Kembaren and MM Martgrita. Fermentation process effect to enhance antioxidant and antibacterial activity of phenolic compounds and its possible application to galactomannan polysaccharides: A review. IOP Conference Series: Earth and Environmental Science 2022; 1097(1), 12027.
[47] R Jafari, NS Naghavi, K Khosravi‐Darani, M Doudi and K Shahanipour. Isolation, molecular and phylogenetic identification of microorganisms from kombucha solution and evaluation of their viability using flow cytometery. Food Science and Technology 2022; 42, 63220.
[48] ADF Nascimento, SMT Ramos, VN Bergamo, EDS Araujo and GA Valencia. Pickering emulsions stabilized using bacterial cellulose from kombucha. Starch 2025; 77(2), 2400103.
[49] NF Lima, GM Maciel, IDAA Fernandes and CWI Haminiuk. Optimising the production process of bacterial nanocellulose: Impact on growth and bioactive compounds. Food Technology and Biotechnology 2023; 61(4), 494-504.
[50] J Gong, S Rao, X Liu, S Cheng, X Cong and D Zhu. Microbiome-metabolome integration reveals fermentation-driven divergence in bioactive compound profiles of cardamine violifolia leaf pickles. Current Research in Food Science 2025; 11, 101121.
[51] JY Kim, TTP Dao, K Song, SB Park, H Jang, MK Park, SU Gan and YS Kim. Annona muricata leaf extract triggered intrinsic apoptotic pathway to attenuate cancerous features of triple negative breast cancer MDA-MB-231 cells. Evidence-Based Complementary and Alternative Medicine 2018; 2018, 7972916.
[52] I Rady, MB Bloch, NC Roxane-Cherille, SB Mbeumi, MR Anwar, H Mohamed, AS Babatunde, K Jules-Roger, FK Noubissi, KAE Sayed, GK Whitfield and JC Chamcheu. Anticancer properties of graviola (Annona muricata): A comprehensive mechanistic review. Oxidative Medicine and Cellular Longevity 2018; 2018, 1826170.
[53] C Cho, A Teghanemt, MA Apicella and WM Nauseef. Modulation of phagocytosis-induced cell death of human neutrophils by Neisseria gonorrhoeae. Journal of Leukocyte Biology 2020; 108(5), 1543-1553.
[54] Z Pan, X Zhang, P Yu, X Chen, P Lu, M Li, X Liu, Z Li, F Wei, K Wang, Q Zheng and D Li. Cinobufagin induces cell cycle arrest at the G2/M phase and promotes apoptosis in malignant melanoma cells. Frontiers in Oncology 2019; 9, 853.
Supplementary material
Supplementary 1 LC-MS based classification of compounds in soursop leaf drink and kombucha with functional implications.
Classification |
Compounds |
Soursop leaf drink |
Soursop leaf kombucha |
Functional effect |
Alkaloid |
Oxeladin |
X |
√ |
Anticancer (1) |
|
Sinomenine |
√ |
√ |
Antimetastatic (2) Immunomodulator and anti-inflammatory (3) |
|
Reboxetine |
√ |
√ |
Anti-inflammatory and neuroprotective (4) |
|
Reticuline |
√ |
X |
Anticancer (5) Anti-inflammatory (6) |
Acetogenin |
Annonahexocin |
√ |
√ |
Anticancer (7) |
|
Annomuricin E |
√ |
√ |
Anticancer (8) |
|
Annonacin |
√ |
X |
Anticancer (9) Antimetastatic (10) |
|
Squamocin |
√ |
X |
Anticancer and Antimetastasis (11) Antitumor (12) Immunomodulator (13) |
Flavonoid |
Justicidin B |
√ |
√ |
Anticancer (14) |
|
Kaempferol-3-O-rutinoside |
√ |
X |
Anti-inflammatory (15) |
|
Rutin |
√ |
X |
Anti-inflammatory (16) |
|
Kaempferol |
X |
√ |
Antioxidant and Antiapoptotic (17) Anti-inflammatory (18) |
|
Quercetin |
X |
√ |
Anti-inflammatory (19) Anticancer (20) |
Terpenoid |
Genipin 1-gentiobioside |
√ |
√ |
Anticancer (21) |
|
Citronellyl acetate |
√ |
X |
Antioxidant (22) |
Monoterpen |
Citronellic acid |
√ |
√ |
Anti-inflammatory and immunomodulator (23) |
Diterpenoid |
Cryptotanshinone |
X |
√ |
Antioxidant (24) |
|
Dihydrotanshinone I |
X |
√ |
Anti-inflammatory (25) Antioxidant (26) |
Triterpenoid |
Cucurbitacin E |
X |
√ |
Immunosuppressants (27) Antiproliferation (28) |
Tetraterpenoid |
Spirilloxanthin |
√ |
√ |
Antioxidant (29) |
Fenilpropanoid |
Asarone |
√ |
X |
Anti-inflammatory (30) Anticancer (31) |
|
Coumarin |
X |
√ |
Antioxidant, Antimicrobial, and Anti-inflammatory (32) |
|
Isocoumarin |
X |
√ |
Anti-inflammatory (33) Anticancer (34) |
Hydroxycoumarin |
Isofraxidin |
X |
√ |
Antioxidant (35) Anti-inflammatory (36) |
Amino acid |
Proline |
√ |
√ |
Anticancer dan Anti-inflammatory (37) Antioxidant (38) |
|
Isoleucine |
√ |
X |
Antioxidant dan Anti-inflammatory (39) |
|
4-acetamidobutanoic acid |
X |
√ |
Antioxidant (40) Antitumor and immunosuppressive (41) |
Amin |
Choline |
√ |
√ |
Anti-inflammatory (42) |
|
Solsolinol |
√ |
X |
Anti-inflammatory (43) |
|
Dibenzylamine |
√ |
X |
Antioxidant (44) |
Fatty acid |
Myristoleic acid |
√ |
X |
Antioxidant (45) |
|
Oleic acid |
√ |
X |
Antitumor (46) Antioxidant (47) |
|
Eicosenoic acid |
√ |
X |
Anti-melanogenesis (48) |
|
butyric acid |
√ |
√ |
Anticancer (49) Immunomodulator (50) |
|
Jasmonic acid |
X |
√ |
Anticancer (51) |
|
Traumatic acid |
X |
√ |
Anticancer (52) |
Steroid |
Nandrolone |
√ |
X |
Anticancer (53) |
|
Stigmasterol glucoside |
√ |
X |
Antioxidant (54) Anti-inflammatory (55) |
|
Stigmasterol |
X |
√ |
Anti-inflammatory and Anticancer (56) |
|
Eplerenone |
X |
√ |
Anti-inflammatory (57) Anticancer (58) |
Fatty alcohol |
1-Heneicosanol |
X |
√ |
Anticancer (59) |
Saponin |
Momordicoside G |
√ |
X |
Anticancer and Anti-inflammatory (60) |
Piperidin |
TEMPOL (4-Hydroxy-tempol) |
X |
√ |
Antioxidant (61) |
Heterocyclic oxygen |
Furan |
X |
√ |
Anticancer (62) Antioxidant (63) |
Benzopiran |
Chromone |
X |
√ |
Anti-inflammatory (64) Anticancer (65) Antimicrobial (66) |
Organic acid |
2-Propenoic acid |
X |
√ |
Anticancer (67) |
|
Crotonic acid |
X |
√ |
Anticancer (68) |
|
Glucuronic acid |
X |
√ |
Anti-inflammatory (69) |
|
Lactic acid |
X |
√ |
Immunomodulator (70) Anticancer (71) |
|
Acetic acid |
X |
√ |
Antiproliferation (72) |
Benzoquinolines |
Acridine |
X |
√ |
Antiproliferation (73) Immunomodulator (74) |
|
9-Aminoacridine |
X |
√ |
Anticancer (75) |
|
Syringol |
√ |
√ |
Anti-inflammatory (76) |
|
Bisoprolol |
X |
√ |
Anti-inflammatory (77) Anticancer (78) |
|
Guaiacol |
X |
√ |
Anticancer (79) |
Benzofuran |
Dihydroactinidiolide |
√ |
√ |
Anti-inflammatory (80) |
Dihydrofurans |
Furanon |
√ |
X |
Anticancer (81) |
Aryl-phenylketones |
Ketorolac |
√ |
√ |
Anticancer (82) Anti-inflammatory (83) |
Aromatic compounds |
2-Furoic acid |
X |
√ |
Immunomodulator dan anticancer (84) |
|
Furfural |
√ |
X |
Ant-inflammatory (85) |
Alcohol furan |
Furfuranol |
X |
√ |
Antimicrobial (86) |