Germinated Cempo Ireng Black Rice (Oryza sativa L.) Demonstrates Functional Anti-Obesity Effects in High-Fat Diet-Induced Rats
Mina Parhatus Salamah1, Yunika Mayangsari1, Hitoshi Shirakawa2,
Afifah Zahra Agista2 and Wahyu Dwi Saputra1,*
1Department of Food and Agricultural Product Technology, Universitas Gadjah Mada, Yogyakarta 55281, Indonesia
2Laboratory of Nutrition, Graduate School of Agricultural Science, Tohoku University, Miyagi 980-8572, Japan
(*Corresponding author’s e-mail: [email protected])
Received: 21 November 2025, Revised: 11 December 2025, Accepted: 18 December 2025, Published: 15 March 2026
Abstract
Obesity is a long-term metabolic disorder caused by an imbalance between energy consumption and energy expenditure, leading to excessive fat accumulation. This condition is closely associated with metabolic disorders, thus requiring effective strategies for prevention and management. One such approach is the consumption of functional foods, such as whole grains like black rice, whose functional value can be enhanced through germination. This study aimed to evaluate the effects of administering germinated black rice (GBR) to rats with a high-fat diet (HFD), compared with non-germinated black rice (NGBR) and white rice (WR). The results showed that GBR significantly suppressed body weight gain, liver mass, and adipose tissue mass, approaching levels observed in the control group. GBR also improved lipid profiles by reducing total cholesterol, triglycerides, and LDL, while increasing HDL and cecal cholesterol excretion. The most significant reduction in blood glucose levels was observed in the GBR group (Δ −49 mg/dL), accompanied by a decrease in hepatic malondialdehyde levels, indicating a reduction in oxidative stress. Additionally, GBR increased the concentrations of acetate, propionate, and butyrate, which contribute to metabolic improvement. Histopathological analysis revealed smaller adipocyte size and reduced hepatic steatosis in the GBR group. Gene expression analysis revealed significant downregulation of Hmgcr, suggesting an inhibition of hepatic cholesterol biosynthesis. These results indicate that germinated black rice exerts stronger anti-obesity effects than NGBR and WR, supporting its potential as a functional food for obesity prevention.
Keywords: Germinated black rice, Obesity, High-fat diet, Functional food
Introduction
Obesity is a chronic disease characterized by excessive fat accumulation due to an imbalance between energy intake and energy expenditure over a long period [1]. This condition is a significant risk factor for various degenerative diseases, including hypertension, cardiovascular disease, type 2 diabetes, and cancer [2]. The prevalence of obesity continues to increase globally. The WHO estimates that by 2025, almost 167 million individuals will suffer health deterioration as a result of being overweight or obese. This increase is primarily due to a high-calorie diet and a sedentary lifestyle. The high consumption of processed and fast
foods that are high in fat and low in fiber, fruits, and vegetables, is exacerbating the obesity epidemic [3,4].
These figures confirm that obesity is a serious problem requiring attention, as well as effective intervention strategies to mitigate it. Actually, this condition can still be prevented by following the WHO recommendations, which include restricting calorie intake from fats and carbohydrates, increasing consumption of whole grains, legumes, fruits, and vegetables, and engaging in regular physical activity [5]. Recently, researchers have shown significant interest in non-pharmacological approaches, particularly the consumption of whole-grain functional foods. Whole grains provide a source of complex carbohydrates and essential nutrients, while also being abundant in dietary fiber and bioactive substances that contribute to metabolic health maintenance. Black rice is distinguished among whole grains by its elevated anthocyanin pigment concentration in the aleurone/bran layer and its substantial dietary fiber content. These 2 components are recognized for their contributions to antioxidant, hypolipidemic, and anti-inflammatory activities [6]. For example, a previous study demonstrated that replacing 20% of the meal with Brazilian black rice variety IAC 600 effectively lowered plasma cholesterol, triglycerides, and LDL levels in hyperlipidemic rats.
Post-harvest alteration through the germination process provides numerous advantages for black rice. This process enhances the bioavailability of bioactive compounds by degrading complex substances such as flavonoids, phenolics, and pyhtates into simpler forms, facilitating their absorption by the body [7]. Moreover, germination increases levels of specific nutrients, such as free amino acids and γ-aminobutyric acid (GABA), which are crucial for regulating fat metabolism and blood pressure [8]. This process increases the concentration of functional compounds, including anthocyanins, phenolic acids, and γ-oryzanol, which are recognized for their antioxidant and anti-inflammatory properties. The elevated levels of these bioactive compounds render germinated black rice superior to non-germinated rice, especially regarding its potential as a functional food for the prevention of obesity and associated metabolic disorders.
Numerous studies have shown that consuming foods rich in fiber and bioactive compounds can alter the expression of genes associated with lipid metabolism and inflammation. Compounds such as anthocyanins, γ-oryzanol, and phenolic acids modulate molecular signaling pathways, consequently inhibiting lipogenesis, enhancing fat oxidation, and diminishing systemic inflammation [9]. Furthermore, consumption of whole grains rich in bioactive compounds such as ferulic acid and γ-Oryzanol has been reported can modulate lipid metabolism by activating the AMPKα pathway and suppressing lipogenic transcription factors such as SREBP, thereby influencing downstream targets including HMGCR and other cholesterol-synthesis enzymes [10,11].
In Indonesia, in addition to imported varieties, there is a local black rice variety called Cempo Ireng, extensively cultivated in Yogyakarta and Central Java. This variety is recognized for its elevated anthocyanin content and has excellent potential for Development as a local functional food. Previous investigations indicate that the black rice variety Cempo Ireng is abundant in anthocyanins [12,13] and has potent antioxidant activity [13]. Nevertheless, the majority of these studies remain confined to in vitro studies, so currently, in vivo evidence directly comparing the effects of germinated and non-germinated Cempo Ireng rice on obesity parameters, including body weight, adipose tissue mass, and regulation of gene expression related to lipid metabolism, is still very limited. Therefore, this study aims to investigate the efficacy of germinated Cempo Ireng black rice as an anti-obesity agent by in vivo investigation, and to compare its effects with those of non-germinated black rice and white rice. The result aims to establish a scientific foundation for the Development of locally produced functional foods to prevent and treat obesity.
Materials and methods
Plant materials
Cempo Ireng paddy was obtained from PD Agriwira Mandiri Sejahtera in Karanganyar Regency, Central Java, Indonesia, while the IR-64 white rice was from Sumber Hasil Bumi Bimomartani, Ngemplak, Sleman, Yogyakarta Special Region, Indonesia.
Black rice germination
Black rice samples were prepared through the hulling process. Whole kernels were separated from broken grains using a rice grader. Germination was carried out following the procedure by Munarko et al. [14] with some modifications. Dehulled black rice Cempo Ireng variety was soaked in a 0.1% hypochlorite solution for 30 min, then rinsed with distilled water. The rice was subsequently soaked in distilled water at a 1:1 (w/v) ratio at 35 °C and incubated at 33 °C for 24 h, with water replaced every 8 h. After soaking, the rice was aerated by spreading it on a sterile tray lined with tissue for 24 h at the same temperature, with distilled water sprayed every 8 h. Drying was carried out using a food dehydrator at 50 °C for 2 h. Flours from germinated and non-germinated rice samples were sieved through an 80-mesh sieve and stored at 4 ℃.
Dietary fiber measurements
Total dietary fiber was determined using the AOAC (1995) multi-enzyme method. Briefly, 0.5 of the sample was sequentially incubated with α-amylase, pepsin, and β-amylase under controlled pH and temperature conditions. The insoluble fraction was collected by filtration, washed with ethanol and acetone, dried at 105 °C overnight, and weighed. In contrast, the soluble fraction was precipitated from the filtrate with warm 95% ethanol, filtered, washed, dried, and weighed. Total dietary fiber was calculated as the sum of insoluble and soluble fractions using the formula:
Total Dietary Fiber = Insoluble Dietary Fiber + Soluble Dietary Fiber.
Animals experimental
A total of 30 male Sprague-Dawley (Rattus norvegicus) rats (8 weeks old, weighing 170 - 200g) were obtained from the Nutrition and Food Research Center, Universitas Gadjah Mada, Yogyakarta, Indonesia. The animals were individually housed in stainless-steel cages under controlled environmental conditions (temperature 24 ± 2 °C, relative humidity 85%, 12 h light/dark cycle, and adequate ventilation). All rats were acclimatized for 7 days with free access to food and water (ad libitum). All experimental procedures were approved by the Ethics Committee of Universitas Gadjah Mada (Ref. No. 00066/X/UN1/LPPT/EC/2024).
Induction of obesity
After acclimatization, the rats were randomly divided into 5 groups. The control group (n = 6) continuously consumed standard AIN-93M pellets during the induction period, while the obesity group (n = 24) was induced with a high-fat diet (HFD) by feeding a modified AIN-93M high-fat diet for 4 weeks. The HFD formulation based on Dourmashkin [15], it contained 50% fat, sourced from lard. The HFD was prepared weekly to prevent spoilage by thoroughly mixing all ingredients, extruding them, and drying them in an oven at 65 °C for 24 h. After the induction period, the average body weight of the HFD group was compared with that of the control group. Rats with a weight exceeding 10% of the maximum body weight of the normal diet rats were classified as obese [16]. Obesity was also confirmed using the following calculation:
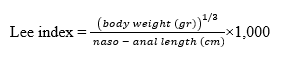
Rats with a Lee Index greater than 0.30 were classified as obese [7]. In addition, lipid profile analysis was performed after induction to confirm the development of obesity in the rats.
Treatment of GBR
After confirming that the HFD-induced rats had developed obesity after 4 weeks of induction, they were further divided into 4 groups: Positive control / HFD group (n = 6), obese rats induced with HFD and substituted with IR-64 white rice diet (WR) (n = 6), obese rats induced with HFD and substituted with non-germinated Cempo Ireng black rice diet (NGBR) (n = 6), and obese rats induced with HFD and substituted with germinated Cempo Ireng black rice diet (GBR) (n = 6). Each group was fed 15 - 20 g daily for 4 weeks. The control group (n = 6) and positive control / HFD group continued consuming standard AIN-93M pellets, while the 4 treatment groups were provided with iso-caloric diet formulations (Table 1).
Table 1 Isocaloric and isofiber diet formulation.
Ingredients (g/kg) |
AIN-93M Standard |
HFD |
WR |
NGBR |
GBR |
Corn starch |
620.692 |
341.25 |
- |
- |
- |
Rice flour |
- |
|
749.47 |
642.65 |
617.37 |
sucrose |
100 |
100 |
100 |
100 |
100 |
CMC |
50 |
50 |
40.38 |
7.20 |
4.13 |
Casein |
140 |
140 |
140 |
140 |
140 |
Soybean oil |
40 |
- |
40 |
40 |
40 |
Lard |
- |
258.3 |
- |
- |
- |
AIN-93 Mineral Mix |
35 |
35 |
35 |
35 |
35 |
AIN-93 Vitamin Mix |
10 |
10 |
10 |
10 |
10 |
L-Cysteine |
1.8 |
1.8 |
1.8 |
1.8 |
1.8 |
Choline Bitartrate |
2.5 |
2.5 |
2.5 |
2.5 |
2.5 |
TBHQ |
0.008 |
0.008 |
0.008 |
0.008 |
0.008 |
Total |
1.000 |
939 |
1.119,16 |
979,16 |
950,80 |
Energy (kcal/kg) |
3.554 |
4.650 |
3.554 |
3.554 |
3.554 |
The AIN-93M diet [17] and a high-fat diet (HFD) [15] were used with modifications.
Anthropometric and dietary measurements
Body weight and food intake were monitored regularly throughout the intervention period. Blood samples were collected from the retro-orbital vein at baseline (pre-induction) and after the intervention (post-treatment).
Biochemical determinations
Plasma was used to determine the levels of total cholesterol, triglycerides, low-density lipoprotein (LDL) cholesterol, high-density lipoprotein (HDL) cholesterol, cecal cholesterol excretion, and glucose levels using commercial diagnostic kits (DiaSys Diagnostic Systems GmbH, Germany). Short-chain fatty acids (SCFAs) were analyzed by gas chromatography equipped with a flame ionization detector (GC-FID) (Shimadzu GC-2010 Plus). Lipid peroxidation was assessed using the TBARS (Thiobarbituric Acid Reactive Substances) method.
Histopathological liver and white adipose tissue
The liver and white adipose tissue were fixed in a 10% formalin buffer solution for 24 h. The fixed tissue was then routinely processed for paraffin block preparation and stained using hematoxylin and eosin (H&E). The stained preparations were observed using an optical microscope (Olympus CX33 Trinocular LED; Tokyo, Japan). Liver images were then analyzed semi-visually and quantitatively using ImageJ software (National Institutes of Health, USA).
Quantitative real-time PCR (qRT-PCR) analysis
Liver tissues were preserved in RNAlater, and total RNA was isolated using Isogen reagent. The extracted RNA was purified and reverse transcribed into cDNA using a commercial reverse transcription kit (according to the manufacturer’s instructions). Quantitative real-time PCR (qRT-PCR) was performed with SYBR Premix Ex Taq (Takara Bio, Otsu, Japan) and gene-specific primers as shown in Table 2 on a Bio-Rad CFX Connect Real-Time PCR Detection System (Hercules, CA, USA). Relative mRNA expression levels were normalized to Eukaryotic translation elongation factor 1α1 (Eef1α1) as the internal reference gene. The relative hepatic mRNA expression of lipid metabolism, namely Fasn (Fatty acid synthase), Hmgcr (3-Hydroxy-3-methylglutaryl-CoA Reductase), and Lxr-α (Liver X receptor alpha).
Table 2 Primer list of genes used for qRT-PCR analysis.
Gene name |
Forward/Reverse |
Primer sequence |
Fasn |
Forward Reverse |
GGCTCACACACCTACGTATTGG TGCTTAATGAAGAAGCATATGGCTT |
Hmgcr |
Forward Reverse |
AATTGTGTGTGGCACTGTGATG GATCTGTTGTGAACCATGTGACTTCT |
Lxr-α |
Forward Reverse |
ACCTCTGCGATCGAGGTGAT |
Statistical analysis
Data were analyzed using 1-way analysis of variance (ANOVA), followed by the Tukey test using IBM SPSS software version 27. When the assumptions of normality or homogeneity of variance were not met, the Kruskal-Wallis test was applied. Statistical significance was set at p < 0.05.
Results and discussion
Dietary fiber
This study analyzed the dietary fiber content in white rice (WR), non-germinated black rice (NGBR), and germinated black rice (GBR). Our results showed that the highest total dietary fiber (TDF) content was found in GBR at 7.43%, followed by NGBR at 6.66%, while WR had the lowest fiber content at 1.28%. The increase in TDF in GBR mainly came from the insoluble dietary fiber (IDF) fraction, which was also significantly higher than in WR and NGBR. Conversely, the soluble dietary fiber (SDF) fraction was relatively smaller than the IDF. However, the analysis showed significant differences between groups (Figure 1). These results indicate that the germination process of black rice increases dietary fiber content, with both soluble and insoluble fractions increasing, with insoluble fiber predominating.
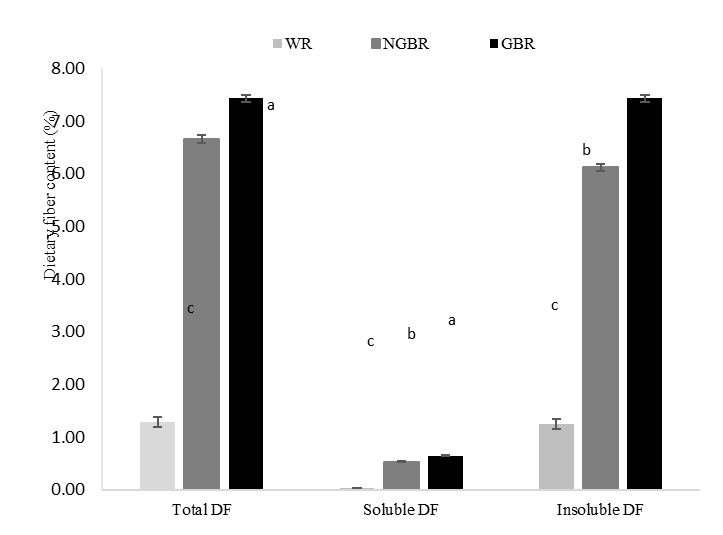
Figure 1 Dietary fiber content of IR-64 white rice (WR), Cempo Ireng black rice non-germinated (NGBR), and Cempo Ireng black rice germinated for 48 h (GBR) was obtained from 3 replications (mean ± SD) (n = 3). Different letters in the same column indicate statistically significant differences among samples (p < 0.05).
The increase in dietary fiber content observed in this study is consistent with previous findings showing that germination can enhance dietary fiber levels in different brown rice varieties [18]. In addition, prior research has demonstrated that germination in the dark can increase IDF by up to 55% compared to light conditions, by 34%. This is associated with metabolic processes in the dark that are more focused on the formation of new cell structures and the provision of energy for seedling growth, thereby increasing cellulose microfibril synthesis and accumulation [19].
Physiologically, germination triggers an increase in the activity of hydrolytic enzymes, particularly β-amylase, which hydrolyzes starch into simple sugars [20]. In the cotyledon, starch is broken down into smaller molecules such as glucose and fructose to provide energy for embryo growth [21]. In general, α-amylase activity is consistent with the pattern of starch degradation, so that the longer the germination process, the greater the conversion of starch into simple sugars. The breakdown of starch reserves increases the proportion of non-starch components, including dietary fiber, which makes up cell walls such as cellulose, hemicellulose, and lignin.
The effect of germinated black rice administration on body weight gain and feed intake
Changes in body weight of rats during the 9-week intervention are shown in (Figure 2(a)). Body weight increased gradually in all groups, except for NGBR and GBR, which tended to remain stable. Repeated-measures ANOVA showed significant differences across weeks (p < 0.001) and a week × group interaction (p = 0.02). From week 4 to week 9, the HFD and WR groups had significantly higher body weights than the control (p < 0.05), while the NGBR and GBR groups showed lower body weights than the HFD and WR groups in weeks 8 and 9, and were not significantly different from the control.
Feed consumption patterns in Figure 2(b) did not differ significantly between groups (p > 0.05), suggesting that metabolic factors and feed composition were more likely to have influenced differences in body weight. These results are consistent with previous studies reporting that rats fed diets containing 50% and 100% substitution of germinated brown rice (GBR) exhibited significant reductions in body weight and feed intake after 8 weeks, and that a similar trend was observed in HFD rats substituted with germinated brown rice IR-64 for 28 days. However, the differences were not statistically significant [22,23]. The weight loss in the Cempo Ireng germinated black rice group is thought to be related to its high fiber content, which reduces feed intake. Dietary fiber can enhance satiety by increasing the volume and viscosity of food in the stomach [24]. In addition, viscous soluble fiber can increase lumen viscosity, slow gastric emptying, and form a mechanical barrier that inhibits starch digestion in the small intestine, thereby reducing energy absorption [25]. Furthermore, bioactive compounds increase during germination and play a role in inhibiting fat accumulation and increasing lipid oxidation [26,27].
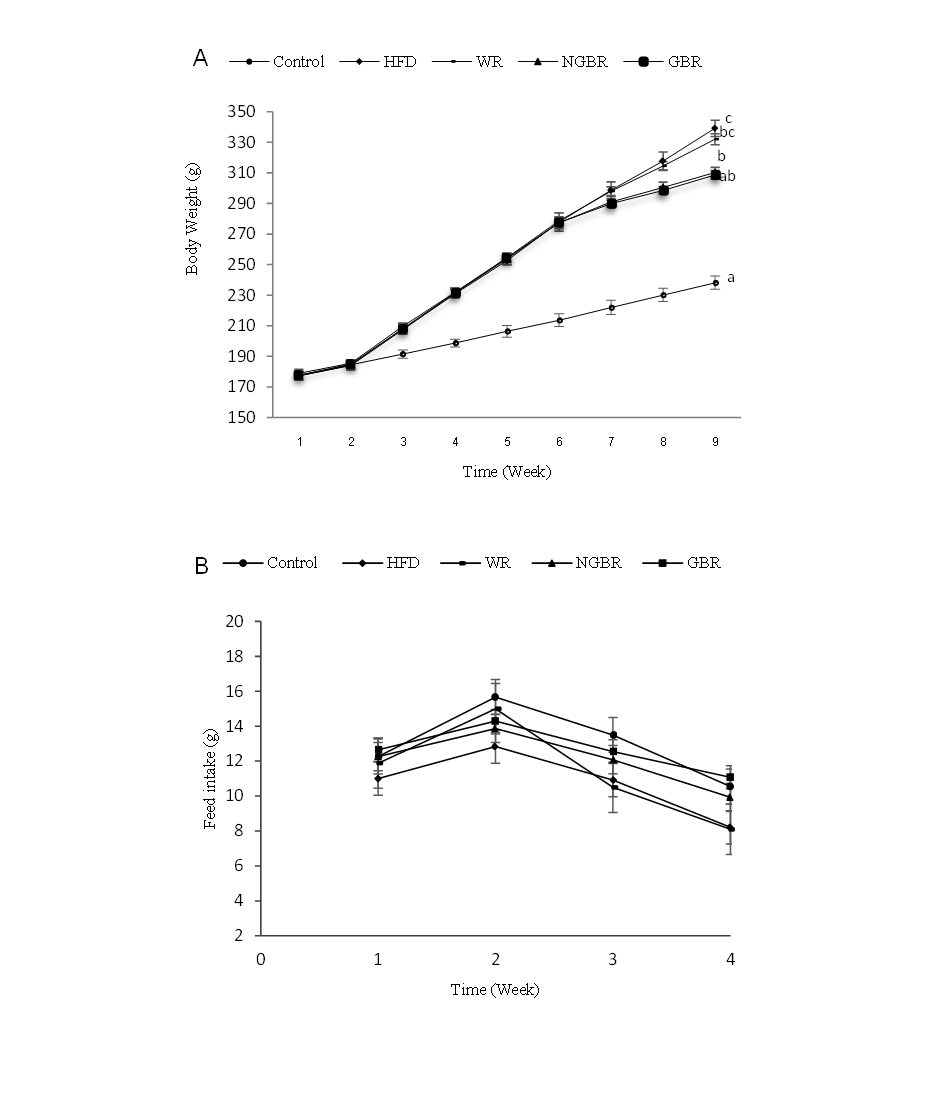 Figure
2 The
effect of germinated black rice administration on body weight and
feed intake of rats during the induction and intervention periods.
Changes in (a) body weight and (b) feed intake were monitored
throughout the experimental period. Standard AIN-93M (Control),
high-fat diet (HFD), high-fat diet + white rice (WR), high-fat diet
+ non-germinated black rice (NGBR), and high-fat diet + germinated
black rice (GBR). Data
are presented as mean
± SD (n = 6). Different letters in the same column indicate
statistically significant differences among samples (p
< 0.05).
Figure
2 The
effect of germinated black rice administration on body weight and
feed intake of rats during the induction and intervention periods.
Changes in (a) body weight and (b) feed intake were monitored
throughout the experimental period. Standard AIN-93M (Control),
high-fat diet (HFD), high-fat diet + white rice (WR), high-fat diet
+ non-germinated black rice (NGBR), and high-fat diet + germinated
black rice (GBR). Data
are presented as mean
± SD (n = 6). Different letters in the same column indicate
statistically significant differences among samples (p
< 0.05).
The effect of germinated black rice administration on biochemical and histopathological parameters
Lee index, liver and adipose tissue weights
The mean liver weight, adipose tissue, Lee index, and organ-to-body weight ratio are presented in Table 3. High-fat diet induction was proven to be effective in triggering obesity in the treatment groups. This is demonstrated by a significant increase in the Lee index in the HFD, WR, NGBR, and GBR groups compared with the control group (p < 0.05), with all treatment groups showing a Lee index ≥ 300 before intervention, indicating obesity in rats [28].
The result showed that the HFD and WR groups had significantly higher liver weights than the Control, NGBR, and GBR groups (p < 0.05). Conversely, GBR (8.98 ± 0.06 g) had the lowest liver weight, which was significantly lower than that of all other groups. Adipose tissue weight and the ratio of adipose tissue to body weight also increased significantly in the HFD and WR groups compared to NGBR and GBR (p < 0.05), with GBR showing the lowest values (1.91 ± 0.05 g; 0.60%). There were no significant differences between GBR and control in adipose parameters, either absolute or ratio.
Table 3 Average Lee index, liver weight, and adipose tissue weight of experimental rats.
No |
Group |
Lee index (Induction) |
Liver Weight (g) |
Adipose tissue weight (g) |
Ratio liver weight (%) |
Ratio adipose tissue weight (%) |
1 |
Control |
286.41 ± 2.05a |
9.49 ± 0.60a |
1.93 ± 0.10a |
3.86c |
0.78b |
2 |
HFD |
317.84 ± 3.21bc |
13.59 ± 0.70c |
5.52 ± 0.42c |
3.77c |
1.53d |
3 |
WR |
321.54 ± 3.09b |
13.54 ± 0.41c |
5.03 ± 0.39c |
3.86c |
1.43d |
4 |
NGBR |
322.25 ± 3.83b |
10.15 ± 0.11bc |
2.83 ± 0.08b |
3.17b |
0.88c |
5 |
GBR |
316.84 ± 2.18b |
8.98 ± 0.06a |
1.91 ± 0.05a |
2.82a |
0.60a |
Standard AIN-93M (Control), high-fat diet (HFD), high-fat diet + white rice (WR), high-fat diet + non-germinated black rice (NGBR), and high-fat diet + germinated black rice (GBR). Data are presented as mean ± SD (n = 6). Different letters in the same column indicate statistically significant differences among samples (p < 0.05).
A similar pattern was observed in liver weight ratio, where GBR at 2.82% and NGBR at 3.17% were significantly lower than the HFD and WR groups (p < 0.05). Overall, HFD increased liver and adipose tissue weight, whereas administration of germinated black rice effectively suppressed the obesity-inducing effect of HFD, with values approaching those of the control group. Brown rice germination administration has been reported to reduce white adipose tissue mass, adipocyte size, and hepatic steatosis in obese rats [23]. In addition, brown rice sprout extract can reduce lipid accumulation in the liver and adipose tissue by decreasing the expression of lipogenic genes (SREBP-1c, FASN), thereby reducing de novo lipogenesis [29].
Plasma lipid profile
Blood lipid profile analysis includes total cholesterol (TC), triglycerides (TG), low-density lipoprotein cholesterol (LDL), and high-density lipoprotein cholesterol (HDL) levels, which reflect lipid metabolism in the intestine. Lipid profile analysis is an essential biomarker for describing the risk of organ damage. The results of lipid profile analysis after intervention showed changes in levels due to dietary treatment. The differences between groups were significant, with analysis based on the change difference (Δ post–pre). The GBR and NGBR diets significantly improved the plasma lipid profile of obese rats compared with the HFD group. Both diets reduced total cholesterol (−134.66 ± 4.26; −120.45 ± 4.89 mg/dL), triglycerides (−82.76 ± 3.27; −60.90 ± 5.47 mg/dL), and LDL (−131.16 ± 3.30; −93.93 ± 3.80mg/dL) significantly (p < 0.05), while the HFD and control groups tended to increase (p > 0.05). The increase in HDL, known as the good cholesterol, was also significant in BRG (+63.54 ± 1.17 mg/dL) and NGBR (+61.08 ± 2.22 mg/dL), whereas the HFD and control groups showed a decrease (Table 4).
The results showed that germinated black rice intervention improved the lipid profile in HFD-induced obese rats. Significant decreases in total cholesterol, triglycerides, and LDL levels, along with increases in HDL levels, confirmed a significant hypolipidemic effect in the GBR group compared with the HFD and control groups. These findings are consistent with previous studies reporting that rice germination, including germinated brown rice (GBR), effectively lowers lipid profiles in HFD-induced obese rats [24,30,31].
Consistently, these results are supported by other literature reporting that pigmented rice, especially black rice, is rich in bioactive compounds such as anthocyanins, γ-oryzanol, tocopherols, and dietary fiber, which help reduce the risk of dyslipidemia. In addition, the germination process in black rice is known to increase the bioavailability of bioactive compounds, such as γ-aminobutyric acid (GABA), free phenols, and bioactive peptides that have beneficial effects on lipid metabolism [8]. Dietary fiber also plays a significant role in lowering lipid profiles through several mechanisms, including binding bile acids and cholesterol during micelle formation in the intestinal lumen and increasing the excretion of bile acids and steroids in feces [32].
Table 4 Plasma lipid profile (total cholesterol, LDL, HDL, and triglycerides) of experimental rats.
Group |
Total cholesterol (mg/dL) |
Triglyceride (mg/dL) |
LDL (mg/dL) |
HDL (mg/dL) |
||||||||
Pre Intervention |
Post Intervention |
Δpost-pre |
Pre Intervention |
Post Intervention |
Δpost-pre |
Pre Intervention |
Post Intervention |
Δpost-pre |
Pre-Intervention |
Post Intervention |
Δpost-pre |
|
Control |
88.36 ± 2.29 |
89.51 ± 2.27 |
1.28 ± 0.49d |
65.37 ± 3.54 |
66.43 ± 3.10 |
1.59 ± 0.72d |
23.07 ± 1.49 |
27.09 ± 1.82 |
4.02 ± 1.90d |
82.77 ± 1.47 |
81.75 ± 1.66 |
−1.24 ± 0.40c |
HFD |
235.62 ± 4.33 |
237.28 ± 5.15 |
0.70 ± 1.00d |
140.75 ± 4.59 |
142.02 ± 4.64 |
0.89 ± 0.58d |
81.20 ± 1.49 |
83.04 ± 2.44 |
1.84 ± 0.96d |
25.51 ± 1.27 |
24.45 ± 1.37 |
−4.33 ± 0.45c |
WR |
233.45 ± 3.28 |
212.84 ± 4.02 |
−9.68 ± 5.24c |
140.40 ± 1.59 |
125.60 ± 2.41 |
−11.79 ± 2.52c |
81.43 ± 1.79 |
76.21 ± 1.75 |
−6.85 ± 1.84c |
28.00 ± 1.17 |
28.83 ± 1.37 |
2.87 ± 2.03c |
NGBR |
232.42 ± 3.72 |
105.43 ± 3.91 |
−120.45 ± 4.89b |
138.87 ± 2.31 |
86.31 ± 3.41 |
−60.90 ± 5.47b |
81.78 ± 2.60 |
42.17 ± 3.65 |
−93.93 ± 3.80b |
27.89 ± 2.06 |
71.65 ± 1.69 |
61.08 ± 2.22b |
GBR |
233.79 ± 2.73 |
99.63 ± 3.40 |
−134.66 ± 4.26a |
140.99 ± 1.32 |
77.14 ± 3.29 |
−82.76 ± 3.27a |
83.04 ± 1.80 |
35.92 ± 2.88 |
−131.16 ± 3.30a |
29.14 ± 1.58 |
79.93 ± 1.37 |
63.54 ± 1.74a |
Standard AIN-93M (Control), high-fat diet (HFD), high-fat diet + white rice (WR), high-fat diet + non-germinated black rice (NGBR), and high-fat diet + germinated black rice (GBR). Data are presented as mean ± SD (n = 6). Different letters in the same column indicate statistically significant differences among samples (p < 0.05).
Cecal cholesterol excretion
The analysis of cecal cholesterol aims to assess the proportion of dietary cholesterol and bile secreted cholesterol (endogenous) that is not reabsorbed in the intestine and is subsequently excreted in the feces. This parameter is important because it reflects decreased intestinal cholesterol absorption and increased conversion of cholesterol to bile acids. In general, cecal cholesterol levels show a pattern opposite to plasma cholesterol levels. In this study, cecal cholesterol levels in rats showed significant differences between treatment groups (Figure 3). The HFD group had the lowest cecal cholesterol levels at 23.83 mg/100 g. In contrast, the control group had the highest at 96.46 mg/100 g. Interestingly, intervention with GBR resulted in higher cecal cholesterol levels than with NGBR, WR, or HFD, indicating increased cholesterol excretion.
This cholesterol-lowering effect is thought to be influenced by the increased insoluble dietary fiber content in the GBR group. This mechanism is likely to act indirectly by triggering early satiation and sustained satiety [33]. In addition, the observed hypocholesterolemic effect is likely reinforced by the presence of soluble dietary fiber, which synergizes with insoluble dietary fiber by binding bile acids in the intestinal lumen, thereby inhibiting cholesterol solubility and absorption at the brush border membrane of intestinal epithelial cells. Previous reports also showed that bile acid excretion in feces increases in dietary fiber groups and is negatively correlated with serum cholesterol levels. This mechanism is further supported by evidence that dietary fiber increases mass in the gastrointestinal tract and can bind cholesterol, thereby reducing its absorption in the liver [34]. This condition promotes increased excretion of cholesterol along with lipids and bile acids through feces. The accumulation of cholesterol in the cecum reflects this process, as the cecum is the primary storage site before excretion, and can therefore be used as an indicator of increased cholesterol excretion due to fiber consumption.
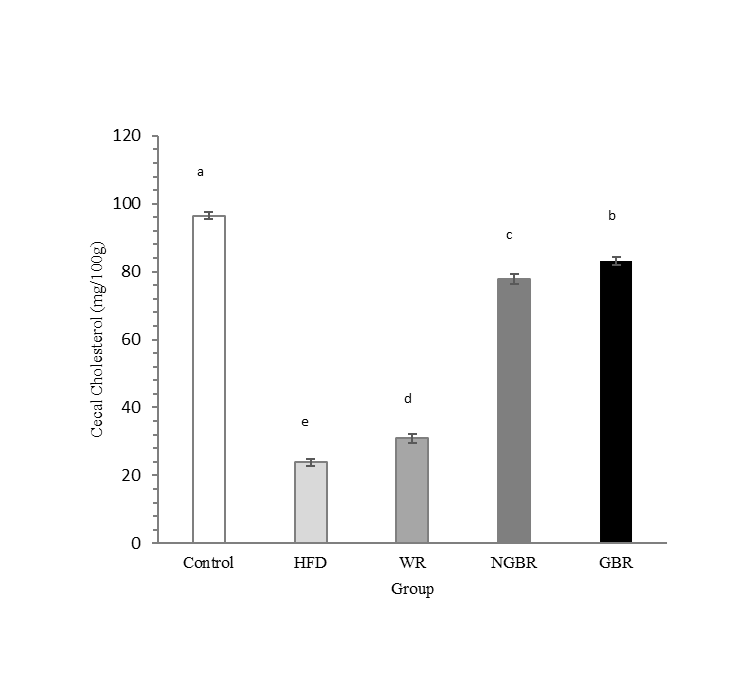
Figure 3 Effect of germinated black rice administration on cecal cholesterol excretion during the intervention. Standard AIN-93M (Control), high-fat diet (HFD), high-fat diet + white rice (WR), high-fat diet + non-germinated black rice (NGBR), and high-fat diet + germinated black rice (GBR). Data are presented as mean ± SD (n = 6). Different letters in the same column indicate statistically significant differences among samples (p < 0.05).
Plasma glucose level
Plasma glucose analysis was performed because obesity is consistently associated with insulin resistance through various mechanisms, including the accumulation of free fatty acids and dysregulation of adipokines, such as adiponectin and TNF-α, which underlie glucose metabolism disorders [35]. As shown in Figure 4, the analysis results indicate differences in the patterns of plasma glucose level changes between groups after 4 weeks of intervention. The control and HFD groups showed a slight increase from baseline to the end (Δ −1 - 2 mg/dL), while the WR group decreased slightly (Δ −3.6 mg/dL). A much greater decrease occurred in NGBR (Δ +41.6 mg/dL), with the most significant reduction observed in GBR (Δ +49.0 mg/dL) (p < 0.05).
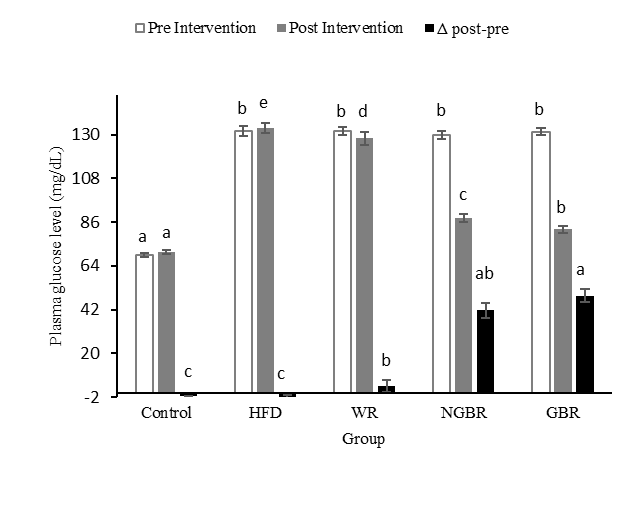
Figure 4 Effect of germinated black rice administration on plasma glucose levels in obese rats during the experiment. Standard AIN-93M (Control), high-fat diet (HFD), high-fat diet + white rice (WR), high-fat diet + non-germinated black rice (NGBR), and high-fat diet + germinated black rice (GBR). Data are presented as mean ± SD (n = 6). Different letters in the same column indicate statistically significant differences among samples (p < 0.05).
The most significant decrease in blood glucose levels was observed in obese rats treated with germinated black rice. This is most likely due to the higher fiber content in germinated Cempo Ireng black rice (7.43%) compared to non-germinated black rice (6.66%). Fiber has a hypoglycemic effect by slowing gastric emptying, inhibiting glucose diffusion, altering gastric peristalsis, reducing amylase activity due to increased intestinal viscosity, and shortening intestinal transit time, thereby decreasing glucose absorption [36]. This condition ultimately affects insulin secretion and glucose utilization in liver cells. In addition, pigmented rice contains bioactive compounds, including anthocyanins, a flavonoid group that has potential for medicinal and nutraceutical ingredients [37]. Several in vitro, in vivo, and clinical studies have demonstrated that the antidiabetic effects of black rice are strongly associated with its anthocyanin content. Anthocyanins, when delivered through fortified foods at effective doses, have been shown to inhibit carbohydrate-digesting enzymes in vitro and subsequently improve postprandial glycemic and lipidemic responses in vivo. These benefits include enhanced insulin sensitivity, reduced glucose absorption in the small intestine, and improved metabolic regulation observed in controlled human trials [27].
Malondialdehyde (MDA) level
Malondialdehyde is the main product of polyunsaturated fatty acid peroxidation and is widely used as an indicator of oxidative stress levels, both in vivo and in vitro. The accumulation of lipid peroxidation products increases with a high-fat diet [38]. In this study, MDA levels were measured from liver tissue after intervention. The results of the analysis showed that the HFD group had the highest MDA levels at 12.17 nmol/mL, while the control group had 2.12 nmol/mL, and the GBR group had 3.10 nmol/mL, showing the lowest levels (p < 0.05) compared to the NGBR and WR groups (Figure 5).
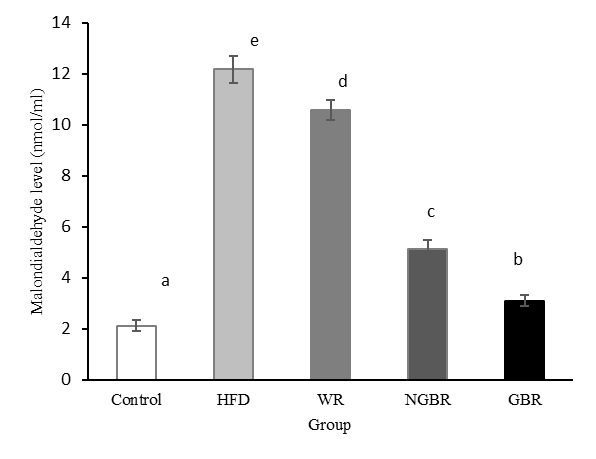
Figure 5 Effect of germinated black rice administration on malondialdehyde level in obese rats during the intervention. Standard AIN-93M (Control), high-fat diet (HFD), high-fat diet + white rice (WR), high-fat diet + non-germinated black rice (NGBR), and high-fat diet + germinated black rice (GBR). Data are presented as mean ± SD (n = 6). Different letters in the same column indicate statistically significant differences among samples (p < 0.05).
Similar findings have been reported, showing that supplementation with germinated brown rice in hypercholesterolemic rats significantly reduced serum MDA levels compared to non-germinated brown rice [22]. In the present study, obesity treatment with germinated black rice also reduced hepatic lipid peroxides. Hyperlipidemic conditions, particularly hypercholesterolemia, can lead to the formation of reactive oxygen species (ROS) via lipid oxidation, as evidenced by elevated malondialdehyde, the primary product of lipid peroxidation. The accumulation of ROS in the body triggers inflammatory responses, especially in the liver. Lipid oxidation can be prevented by hydrogen atom donors that react with lipid radicals to produce stable non-radical compounds. Molecules that can donate hydrogen atoms without becoming radicals themselves are antioxidants.
Short-Chain Fatty Acid (SCFA)
Short-chain fatty acids (SCFAs), such as acetate, propionate, and butyrate, are the primary products of dietary fiber fermentation by gut microbiota. These 3 fatty acids are the main SCFA components in the colon. SCFA concentrations are greatly influenced by the composition of the microbiota, dietary patterns, and the availability of fermentable substrates, especially dietary fiber. Consuming fiber-rich foods increases the availability of fermentable substrates for gut microbes, thereby boosting SCFA production. It is known that the germination process in Cempo Ireng black rice can increase dietary fiber content. This increase in dietary fiber content has the potential to improve the short-chain fatty acid (SCFA) profile in obesity.
The results of this study show a significant difference in total SCFA concentration between groups (p < 0.05). The control group had the highest total SCFA levels at 123.3 mmol, while the HFD group had the lowest at 49.6 mmol. GBR administration increased total SCFA to 80.3 mmol, which was higher than NGBR (75.8 mmol), WR (63.9 mmol), and HFD. For all SCFA types, the control group had the highest levels and differed significantly from the HFD group. GBR increased acetate levels compared to HFD, although it did not differ considerably from NGBR. In propionate, GBR was also higher than HFD (p = 0.004) but did not differ from WR and NGBR. For butyrate, the GBR group showed a significant increase compared to HFD (p = 0.004), although it did not reach the level of the control group (Figure 6).
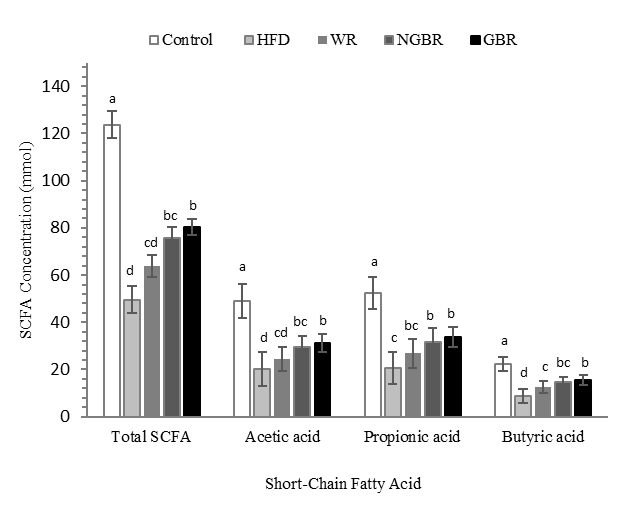
Figure 6 Effect of germinated black rice administration on short-chain fatty acid concentration in obese rats during the intervention. Standard AIN-93M (Control), high-fat diet (HFD), high-fat diet + white rice (WR), high-fat diet + non-germinated black rice (NGBR), and high-fat diet + germinated black rice (GBR). Data are presented as mean ± SD (n = 6). Different letters in the same column indicate statistically significant differences among samples (p < 0.05).
These results indicate that germinated black rice intervention can improve SCFA production in obesity. Fermentability of dietary fiber varies greatly. Soluble dietary fiber is generally almost completely fermentable in the colon, producing short-chain fatty acids (SCFA) that play a role in maintaining gut health, including providing anti-inflammatory effects and maintaining epithelial barrier integrity [39]. In contrast, insoluble dietary fiber undergoes fermentation to a lesser extent and at a slower rate. Acetate, propionate, and butyrate are the most abundant SCFAs, and they are present in the colon in a ratio of approximately 60:20:20, respectively [40]. The increase in total SCFA levels observed in the GBR group indicates that the dietary fiber contained in germinated black rice undergoes active fermentation in the colon. This shows an increase in the activity of microbes that produce SCFA, which, in turn, improves metabolic function.
Mechanistically, SCFAs play an essential role in metabolic regulation. Propionate in humans has been shown to increase the secretion of anorexigenic hormones such as peptide YY (PYY) and glucagon-like peptide-1 (GLP-1). These 2 hormones play a role in regulating appetite, increasing satiety, and reducing energy intake and weight gain [41]. Thus, the increase in propionate levels observed in the GBR group can be linked to the effects of suppressing energy intake and improving the body's energy balance by stimulating these intestinal hormones.
Histopathological analysis
Histopathological analysis of the liver (Figure 7) revealed differences in the degree of fatty liver between groups, as assessed by both semi-quantitative scores and area measurements using ImageJ. The control group showed almost no fatty liver, with a score of 0.6 ± 1.15 (area 23.02%). In the HFD group, severe fatty liver was observed, with a score of 3.0 ± 0.00 (area 31.23%). The WR treatment still showed moderate fatty liver, with a score of 2.0 ± 1.00 (area: 29.80%). Meanwhile, the NGBR treatment reduced the degree of fatty liver to mild, with a score of 1.5 ± 0.57 (area 22.08%). The most noticeable effect was observed in the GBR group, with a score of 1.0 ± 0.00 (area 13.54%), indicating the lowest degree of fatty liver among the treatment groups (Table 5). White adipose tissue in the HFD and WR groups showed adipocytes with larger sizes. Meanwhile, in the NGBR treatment group, adipocyte size appeared slightly reduced compared to the HFD group. In the GBR group, adiposity decreased significantly and approached levels similar to those of the control group (Figure 8).
In the HFD and WR intervention groups, swollen hepatocytes persisted, whereas the NGBR and GBR interventions showed improvements in hepatocyte morphology and adiposity. The histopathological results showed similar findings, namely a reduction in micro- and macrovesicular steatosis in obese rats treated with 100% GBR [23]. This is consistent with another report stating that germinated brown rice has the potential to reduce fat accumulation by inhibiting adipocyte differentiation and stimulating lipolysis in adipocytes, as shown in in vitro studies [42].
Table 5 Semi-quantitative visual scoring and quantitative image analysis of hepatic steatosis in rats.
Group |
Visual score (Mean ± SD) |
Visual description |
Steatosis area (%) |
Control |
0.6 ± 1.15 |
Non to very mild |
23.02b |
HFD |
3.0 ± 0.00 |
Severe |
31.23c |
WR |
2.0 ± 1.00 |
Moderate |
29.80c |
NGBR |
1.6 ± 0.57 |
Mild |
22.08b |
GBR |
1.0 ± 0.00 |
Mild |
13.54a |
Representative data of hepatic steatosis evaluation in liver sections. Semi-quantitative scoring was performed by visual histopathological assessment of Hematoxylin and Eosin-stained sections (0 = none; 1 = mild; 2 = moderate; 3 = severe), while quantitative analysis was conducted by measuring the steatosis area (%) using ImageJ software. Values are presented as mean ± SD (n = 3).
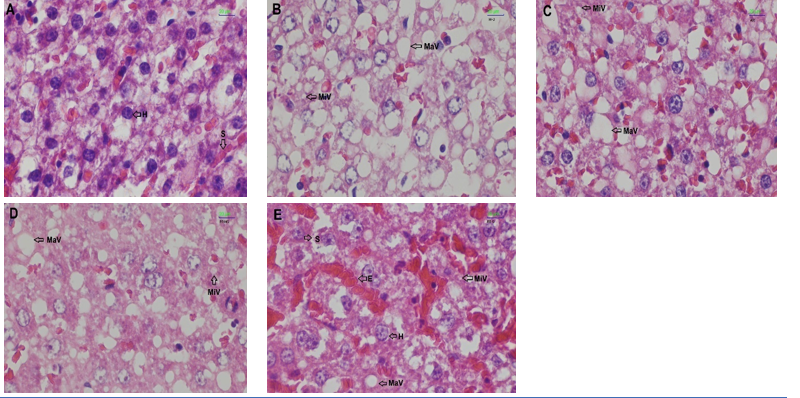
Figure 7 Liver photomicrographs (H&E, 4×) of rats: (A) Control, (B) HFD, (C) WR, (D) NGBR, and (E) GBR groups. H, Hepatocytes; MaV, Macrovesicular; MiV, Microvesicular; E, Erythrocytes; S, Sinusoid.
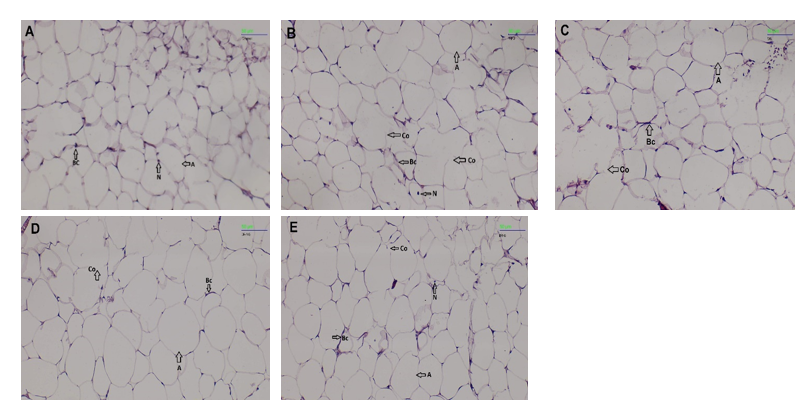
Figure 8 White adipose tissue photomicrographs (H&E, 10×) of rats: (A) Control, (B) HFD, (C) WR, (D) NGBR, and (E) GBR groups. A, Adipocytes; Bc, Blood capillaries; Co, Coalescence; N, Nucleus.
The effect of germinated black rice administration on lipid metabolism–related gene expression
The effect of germinated black rice administration on mRNA levels was analyzed using qRT-PCR in the liver to evaluate the expression of genes involved in lipid metabolism, namely Fasn (Fatty acid synthase), Hmgcr (3-Hydroxy-3-methylglutaryl-CoA Reductase), and Lxr-α (Liver X receptor alpha). Expression levels were normalized to the housekeeping gene Eukaryotic translation elongation factor 1α1 (Eef1α1) and calculated using the 2^-ΔΔCt method. The data were normalized to the HFD group, which was used as the baseline for comparison with the GBR group.
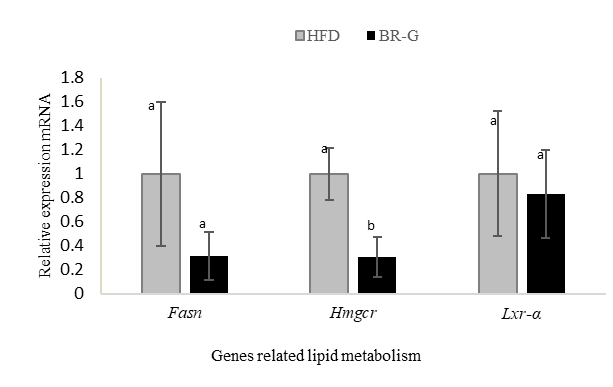
Figure 9 Effect of germinated black rice administration on gene expression. High-fat diet (HFD), high-fat diet + germinated black rice (GBR). Data are presented as mean ± SD (n = 3). Different letters in the same column indicate statistically significant differences among samples (p < 0.05).
Based on Figure 9, among the 3 genes analyzed, Hmgcr showed a statistically significant difference, with a very strong effect size (Cohen’s d = 3.61), indicating that germinated black rice has a strong regulatory effect on cholesterol synthesis. In contrast, Fasn showed a strong Cohen’s d effect value at 1.525 but was not statistically significant, suggesting that it may still have a biological tendency even though it is not yet significant. Meanwhile, Lxr-α showed a small Cohen's d effect value at 0.376 and was not significantly different, indicating that the effect of germinated black rice on lipid regulation via Lxr-α is relatively limited. Overall, these results suggest that germinated black rice (GBR) exerts a stronger effect on the cholesterol synthesis pathway than on the lipogenesis pathway.
These results are consistent with findings that feeding pre-germinated brown rice (PGBR) to HFD rats reduced HMGCR protein levels by approximately 78% [43]. Thus, although this study is still limited to the mRNA level, these findings reinforce the possibility that the decrease in Hmgcr expression induced by GBR may extend to the protein and enzymatic activity levels, which need to be confirmed in further research. HMG-CoA reductase is a key enzyme in the cholesterol biosynthesis pathway (mevalonate pathway) that converts HMG-CoA to mevalonate, an essential step in cholesterol formation. HMGCR regulation can occur at several levels, namely, SREBP-2-mediated transcription downregulation, mRNA stability downregulation, and post-translational regulation, such as phosphorylation and ubiquitination, which accelerate protein degradation [44]. Therefore, the observed changes in Hmgcr mRNA expression likely reflect one or a combination of these mechanisms, which imply a reduction in intrahepatic cholesterol synthesis.
In line with this mechanism, the increase in SCFA levels, especially propionate, observed in the GBR group indicates that the effect of decreased Hmgcr gene expression is also likely influenced by propionate's suppression of cholesterol biosynthesis in the liver. This effect may occur by inhibiting HMG-CoA reductase and acetyl-CoA reductase, thereby altering transcriptional regulation via SCFA-mediated signaling, particularly propionate [45]. These findings are consistent with previous reports that propionate can modulate various critical metabolic processes, including gluconeogenesis, ureogenesis, and ketogenesis, further emphasizing its multifunctional role in maintaining energy and lipid homeostasis [46]. However, the gene expression findings were based solely on mRNA levels and were not validated by Western blot, limiting the strength of molecular interpretation.
Conclusions
This study shows that substitution with germinated black rice Cempo Ireng variety protects against obesity in rats fed a high-fat diet, compared with non-germinated black rice and white rice. The 48 h germination process has been shown to increase the content of bioactive compounds, particularly dietary fiber, thereby improving lipid and glucose metabolism. These effects are demonstrated by reduced liver and adipose tissue weight, improved lipid profile (decreased plasma triglycerides, total cholesterol, and LDL cholesterol), increased HDL cholesterol and cecal cholesterol excretion, and reduced plasma glucose and malondialdehyde levels. These benefits are mediated by the hypoglycemic and hypolipidemic effects of dietary fiber, modulation of gut microbiota and short-chain fatty acid production, and antioxidant properties associated with genetic regulation. Overall, these findings indicate that germinated black rice has the potential to be developed as a functional food in obesity diet management.
Acknowledgements
This research was funded by the Indonesian Endowment Fund for Education (LPDP), Ministry of Finance of the Republic of Indonesia, awarded to Mina Parhatus Salamah under scholarship ID 202312110309909. The authors gratefully acknowledge the facilities provided by the Laboratory of Food and Nutrition, Universitas Gadjah Mada, Indonesia and the Laboratory of Nutrition, Tohoku University, Japan.
Declaration of Generative AI in Scientific Writing
The authors employed generative AI tools, including ChatGPT (OpenAI, USA) and Perplexity, to support language refinement, improve clarity, and assist with grammar correction during manuscript preparation. These tools were not used to generate original scientific content, interpret results, or perform data analysis. All AI-assisted outputs were carefully reviewed, edited, and validated by the authors, who take full responsibility for the accuracy, integrity, and conclusions of the final manuscript.
CRediT Author Statement
Mina Parhatus Salamah: Methodology, formal Analysis, statistical analysis, visualization, writing-review, editing; Yunika Mayangsari: Supervision, methodology, writing-review, editing; Hitoshi Shirakawa: Methodology, writing-review, editing; Afifah Zahra Agista: Methodology, formal Analysis; Wahyu Dwi Saputra: Supervision, conceptualization, curation, methodology, visualization, writing-review, editing. All authors read and approved the final manuscript.
Reference
[1] JS Garrow. Obesity and related disease. Churchill Livingstone, Edinburgh, 1988.
[2] AH Mokdad, BA Bowman, ES Ford, F Vinicor, JS Marks and JP Koplan. The continuing epidemics of obesity and diabetes in the United States. Journal of the American Medical Association 2001; 286(10), 1195-1200.
[3] PA Stapleton, ME James, AG Goodwill and JC Frisbee. Obesity and vascular dysfunction. Pathophysiology 2008; 15(2), 79-89.
[4] AK Singh, SK Singh, N Singh, N Agrawal and K Gopal. Obesity and dyslipidemia 2011, Available at: https://www.biomedscidirect.com, accessed August 2025.
[5] WHO. Obesity and overweight 2025, Available: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight, accessed December 2025.
[6] N Aini, H Dwiyanti and MP Salamah. Antioxidants and in vitro starch digestibility of coloured rice, and its effect on blood sugar and malonaldehyde in streptozotocin-nicotinamide-induced diabetic rats. International Food Research Journal 2023; 30(3), 723-735.
[7] S Singh, M Habib, D Mondal, M Thakur, Y Kumar, K Bashir, S Jan and K Jan. Effects of germination on the physicochemical, thermal, in vitro protein digestibility, Antinutrients, and antioxidant properties of sorghum flour. International Journal Food Science Technology 2024; 59(9), 6513-6521.
[8] H Jin, H Han, G Song, HJ Oh and BY Lee. Anti-obesity effects of GABA in C57BL/6J mice with high-fat diet-induced obesity and 3T3-L1 adipocytes. International Journal of Molecular Sciences 2024; 25(2), 995.
[9] IO Owolabi, K Chakree and CT Yupanqui. Bioactive components, antioxidative and anti-inflammatory properties (on RAW 264.7 macrophage cells) of soaked and germinated purple rice extracts. International Journal Food Science Technology 2019; 54(7), 2374-2386.
[10] L Chen, J Kan, N Zheng, B Li, Y Hong, J Yan, X Tao, G Wu, J Ma, W Zhu, L Sheng, L Chen, B Li, J Zhong, J Du and H Li. A botanical dietary supplement from white peony and licorice attenuates nonalcoholic fatty liver disease by modulating gut microbiota and reducing inflammation. Phytomedicine 2021; 91, 153693.
[11] Y Gu, Y Zhang, M Li, Z Huang, J Jiang, Y Chen, J Chen, Y Jia, L Zhang and F Zhou. Ferulic acid ameliorates atherosclerotic injury by modulating gut microbiota and lipid metabolism. Frontiers in Pharmacology 2021; 25(12), 621339.
[12] A Basith, S Noer and M Faizah. Variation in anthocyanin content level in 4 local varieties of black rice (Oryza sativa L.) from Indonesia. Jurnal Pertanian 2023; 14(1), 1-6.
[13] AH Arifa, E Syamsir and S Budijanto. Physicochemical properties of black rice (Oryza sativa L.) from West Jawa, Indonesia. AgriTECH 2021; 41(1), 15-24.
[14] H Munarko, AB Sitanggang, F Kusnandar and S Budijanto. Germination of 5 Indonesian brown rice: Evaluation of antioxidant, bioactivcompounds, fatty acids and pasting properties. Food Science and Technology 2022; 42, 19721.
[15] JT Dourmashkin, GQ Chang, EC Gayles, JO Hill, SK Fried, C Juliene and SF Leibowitz. Different forms of obesity as a function of diet composition. International Journal of Obesity 2005; 29(11), 1268-1278.
[16] EL Simson and RM Gold. The lee obesity index vindicated? Physiology & Behavior 1982; 29(2), 371-376.
[17] PG Reeves, FH Nielsen and GC Fahey. AIN-93 purified diets for laboratory rodents: Final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. Journal of Nutrition 1993; 123(11), 1939-1951.
[18] B Hong, S Zhang, D Yuan, S Shan, J Zhang, D Sha, D Chen, W Yin, S Lu and C Ren. Changes in bioactive constituents in black rice metabolites under different processing treatments. Foods 2025; 14(9), 1630.
[19] MA Martín-Cabrejas, N Ariza, R Esteban, E Mollá, K Waldron and FJ López-Andréu. Effect of germination on the carbohydrate composition of the dietary fiber of peas (Pisum sativum L.). Journal of Agricultural and Food Chemistry 2003; 51(5), 1254-1259.
[20] S Lekjing and K Venkatachalam. Effects of germination time and kilning temperature on the malting characteristics, biochemical and structural properties of HomChaiya rice. RSC Advances 2020; 10(28), 16254-16265.
[21] H Nonogaki, GW Bassel and JD Bewley. Germination-still a mystery. Plant Science 2010; 179(6), 574-581.
[22] SA Mantiri, Y Mayangsari, Ardiansyah, H Shirakawa and WD Saputra. In vivo evaluation of germinated IR-64 brown rice dietary administration on the lipid blood profiles of rats with hypercholesterolemia. Nutrire 2024; 49(2), 44.
[23] SM Lim, YM Goh, N Mohtarrudin and SP Loh. Germinated brown rice ameliorates obesity in high-fat diet induced obese rats. BMC Complementary Medicine and Therapies 2016; 16(1), 140.
[24] LM Guardianelli, MV Salinas and MC Puppo. Chemical and thermal properties of flours from germinated amaranth seeds. Journal of Food Measurement and Characterization 2019; 13, 1078-1088.
[25] CB Ioniță-Mîndrican, K Ziani, M Mititelu, E Oprea, S Neacșu, E Moroșan, D Dumitrescu, A Roșca, D Drăgănescu and C Negrei. Therapeutic benefits and dietary restrictions of fiber intake: A state of the art review. Nutrients 2022; 14(13), 2641.
[26] C Ren, S Lu, S Shan, S Zhang, B Hong, D Yuan, J Zhang, S Gao, Q Liu and X Fan. Nutrient-Enriched germinated brown rice alters the intestinal ecological network by Regulating Lipid Metabolism in Rats. International Journal of Molecular Sciences 2025; 26(16), 7693.
[27] SJL Ou, D Yang, HP Pranata, ES Tai and MH Liu. Postprandial glycemic and lipidemic effects of black rice anthocyanin extract fortification in foods of varying macronutrient compositions and matrices. npj Science of Food 2023; 7, 59.
[28] MO Lee. Determination of the surface area of the white rat with its application to the expression of metabolic results. American Journal of Physiology-Legacy Content 1929; 89(1), 24-33
[29] C Huang, X Gao, Y Shi, L Guo, C Zhou, N Li, W Chen, F Yang, G Li, Y Zhuang, P Liu, G Hu and X Guo. Inhibition of hepatic AMPK pathway contributes to free fatty acids-induced fatty liver disease in laying hen. Metabolites 2022; 12(9), 825.
[30] MW Furnes, CM Zhao and D Chen. Development of obesity is associated with increased calories per meal rather than per day. A study of high-fat diet-induced obesity in young rats. Obesity Surgery 2009; 19(10), 1430-1438.
[31] MW Zhang, RF Zhang, FX Zhang and RH Liu. Phenolic profiles and antioxidant activity of black rice bran of different commercially available varieties. Journal of Agricultural and Food Chemistry 2010; 58(13), 7580-7587.
[32] TS Kahlon and FI Chow. In vitro binding of bile acids by rice bran, oat bran, wheat bran, and corn bran. Cereal Chemistry 2000; 77(4), 518-521.
[33] B Burton-Freeman. Dietary fiber and energy regulation. Journal of Nutrition 2000; 130(2), 272S-275S.
[34] N Sun, L Tong, T Liang, L Wang, L Liu, X Zhou and S Zhou. Effect of oat and tartary buckwheat - based food on cholesterol – lowering and gut microbiota in hypercholesterolemic hamsters. Journal of Oleo Science 2019; 68(3), 251-259.
[35] M Dewi. Resistensi insulin terkait obesitas: Mekanisme endokrin dan intrinsik sel. Jurnal Gizi dan Pangan 2007; 2(2), 49-54.
[36] EB Giuntini, FAH Sardá and EW de Menezes. The effects of soluble dietary fibers on glycemic response: An overview and futures perspectives. Foods 2022; 11(23), 3934.
[37] FM Bhat, SR Sommano, CS Riar, P Seesuriyachan, T Chaiyaso and C Prom-U-Thai. Status of bioactive compounds from bran of pigmented traditional rice varieties and their scope in production of medicinal food with nutraceutical importance. Agronomy 2020; 10(11), 1817.
[38] J Wang, Y Jiang, L Jin, C Qian, W Zuo, J Lin, L Xie, B Jin, Y Zhao, L Huang and Y Wang. Alantolactone attenuates high-fat diet-induced inflammation and oxidative stress in non-alcoholic fatty liver disease. Nutrition & Diabetes 2024; 14(1), 41.
[39] D Mudgil and S Barak. Composition, properties and health benefits of indigestible carbohydrate polymers as dietary fiber: A review. International Journal of Biological Macromolecules 2013; 61, 1-6.
[40] C Martin-Gallausiaux, L Marinelli, HM Blottière, P Larraufie and N Lapaque. SCFA: Mechanisms and functional importance in the gut. Proceedings of the Nutrition Society 2021; 80(1), 37-49.
[41] JJ Lim, Y Liu, LW Lu, IR Sequeira and SD Poppitt. No evidence that circulating GLP-1 or PYY are associated with increased satiety during low energy diet-induced weight loss: modelling biomarkers of appetite. Nutrients 2023; 15(10), 2399.
[42] SM Lim, YM. Goh, WB Kuan and SP Loh. Effect of germinated brown rice extracts on pancreatic lipase, adipogenesis and lipolysis in 3T3-L1 adipocytes. Lipids in Health and Disease 2014; 13(1), 169.
[43] K Shen, C Hao, H Yen, C Chen, J Chen, F Chen and H Lin. Pre-germinated brown rice prevented high fat diet induced hyperlipidemia through ameliorating lipid synthesis and metabolism in C57BL/6J mice. Journal of Clinical Biochemistry and Nutrition 2016; 59(1), 39-44.
[44] PR Clarke and DG Hardie. Regulation of HMG-CoA reductase: Identification of the site phosphorylated by the AMP-activated protein kinase in vitro and in intact rat liver. EMBO Journal 1990; 9(8), 2439-2446.
[45] SM Lim, YM Goh, W Bin Kuan and SP Loh. Effect of germinated brown rice extracts on pancreatic lipase, adipogenesis and lipolysis in 3T3-L1 adipocytes. Lipids in Health and Disease 2014; 13(1), 169.
[46] C Remesy and C Demigne. Determination of volatile fatty acids in plasma after ethanolic extraction. Biochemical Journal 1974; 141(1), 85-91.