Trends
Sci.
2026; 23(8): 13125
Salinity-Dependent Modulation of Antioxidant Defense, Apoptosis Enzymes, and Molecular Complexes Interaction in Tilapia Exposed to Polystyrene Nanoplastics
Muhammad
Nuh Fathsyah Siregar1,
![]() ,
Alfiah Hayati1,2,*,
,
Alfiah Hayati1,2,*,
![]() ,
Muhammad Iqbal1,
,
Muhammad Iqbal1,
![]() ,
,
Mochammad
Aqilah Herdiansyah1,
![]() ,
Manikya Pramudya1,
,
Manikya Pramudya1,
![]() ,
,
Firli
Rahmah Primula Dewi1,
![]() ,
Aunurohim3,
,
Aunurohim3,
![]() ,
Hisyam Rizky Saputra1,
,
Hisyam Rizky Saputra1,
![]()
and
Febriyansyah Saputra1,
![]()
1Department of Biology, Faculty of Science and Technology, University of Airlangga, East Java, Indonesia
2Developmental Biology & Biomedical Science Group, University of Airlangga, East Java, Indonesia
3Department of Biology, Faculty Science and Data Analitics, Institut Teknologi Sepuluh Nopember,
Jawa Timur, Indonesia
(*Corresponding author’s e-mail: [email protected])
Received: 23 December 2025, Revised: 5 February 2026, Accepted: 12 February 2026, Published: 20 March 2026
Abstract
Nanoplastics (NPs) are emerging contaminants due to their persistence, bioaccumulation, and small particle size. These pollutants are derived from the breakdown of larger plastic debris and frequently enter aquatic environments through runoff from unmanaged landfills and illegal dumping sites. Based on the habitat, fish could be found in some ecosystem actually, such as in fresh water, breakish water, and marine water. Moreover, NPs can affect the physiology and biochemistry of fish, potentially altering oxidative balance and triggering apoptosis. The information about the effects of NP exposure on fish in different salinity level was still limited, therefore this study aimed to examine the effects of polystyrene nanoplastics (PS-NPs) on antioxidant activity and apoptosis-inducing enzymes in tilapia’s strain Jatimbulan (Oreochromis niloticus) under varying salinity levels. Tilapia were divided into control groups (without NPs) and treatment groups exposed to PS-NPs (2 µL/kg) at salinities of 0, 2.5, 5, 10, and 15 ppt for 25 days. Antioxidant enzymes (catalase (CAT) and superoxide dismutase (SOD)) and apoptosis-related enzymes (caspase-3 and caspase-9) were quantified using ELISA test. Molecular docking was conducted using the styrene monomer (CID: 7501) as the ligand, and CAT, SOD, caspase-3, and caspase-9 as target receptors. Docking analysis and validation revealed that exposure of PS-NPs significantly reduced antioxidant levels and increased activity of apoptosis-related enzymes, particularly in fish reared at higher salinities. The results showed that exposure to PS-NPs on 0 ppt significantly reduced concentration of antioxidant enzymes and increased apoptosis-related enzymes levels. On different salinity level, it showed that salinity modulates NP toxicity by influencing antioxidant enzymes and apoptosis-related enzymes. Moreover, the molecular docking experiment revealed that CAT (−5.3 kcal/mol) and caspase-9 (−4.6 kcal/mol) proteins were found to be most affected by the presence of PS-NPs in cells, as evidenced by highest binding affinity compared to other test proteins. Molecular docking indicates direct interactions of styrene monomers with catalase and caspase-9, suggesting disruption of oxidative stress regulation and apoptosis pathways. In conclusion, this study highlighted the risk of NP pollution in different salinity level, representative of some aquatic ecosystems, persuading to be care and find mitigation solution to reduce the impact of NP pollution, such as remediation.
Keywords: Nanoplastics, Fish, Health, Apoptosis, Molecular docking
Introduction
The widespread presence of plastic waste in aquatic environments has developed into a significant ecological problem, especially when it degrades into extremely small molecules known as NPs. The emergence of NP particles smaller than 100 nm poses substantial risks to the health of aquatic biota [1]. NP particles originate from the degradation of larger plastic fragments (microplastics), which are generally produced as a result of improper plastic waste disposal or mismanagement. Through surface runoff and river flow, NPs are carried from terrestrial sources into freshwater bodies and eventually discharged into marine ecosystems [2]. Their very small size and persistent nature facilitate wide distribution and potential for biological accumulation, thus posing serious threats to aquatic organisms and overall ecosystem health [3,4]. Among various types of nanoplastics, polystyrene nanoplastics (PS-NPs) are recognized as one of the most abundant and frequently detected forms in marine environments, owing to the extensive global use of polystyrene in packaging, food containers, and single-use plastic products [5]. The field study inform that fish could accumulate MPs and NPs in organs [6] . NPs also could uptake in zebrafish embryonic fibroblast cells (ZF4) [7,8], even NPs influenced on increasing MDA levels, which is indicator of rise of ROS [9]. While NPs’ effects on aquatic life are studied, the combined influence of salinity and NPs on freshwater species like Tilapia remains poorly understood.
Due to their extremely small size, NP particles are capable of penetrating biological membranes, interacting with, and accumulating within cells and tissues. One of the main mechanisms of NP toxicity is the induction of oxidative stress through the formation of reactive oxygen species (ROS). ROS are reactive molecules produced as byproducts of normal oxygen metabolism; however, their production can increase significantly upon exposure to pollutants, including NPs [10]. Oxidative stress caused by NPs leads to cellular damage, including lipid peroxidation and DNA damage, disrupting normal cell functions, as well as activating apoptosis pathways [11].
To maintain oxidative homeostasis, aquatic organisms such as tilapia possess antioxidant defense systems, both enzymatic and non-enzymatic. Two key
enzymes in this defense system are superoxide dismutase (SOD), which plays an important role in converting superoxide radicals into hydrogen peroxide, and catalase (CAT), which decomposes hydrogen peroxide into water and oxygen, thereby reducing potential oxidative damage. Previous study demonstrated that NP exposure can significantly increase ROS levels in fish , thereby disturbing enzyme activities [12]. Changes in SOD and CAT activities at advanced stages are indicative of the disruption of the antioxidant defense system in dealing with excessive oxidative stress [6]. This condition increases the likelihood of lipid peroxidation, indicated by elevated malondialdehyde levels, as well as DNA damage and activation of apoptosis pathways through caspases [10]. Overall, the increase in ROS due to NP exposure, followed by changes in SOD and CAT activities, serves as an early indicator of oxidative stress in aquatic organisms, which can progress into further cellular damage and threaten the survival and physiological functions of fish [9].
Moreover, ROS accumulation leads to damage of lipids, proteins, and DNA, ultimately activating apoptosis or programmed cell death pathways [11]. Apoptosis is an essential mechanism to maintain tissue homeostasis by eliminating damaged or abnormal cells. One of the main apoptosis pathways is the intrinsic (mitochondrial) pathway, which is triggered by severe oxidative stress, DNA damage, or disturbances in cellular redox balance [11]. Activation of this pathway involves the release of cytochrome c from mitochondria into the cytosol, which subsequently induces apoptosome formation and activates caspase-9. Caspase-9 then activates caspase-3, the main executioner caspase in apoptosis, which cleaves vital intracellular substrates, induces DNA fragmentation, and ultimately leads to cell death [13].
On the other hand, salinity is an important environmental factor that can modulate the behavior and toxicity of NPs in aquatic ecosystem. On varies salinity, the previous research have shown that fish in high-salinity water accumulate higher than in low salinity condition [14,15]. Variations in salinity affect NP aggregation, colloidal stability, and bioavailability. For instance, in high salinity environments, salt ions such as Na⁺ and Cl⁻ can suppress electrostatic repulsion between NP particles, promoting aggregation and modifying NP bioavailability, which in turn influences ROS levels and antioxidant responses [16]. This aggregation may increase the effective particle size and reduce solubility, but on the other hand, it also increases the likelihood of ingestion by filter-feeding or omnivorous fish [17]. Furthermore, high salinity can affect cell membrane permeability, thus increasing NP uptake into cells. Salinity is also known to affect plasma protein stability and ionic homeostasis in fish, exacerbating the toxic effects of NPs and increasing susceptibility to cellular damage [18].
Therefore, information on the interaction between NPs and salinity is crucial to understand the multifactorial conditions that exacerbate toxicity in aquatic biota, particularly fish with high ecological and economic value. A deep understanding of this relationship is essential to formulate ecosystem-based plastic pollution mitigation strategies that are adaptive to varying aquatic environmental conditions.
Materials and methods
Animal and ethical clearance
This study was conducted in accordance with international ethical standards for animal research, ensuring humane treatment and minimizing distress. All procedures including handling, NP exposure, sample collection, and euthanasia were carefully designed to adhere to ethical guidelines. Ethical approval for this research was granted by the Health Research Ethics Clearance Commission, Faculty of Dental Medicine, Universitas Airlangga (0885/HRECC.FODM/VIII/2024).
Anesthetic and euthanasia method
Fish were anesthetized with 0.1 mL/L clove oil, a natural sedative containing eugenol known for its effectiveness and low physiological impact [19]. Clove oil was emulsified before dilution in water for even distribution. Sedation was confirmed by reduced eye movement and loss of balance. Euthanasia was performed by immersion in 0.3 mL/L clove oil for at least 10 min until reflexes ceased [19]. All procedures adhered to ethical guidelines and were approved by the relevant research ethics committee.
Experimental design
Twenty-four male tilapia (Jatimbulan Strain, with size of each fish about 100 ± 10 g) were obtained from the UPT Umbulan freshwater aquaculture center in Pasuruan, East Java, Indonesia. Prior to experimentation, fish were quarantined for seven days in 100L aquarium containing 40L water, under controlled conditions (28 - 30 °C, pH 7 - 8, 12-hour light/dark cycle) at Universitas Airlangga. Each tank was equipped with a filtration system (Armada-Aquarium Top Filter, Color Aquatic Indonesia) and an aeration device (Amara-AA350, MTA-4362145-Indonesia) to maintain water quality. Fish were fed commercial pellets at 2% - 3% of body weight twice daily (8 AM and 4 PM) for 25 days. The amount of feed was carefully measured to ensure that there is no uneaten feed. To understand the effect of NPs under different salinity conditions, fish were gradually acclimated to 15 ppt over four days, with salinity increased from 2.5 ppt on first day, to 5 ppt on second day, 10 ppt on day third day, and 15 ppt on forth day. After acclimation, fish were randomly assigned to six groups with four replicates each. On NPs treatment in this study, fish were fed the contamintated-NPs pellets. The 100 nm PS-NPs (concentration 2 µL/kg; Sigma-Aldrich, USA) diluted (in 60 mL distilled water) was sprayed on 1.95 kg pellets, followed by 24-hour air drying. PS-NPs were thoroughly mixed into fish food and confirmed through spectroscopy to ensure uniform distribution before exposure [3].
Enzyme-Linked Immunosorbent Assay (ELISA) for antioxidants and apoptosis related enzymes
Liver samples were collected for the analysis of antioxidant activity and apoptosis-related enzymes after 25 days of PS-NP exposure. Liver samples were homogenized, centrifuged, and stored at −80 °C for further analysis of antioxidant and apoptosis-related enzyme levels using ELISA. Practically, the samples were centrifuged at 13,000 rpm for 10 min at 4 °C using an Eppendorf 5424R centrifuge, followed the last study on spleen [9]. The supernatant was carefully transferred into 1.5 mL microcentrifuge tubes and stored at −80 °C for further analysis. The levels of Caspase-3 and Caspase-9 were measured using a sandwich-ELISA kit, while the activities of CAT and SOD were assessed using competitive-ELISA kits (Bioassay Technology Laboratory, Shanghai, China). ELISA procedures were carried out according to the manufacturer’s instructions, and absorbance readings were taken using a microplate reader (Bio-Rad Model 680, USA). All samples and standards were analysed in triplicates to ensure accuracy and reliability.
ORP Measurement
Measurement of oxidation-reduction potential (ORP) in the liver tissue was conducted to assess the redox status as an indicator of oxidative stress. Approximately 1 g of liver tissue was homogenized in 9 mL of cold phosphate-buffered saline (PBS, pH 7.4) using a glass homogenizer. The homogenate was then filtered and the ORP value was measured using a calibrated ORP electrode (mV) connected to a digital multimeter. All measurements were performed at room temperature and conducted in triplicate to ensure accuracy and reproducibility [20].
Histological observation and quantification
Histological assessment was conducted using the standard haematoxylin and eosin (HE) staining method [3,9], which allows for the differentiation of cellular and tissue structures based on their affinity to the respective dyes. Haematoxylin selectively stains nuclear components blue-purple, while eosin counterstains cytoplasmic and extracellular matrix elements pink, providing optimal contrast for morphological evaluation. Liver Tissue samples were fixed in 10% buffered formalin, dehydrated through a graded series of alcohol, cleared in xylene, and embedded in paraffin. Sections approximately 5 µm in thickness were prepared using a microtome and mounted on glass slides, followed by staining with HE according to standardized protocols. Histological features were examined under a light microscope to assess tissue architecture, cellular integrity, and specific pathological alterations such as edema, necrosis, or inflammatory infiltration. For quantitative analysis, digital images of the stained sections were captured and analyzed using the ImageJ software (NIH, Bethesda, MD, USA). Parameters such as area, cellular density, or lesion scoring were measured by thresholding and segmentation tools available within the software, allowing for objective and reproducible quantification of histopathological findings [1].
Molecular docking materials
In this study, molecular docking experiments were conducted using Dell Vostro 143000 and Intel® AMD Ryzen 5 3500U models. The operating system used was Windows 10 Ultimate 64-bit with Radeon Vega MobilrGfx 2.10 GHz. The test compound used was styrene monomer from the polystyrene polymer complex (CID:7501). Meanwhile, the test receptors used were from the apoptosis-related enzyme group (caspase-3 and caspase-9) and endogenous antioxidants (SOD and CAT). The entry numbers used for each test receptor are as follows: Caspase-3 (UniProt ID: A0A669EPP7), caspase-9 (UniProt ID: A0A669DL02), SOD (UniProt ID: F5CT17), and CAT (UniProt ID: A0A669BAJ8).
Ligand and receptor preparation
The test compound preparation was performed using the PubChem webserver (https://pubchem.ncbi.nlm.nih.gov/). Meanwhile, the test receptor sequence was obtained from the UniProt webserver (https://www.uniprot.org/) with the specific organism Oreochromis niloticus. Next, protein modeling was performed using the SwissModel webserver (https://swissmodel.expasy.org/). The modeling process uses sequences copied from the UniProt web server. At the end of the protein modeling process, there will be several protein options with varying percent identities. The protein is selected based on the highest percent identity value and downloaded in PDB file format (.pdb).
Molecular docking experiment and visualization of complexes
Molecular docking experiments were conducted using PyRx 0.8 software (SourceForge Headquarters, San Diego, California, USA). This molecular docking method was used to explore the molecular interaction model between styrene monomers and endogenous antioxidant proteins and apoptosis-related enzymes. This method is often used to predict in advance the effects of styrene monomers on the affected test proteins based on their binding affinity values. The more negative the binding affinity value, the greater the binding ability between the ligand and the receptor [21]. This molecular model is important to know to ellaborate in vivo test results [22]. Furthermore, the chemical interaction visualization of molecular docking results was carried out using PyMol v1.74 software (Schrodinger, LLC, USA). Visualization of molecular docking results is performed to determine the type of binding in amino acid residues, whether it be Van der Walls bonds, hydrophobic interactions, or hydrogen interactions.
Validation of molecular docking experiment
Validation of molecular docking test was conducted using CABS-flex 3.0 webserver (https://Icbio.pl/cabsflex3) [23]. This method is used to determine the stability of styrene monomer complexes and test proteins. Parameter used in this simulation were protein rigidity, restraints, C-alpha restraints weight, side-chain restraint weight, trajectory, temperature range, number of cycles, and RNG seed.
Statistical analysis
Data were analysed using GraphPad Prism 10.3.1 software (GraphPad Software, USA). The normality of the data was first assessed using the Shapiro-Wilk test. For comparisons between multiple groups, one-way analysis of variance (ANOVA) followed by Tukey’s post-hoc test was performed to evaluate differences between experimental groups. Results were expressed as mean ± standard deviation (SD). A p-value of less than 0.05 was considered statistically significant.
Results and discussion
Effects of nanoplastics on antioxidants (CAT, and SOD)
The activity of the antioxidant CAT and SOD at Figure 1 showed a non-linear response to NPs exposure across different salinity levels. At 0 ppt (freshwater), NP exposure itself significantly decreased CAT and SOD levels compared to the control treatment (without NPs). On different salinity level, exposure NPs on lower salinity (0 - 5 ppt) increased the activities of superoxide dismutase (SOD) and catalase (CAT). However, a significant decline in CAT and SOD activity was observed at higher salinities (10 and 15 ppt).
NP exposure on fish at 0 ppt significantly decreased CAT and SOD levels compared to the control treatment (0 ppt, non- NPs), by T-test statistic analyzis with p < 0.05. It showed a consistent result with other study that exposure to high concentrations, NPs could inhibit the activities SOD and CAT in medaka fish (Oryzias latipes) [24]. This reduction is most likely due to direct oxidative stress triggered by nanoparticles (NPs), which leads to increased ROS production. The rise in ROS results from the entry of NPs into the plasma membrane via endocytosis mechanisms [25]. The excessive accumulation of ROS surpasses the capacity of the antioxidant defence system.
At lower salinity (0 - 5 ppt), exposure NPs enhanced significantly the activities of superoxide dismutase (SOD) and catalase (CAT) with p < 0.05, in which it shows a similar trend with the last study that increasing salinity may also activate antioxidant enzymes in fish [26]. This aligns with findings that mild environmental stressors can stimulate adaptive responses in fish, including upregulation of antioxidant enzymes.
While, exposured NPs at higher salinities (10 and 15 ppt) decreased significantly antioxidant activity of fish (with p < 0.05), reflected the combined oxidative burden from both high salinity and NP toxicity, consequently, the antioxidant activity declines due to cellular damage and exhaustion of the antioxidant defense system. Elevated salinity levels can cause ionic imbalance, dehydration, and increased metabolic stress in freshwater fish like tilapia, which are not fully adapted to marine-like conditions [27] When combined with NP exposure, this stress may surpass the fish’s compensatory antioxidant capacity, leading to enzymatic depletion or inhibition [28]. High salinity affects the permeability of cell membranes, which indicates allowing NPs to easily enter cells and cause ROS.These findings demonstrate that the effect of NPs on antioxidant enzymes is not only dose-dependent but also influenced by the salinity of the aquatic environment, highlighting the importance of environmental context when assessing NP toxicity in aquatic organisms.
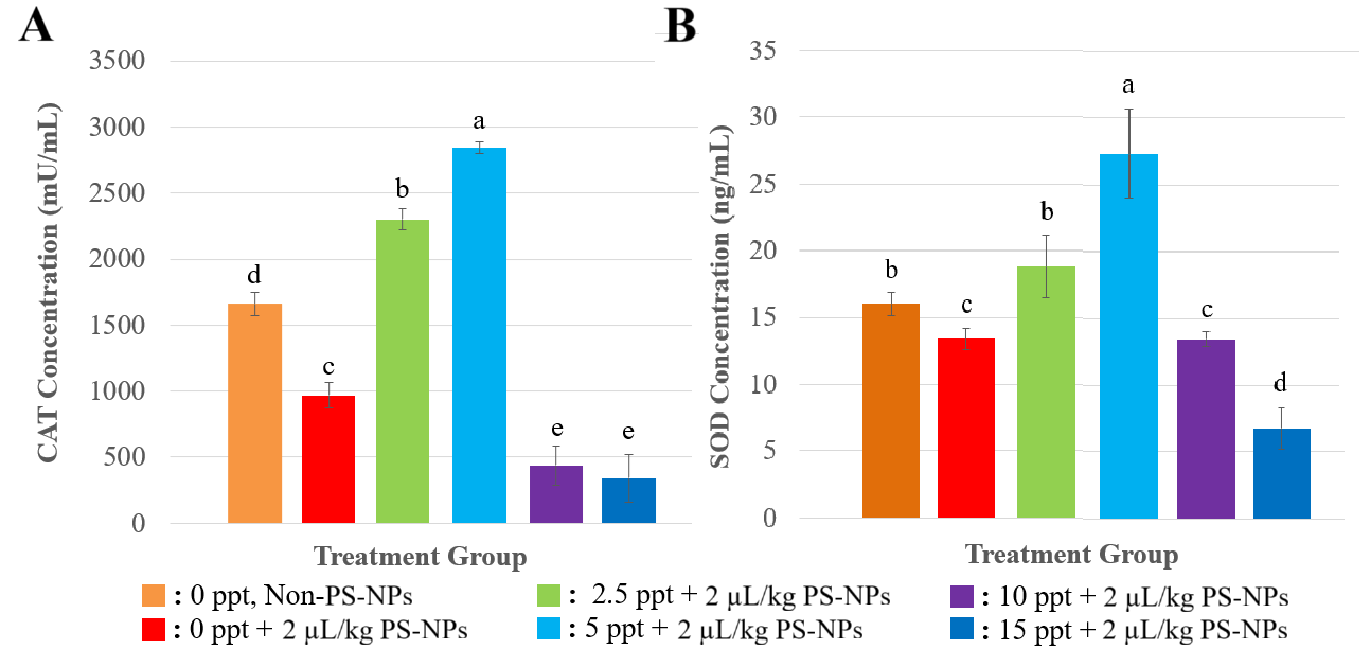
Figure 1 Concentration of Antioxidants after response to Several Treatments (A) CAT concentration, and (B) SOD concentration. All of the data were displayed by mean ±SD (α = 0.05). Different letters indicate statistically significant differences (p < 0.05).
Effects of nanoplastics on apoptosis related enzymes (Caspase-3, dan Caspase-9)
The levels of apoptosis-related enzymes Caspase-3 and Caspase-9 at Figure 2 showed dynamic changes in response to PS-NP exposure under varying salinity conditions. At 0 ppt (freshwater), NP exposure significantly increased (p < 0.05) in both Caspase-3 and Caspase-9 expression, compared with control treatment (0 ppt, non NPs). Caspase-3 and Caspase-9 levels were slightly reduced at lower salinity (2.5 and 5 ppt, meanwhile in higher salinities (10 and 15 ppt) showed a significant upregulation of Caspase-3 and Caspase-9 was observed.
NP exposure at 0 ppt (freshwater) significantly increased both Caspase-3 and Caspase-9 expression (p < 0.05), indicating elevated apoptotic activity. This response is likely driven by oxidative stress caused by accumulated ROS due to NP exposure[29]. The concurrent decrease in antioxidant enzymes (CAT and SOD) at this salinity supports the notion that the antioxidant defence was insufficient to neutralize ROS, thereby triggering mitochondria-mediated apoptosis pathways involving Caspase-9 activation followed by Caspase-3 [13]. The last study in the liver of Monopterus albus, exposure to PS-MPs elevated the expression of caspase 9, suggesting that NPs trigger apoptosis in liver tissue [30]. NPs increased expression gen Casp3a in Oreochromis niloticus organ [31].
At lower salinity (0 - 5 ppt), exposure NPs reduced significantly Caspase-3 and Caspase-9 levels of fish (p < 0.05). This pattern coincided with a rise in CAT and SOD activity, suggesting that moderate salinity may have helped restore redox balance, alleviating oxidative damage and suppressing apoptotic signalling. Previous studies have demonstrated that mild salinity stress can enhance antioxidant enzyme efficiency and promote cellular homeostasis in euryhaline species like Oreochromis niloticus. By improving osmoregulatory function and reducing oxidative load, moderate salinity may have attenuated mitochondrial distress, thereby reducing caspase activation.
While, in higher salinities (10 and 15 ppt), the graph show a significant upregulation of Caspase-3 and Caspase-9 (p < 0.05), reflecting intensified apoptotic responses. Paralleled the sharp decline in CAT and SOD activity, suggesting that oxidative stress once again exceeded the fish’s antioxidant capacity. The dual stress from salinity and NP exposure likely results in mitochondrial dysfunction and increased cytochrome c release, which activates the intrinsic apoptosis pathway via Caspase-9 and subsequently Caspase-3 [28]. The observed pattern indicates a strong interplay between oxidative stress and apoptotic pathways. When antioxidant defences are functional (as seen at 2.5 until 5 ppt), apoptosis is minimized. However, under conditions of antioxidant depletion (0, 10, and 15 ppt), caspase activation intensifies. These findings highlight the importance of maintaining redox balance in mitigating NP-induced cytotoxicity, and the modulatory role of environmental salinity in shaping physiological responses.
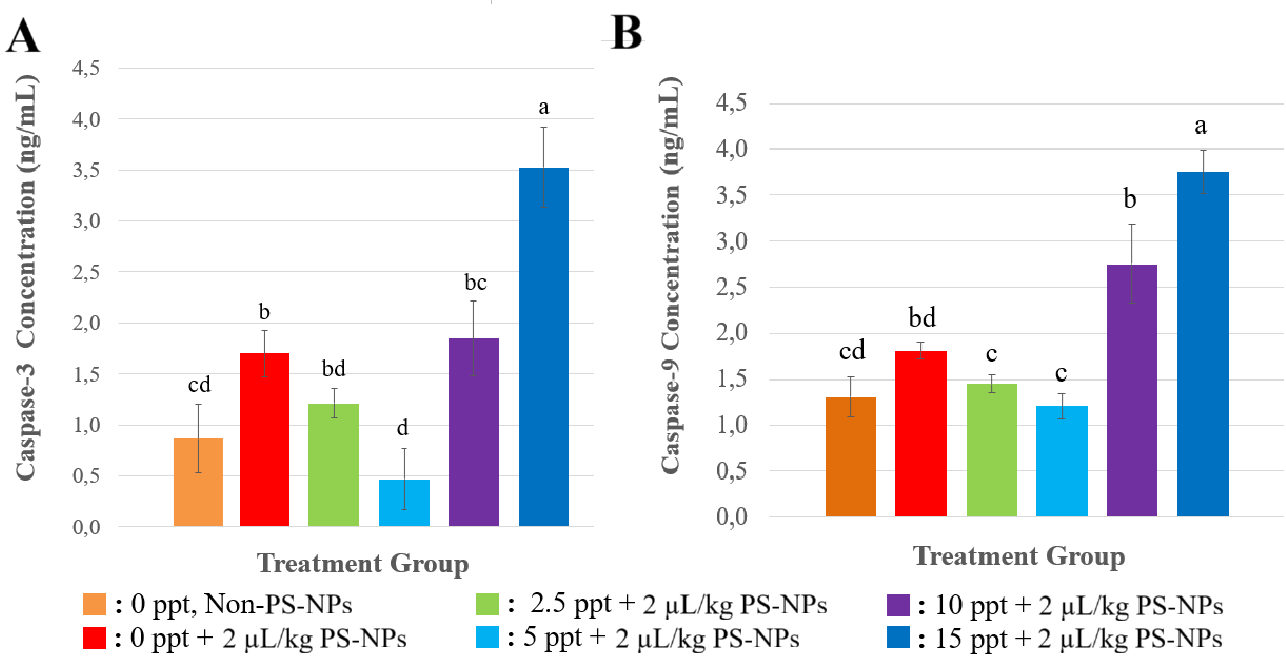
Figure 2 Concentration of Apoptosis Related Enzymes after response to Several Treatments (A) Caspase-3 concentration, and (B) Caspase-9 concentration. All of the data were displayed by mean ±SD (α = 0.05). Different letters indicate statistically significant differences (p < 0.05).
Effects of nanoplastics on liver tissue
On ORP analyzis, between treatment NPs and non-NPs in 0 ppt, the oxidation reduction potential (ORP) at Figure 3 in the liver tissue of Nile tilapia significantly increased on exposed-NPs’s fish (p < 0.0001). This elevation in ORP indicates a dominantly oxidative environment within the hepatic tissue, reflecting a reduction in intracellular antioxidant capacity as a result of NP exposure. In different salinity level, the ORP of the liver tissue of fish exposed NPs on moderate levels of 2.5 to 5 ppt showed a marked decline (p < 0.0001). However, in higher salinity (10 - 15 ppt), the ORP raised on fish exposured NPs (p < 0.0001).
The elevation in ORP at salinity of 0 ppt caused by NP exposure indicates a dominantly oxidative environment within the hepatic tissue, reflecting a reduction in intracellular antioxidant capacity as a result of NP exposure. Such an oxidative condition is likely to exacerbate oxidative stress through the accumulation of reactive oxygen species (ROS) that cannot be adequately detoxified. On lower salinity level (2.5 to 5 ppt), this suggests a shift towards a more reductive condition, which may be associated with physiological adaptation of the fish to moderate salinity, including the upregulation of endogenous antioxidant systems. The activities of the antioxidant enzymes superoxide dismutase (SOD) and catalase (CAT) were observed to increase under these conditions, indicating the tissue’s capacity to neutralise ROS and thereby reduce oxidative pressure, as reflected in the lower ORP values.
While, under high salinity conditions (10 - 15 ppt) Nile tilapia was unable to maintain osmotic and redox homeostasis, leading to renewed oxidative stress. This was corroborated by a tendency for SOD and CAT activities to decrease at higher salinities, suggesting antioxidant system exhaustion or impairment due to the combined stress of nanoplastics and hyperosmotic challenge. Overall, the fluctuations in ORP within the hepatic tissue of tilapia exposed to NPs under varying salinities were closely related to the dynamics of the antioxidant enzymes SOD and CAT. Optimal antioxidant activity was observed at moderate salinities (2.5 - 5 ppt), whereas at extreme salinity levels (0 and ≥ 10 ppt), the failure of the antioxidant response was reflected in the elevation of tissue ORP [32].
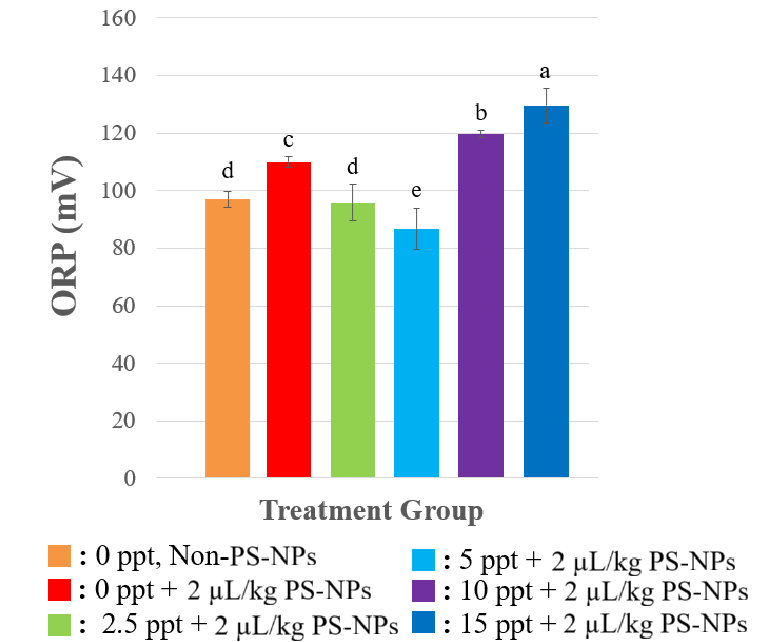
Figure 3 ORP Value in the liver tissue after response to Several Treatments. All of the data were displayed by mean ±SD (α = 0.05). Different letters indicate statistically significant differences (p < 0.05).
On hepatic cell analyzis, exposure to NP particles at 0 ppt salinity resulted in a significant reduction in the normal hepatic cell count (p < 0.0001) (Figure 4). Conversely, at moderate salinities of 2.5 - 5 ppt, normal cell density was restored closer to control levels, suggesting that mild salinity may promote ionic interactions that reduce NP absorption or enhance osmotic balance, thus protecting hepatocyte integrity. However, under higher salinities (10 - 15 ppt) the normal cell count declined again significantly.
The decrease in the normal hepatic cell counts at 0 ppt caused by NP exposure is likely attributable to heightened cellular stress and damage induced by NP accumulation in freshwater conditions, where ionic competition is minimal, facilitating higher NPs bioavailability and uptake into hepatic tissues. In contrast, at intermediate salinity levels (2.5 - 5 ppt), the cell density became comparable to that of the control group, implying that mild salinity might support ionic interactions that either reduce NP absorption or enhance osmotic homeostasis, thus maintaining hepatocyte structure. Nonetheless, when salinity increased to higher levels (10 - 15 ppt), there was a marked reduction in normal cell numbers once again. This pattern may reflect compounded osmotic stress and higher ionic imbalances, exacerbating nanoplastic toxicity through synergistic osmotic and oxidative damage [33].
About the presence of oedematous cells, necrotic cells, Kupffer cells, as well as the diameters of the portal vein, central vein, and hepatic sinusoids, all these showed a consistent trend of increasing significantly (p < 0.0001) upon NP exposure at 0 ppt, compared control (0 ppt, non-NPs). Exposure NPs in different salinity levels, about 2.5 - 5 ppt, these histological indicators diminished, in line with improved hepatocyte viability and less severe vascular or inflammatory lesions, suggesting a protective role of moderate salinity against NPs-induced damage. Nevertheless, in higher salinity levels (10 - 15 ppt), all these lesion parameters increased after exposure NPs, reflecting aggravated hepatic stress.
NP exposure at 0 ppt led to a consistent increase in the number of edematous and necrotic cells, Kupffer cell presence, and the diameters of the portal vein, central vein, and hepatic sinusoids. These pronounced elevations suggest significant inflammatory activity, vascular congestion, and damage to hepatocytes, likely resulting from heightened NP accumulation under freshwater conditions. However histological alterations at salinity levels between 2.5 and 5 ppt, were reduced, corresponding with improved hepatocyte health and fewer vascular or inflammatory abnormalities, indicating that moderate salinity may exert a protective effect against NP-induced hepatic damage, but because of the increasing salinity to 10 - 15 ppt, these pathological markers rose once more, suggesting intensified liver stress, related with the last study showed that in higher salinity, histoligical assesment showed more abnormality in liver [9]. This may be due to elevated salinity disrupting ionic balance, amplifying oxidative stress, and weakening both cellular and vascular structures, thereby worsening the effects of nanoplastic exposure.
Overall, the pattern suggests that lower salinity (2.5 - 5 ppt) provides a physiological buffer that mitigates NP toxicity by supporting osmotic and ionic stability, while in higher salinity (10 - 15 ppt) intensified histological damage through either increased nanoplastic uptake or a combination of osmotic disturbance and nanoplastic stress. This is evidenced by increases in sinusoidal diameter, portal and central vein dilation, as well as the proliferation of Kupffer cells, all of which reflect intensified inflammation and vascular congestion under extreme salinity or zero-salinity conditions [9]. These findings suggest that NP pollution, particularly under fluctuating salinity, could severely impact fish health and ecosystem stability.
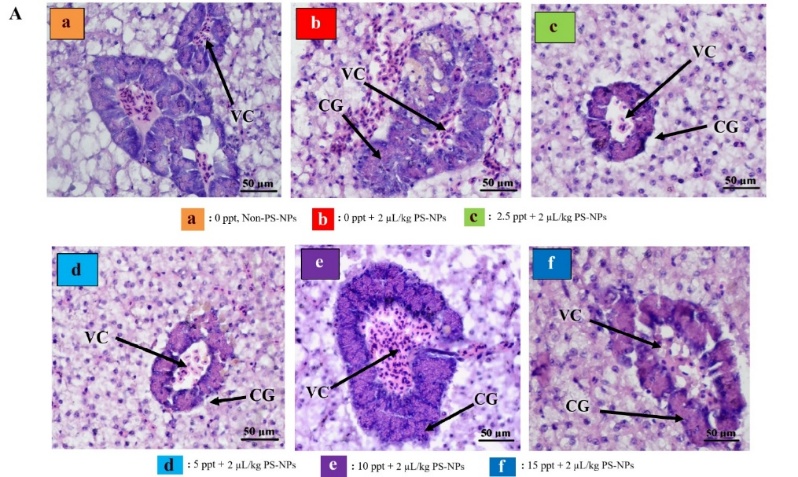
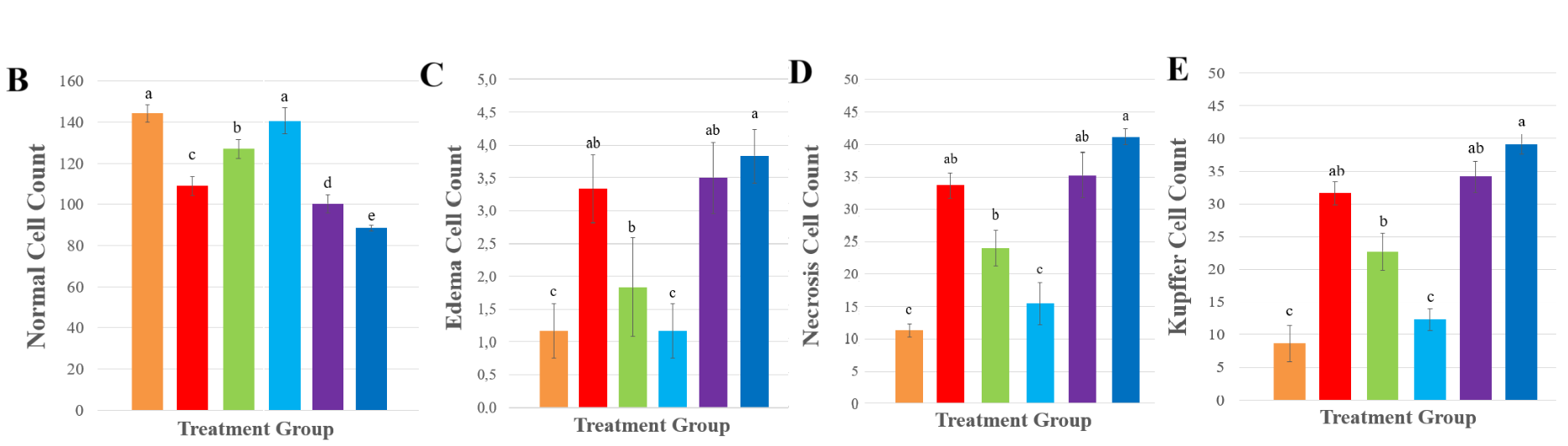
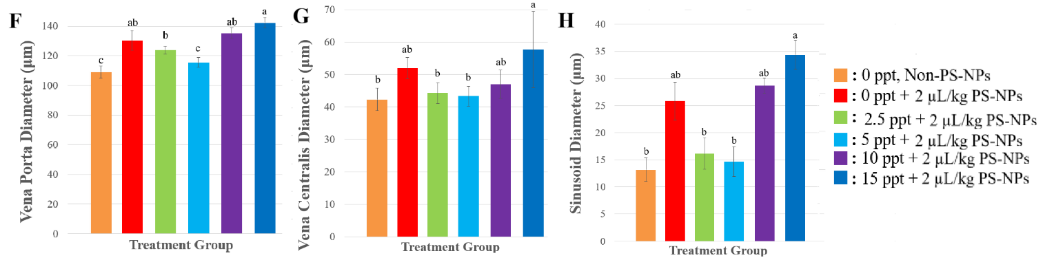
Figure 4 Effect of PS-NPs to Tilapia Histology (A) Histology of liver injury at 400× magnification. VC: Vena Centralis and CG: Congestion. Sections were stained with hematoxylin and eosin. (B) normal cells, (C) edema cells, (D) necrosis cells, (E) Kupffer cells, (F) vena porta diameter, (G) vena centralis diameter, and (H) sinusoid diameter. All of the data were displayed by mean ± SD (α = 0.05). Different letters indicate statistically significant differences (p < 0.0001).
Molecular docking assesment results on the effects of polystyrene nanoplastics on antioxidant endogenous and apoptosis Related-Enzyme
The molecular docking results in Table 1 and Figure 5 illustrate the predicted binding interactions between the styrene monomer and target proteins involved in antioxidant defense and apoptosis. Molecular docking analysis predicts binding affinity and chemical interactions between amino acids in the ligand and receptor [34]. Among the tested proteins, styrene showed the highest binding affinity with catalase (−5.3 kcal/mol) and caspase-9 (−4.6 kcal/mol), compared to superoxide dismutase and caspase-3. The interaction between styrene and CAT involved Van der Walls and hydrophobic contacts with several amino acid residues on chain A, while styrene and caspase-9 binding was also dominated by Van der Waals and hydrophobic interactions. These interactions patterns suggest that styrene monomers may associate with regions of catalase and caspase-9 that are relevant to protein stability or function [35,36].
In apoptosis-related proteins, caspase-9 showed stronger predicted interaction with styrene than caspase-3. Caspase-9 is known to play a central role in the intrinsic apoptosis pathway and in mitochodrial homeostasis regulation [37]. Therefore, interaction of styrene with caspase-9 may be relevant for understanding potential molecular-level disturbances associated with nanoplastic exposure, although validation is required to confirm these effects. Disruption of caspase-9 protein by styrene monomers can disrupt the caspase activation pathway and potentially disrupt body homeostasis [38].
It is important to note that docking results provide predictive information on binding feasibility rather than direct evidence of functional inhibition. The observed binding affinities indicate potential molecular interactions that may contribute to altered protein behavior under nanoplastic exposure, but they do not confirm enzymatic inactivation or direct biological effects in vivo. Similar docking-based observations of nanoplastic-protein interactions have been reported for other enzymes, supporting the use of in silico approaches as an initial mechanistic screening tool [35].
The study showed that SOD and CAT activity decreased significantly at freshwater salinity levels because of high reactive oxygen species (ROS) buildup. Biological membranes receive penetration from NPs which leads to oxidative stress because ROS accumulates beyond liver tissue antioxidant capacity. Elevated caspase-3 and caspase-9 activity indicates the start of mitochondria-mediated apoptosis which matches cytochrome c release and apoptosome formation. This pathway ultimately leads to cellular degradation and death.
Interestingly, at moderate salinities (2.5 - 5 ppt), antioxidant enzyme activity increased, whereas apoptosis markers were reduced. This indicates an adaptive physiological response in which mild salinity stabilizes protein folding, reduces NP uptake, or enhances ionic/osmotic balance. These conditions may enable hepatic resilience through optimizing enzymatic function and minimizing oxidative burden. However, at increased salinities (10 - 15 ppt), the reverse trend was observed again. SOD and CAT activity reduced once more, and caspase activation heightened. This is likely due to the combined effect of salinity stress and nanoplastic toxicity, both leading to damage in membrane permeability, protein synthesis, and mitochondrial integrity [28]. These findings validate the point that extreme salinity, either hyp- or hyperosmotic, can enhance nanoplastic toxicity.
To understand the molecular mode of enzyme destabilization, in silico molecular docking studies were carried out with styrene monomer (CID:7501). The findings revealed the maximum binding affinity for styrene with CAT (−5.3 kcal/mol) and for styrene with caspase-9 (−4.6 kcal/mol), indicating the disruption of protein structure and function by nanoplastics. Interaction with certain active site critical residues via Van der Waals and hydrophobic interactions can initiate conformational adjustments that result in reduced enzyme activity or launch pro-apoptotic pathways [35]. Styrene molecules form dominant hydrophobic interactions, including π interactions with aromatic residues in the binding pocket. Although docking affinity values provide an initial indication of binding possibility, docking scores are semi-quantitative and are influenced by scoring function parameters and protein flexibility treatment.
Another Study reported that long-term exposure to PS-NPs in animal models of Epinephelus coioides and Larimichthys polyactis fish showed inhibition of digestive enzymes and endogenous antioxidant modulators [39]. Digestive enzymes play an important role in living organisms. Salivary glands that secrete α-amylase in the mouth, which is useful for helping break down polysaccharides into monosaccharides, appear to be affected by nanoplastics [40]. At the cellular and molecular levels, polystyrene nanoplastics will affect performance in the body of organisms and can trigger long-term plastic accumulation in the body.
Table 1 Molecular docking experiment results.
Compound |
Protein receptors |
Type of protein receptors |
Complex binding affinity (kcal/mol) |
Type of interaction |
Amino acids involved in interaction |
Styrene |
Superoxide dismutase (SOD) |
Antioxidant endogenous |
−4.7 |
vdw |
Ser72(D), Pro73(D) |
HI |
Val8(D), Leu4(D), Leu71(D) |
||||
Catalase (CAT) |
−5.3 |
vdw |
Thr146(A), Ser114(A), Gly131(A), Arg112(A), Arg72(A), Ala333(A), Tyr358(A) |
||
HI |
Phe334(A), His362(A) |
||||
Caspase-3 |
Apoptosis-related enzyme |
−4.0 |
vdw |
Glu261(A), Asn221(A), Leu230(A), Tyr224(A) |
|
HI |
Trp227(A) |
||||
Caspase-9 |
−4.6 |
vdw |
Gln253(A), Thr318(A), Pro317(A), Ser320(A), Asp321(A), Gly254(A), Lys257(A) |
||
HI |
Leu316(A) |
PHI: Polar hydrogen interaction; HI: Hydrophobic interaction; vdw: Van der Walls interaction.
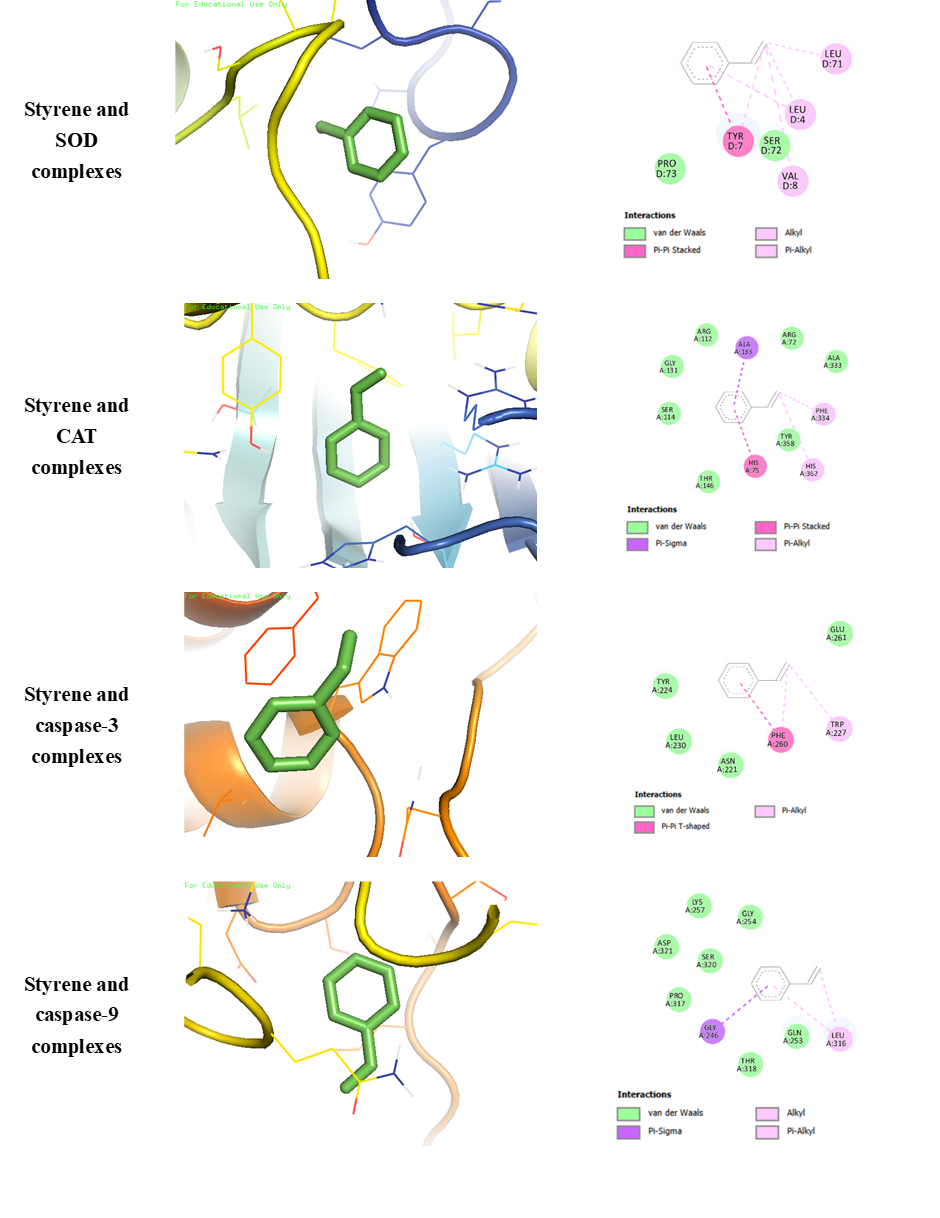 Figure
5
Visualization of each complexes binding (left: Three-dimensional
molecular interaction binding; right: Two-dimensional molecular
interaction binding with amino acids involved).
Figure
5
Visualization of each complexes binding (left: Three-dimensional
molecular interaction binding; right: Two-dimensional molecular
interaction binding with amino acids involved).
Validation of molecular docking experiment results
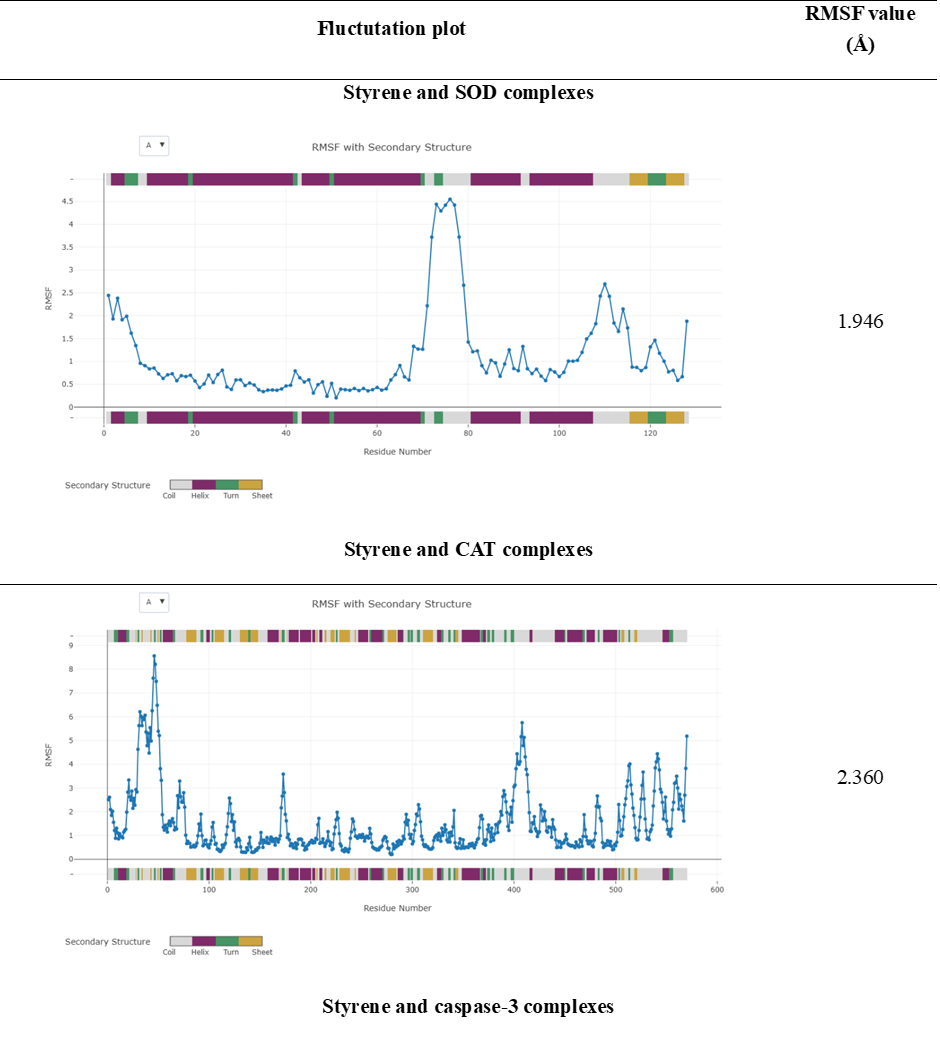
Validation of the molecular docking results was performed to assess the structural stability of ligand-protein complexes through root mean square fluctuation (RMSF) analysis. RMSF values are commonly used to evaluate residue-level flexibility, where values below 3 Å indicate stable interactions between ligands and protein receptors [41]. As shown in Table 2, all styrene-protein complexes exhibited RMSF values below 3 Å, indicating overall structural stability during the simulations. The styrene-SOD complex showed the lowes RMSF value (1.946 Å), followed by styrene-caspase-3 (2.144 Å), styrene-caspase-9 (2.263 Å), and styrene-CAT (2.360 Å). These results suggest that the predicted docking poses are structurally acceptable and do not induce excessive conformational fluctuations in the protein backbones. Therefore, nanoplastics most likely affect enzyme activity not just by inducing oxidative stress, but also by the direct alteration of enzyme structure by molecular interaction, a condition exacerbated under stressful environmental pressures such as salinity high or very low. It should be noted that the use of styrene monomers is a simplification of actual polystyrene particles; nanoplastics in biological environments generally form protein/eco-coronas that significantly alter surface properties and biological interaction patterns.
The complex with the lowest RMSF (styrene–SOD) tends to be more stable than other complexes, so this interaction can interfere with enzyme activity more consistently. Conversely, a slightly higher RMSF value in the styrene–CAT complex (2.360 Å) indicates conformational flexibility in certain residues, although it is still in the stable category. Validation using RMSF analysis is crucial because static docking often only provides binding predictions for a single protein conformation. With dynamic simulations, it is possible to determine how bond stability holds up under conditions that more closely resemble the biological environment. It should be emphasized that RMSF analysis reflects the dynamic stability of docked complexes rather than direct functional inhibition of proteins. While stable ligand-protein interactions may support the feasibility of molecular association [42].
Table 2 Molecular dynamics results.
Conclusions
This study demonstrates that NPs significantly influence the oxidative stress response and apoptosis regulation in Oreochromis niloticus under varying salinity conditions. Between NP and non-NP treatment in the same salinity, NP exposure reduced antioxidant enzyme activity (CAT and SOD) and elevated apoptosis-related enzymes (Caspase-3 and Caspase-9), indicating that NPs intrinsically induce oxidative stress and apoptotic responses in fish. Exposure NPs in higher salinities (10 - 15 ppt) indicated physiological stress, indicated that elevated salinity acts synergistically with NP exposure to intensify oxidative stress and apoptosis. Interestingly, lower salinity levels (2.5 - 5 ppt) appeared to buffer NP-induced toxicity, as reflected by improved antioxidant activity and reduced apoptotic signalling.
Molecular docking experiments also showed evidence that NPs can affect the stability of proteins from the endogenous antioxidant and apoptosis-related enzyme groups. The CAT (−5.3 kcal/mol) and caspase-9 (−4.6 kcal/mol) proteins were found to be most affected by the presence of NPs in these cells, as evidenced by their highest binding affinity compared to other test proteins.
These findings highlight the complex interaction between environmental salinity and NP toxicity, emphasizing the need for context-dependent risk assessments of NPs in aquatic ecosystems. Therefore, It will be important to explore potential remediation strategies by other researcher, such as the use of biofiltration or phytoremediation, to address the growing issue of NP contamination in aquatic ecosystems. Future research should assess the broader ecological impacts of NP pollution on aquatic biodiversity and food webs, as these findings may have cascading effects on ecosystem stability. Moreover, the long-term effects of NP exposure, including potential transgenerational impacts, should be investigated to understand the full ecological consequences of plastic pollution.
Acknowledgements
The
authors express their sincere gratitude to the Directorate of
Research, Technology, and Community Service, Ministry of Education,
Culture, Research, and Technology, Universitas Airlangga, Indonesia,
for providing research funding through the Master’s Thesis
Research Grant for the 2025 fiscal year, under contract number
2419/B/UN3.LPPM/PT.01.03/2025.
Novelty statement
Variations in salinity levels have been shown to significantly influence NP toxicity, with higher salinity intensifying the toxic effects of NPs, particularly regarding antioxidant activity, apoptosis induction, and liver histological damage in fish.
Ethics approval and consent to participate
This research conformed to the guidance of animal ethical treatment for the care and use of experimental animals. Ethical approval for this research was granted by the Health Research Ethics Clearance Commission, Faculty of Dental Medicine, Universitas Airlangga (0885/HRECC.FODM/VIII/2024).
Declaration of Generative AI in Scientific Writing
Manuscript did not involve the use of artificial intelligence–based tools. All content was authored and validated by the authors.
CRediT Author Statement
Muhammad Nuh Fathsyah Siregar: Writing – Original Draft, Conceptualization; Methodology, Formal Analysis, Investigation, Data Curation, Visualization, and Writing - Review & Editing. Alfiah Hayati: Writing – Original Draft, Conceptualization; Methodology, Validation, Resources, Funding acquisition, Supervision, Writing - Review & Editing, and Project administration. Muhammad Iqbal: Writing – Original Draft, Conceptualization; Methodology, Formal Analysis, Investigation, Project administration, Data Curation, Visualization, and Writing - Review & Editing. Mochammad Aqilah Herdiansyah: Writing – Original Draft, Software and Formal Analysis. Firli Rahmah Primula Dewi: Writing - Review & Editing and Formal Analysis. Manikya Pramudya: Writing - Review & Editing, Investigation, Data Curasion, Project administration, and Formal Analysis. Aunurohim: Writing - Review & Editing. Hisyam Rizky Saputra: Writing - Review & Editing and Investigation. Febriyansyah Saputra: Writing - Review & Editing.
References
[1] A Hayati, M Pramudya, H Soepriandono, AR Astri, MR Kusuma, S Maulidah, W Adriansyah and FRP Dewi. Assessing the recovery of steroid levels and gonadal histopathology of tilapia exposed to polystyrene particle pollution by supplementary feed. Veterinary World 2022; 15(2), 517-523.
[2] Aunurohim, D Saptarini, A Junaedi, S Dewi, E Danilyan, H Adro’i, P Putra and A Hayati. Characteristics of microplastics in water and fish and their relationship with migration from the East Coast of Surabaya, Indonesia. Egyptian Journal of Aquatic Biology and Fisheries 2025; 29(2), 2371-2392.
[3] A Hayati, WP Wilujeng, M Pramudya, FRP Dewi, Sugiharto, B Muchtaromah and RJK Susilo. The effect of exposure to polystyrene nanoplastics on cytokine levels and reproductive system of male tilapia. Tropical Journal of Natural Product Research 2024; 8(2), 6300-6303.
[4] H Triwahyudi, L Soehargo, L Muniroh, RN Qolbi, TQ Aini, RFZ Kurnia, PAD Putra, M Pramudya, B Muchtaromah and A Hayati. Potential of red seaweed ( Dichotomania obtusata ) on immune response and histopathology of rat testis exposed to nanoplastics. Tropical Journal of Natural Product Research 2023; 7(5), 2969-2973.
[5] J Kong, L Zhao, Q Chen, T Gao, P Xu and F Sun. Review of the toxic effects and mechanisms of polystyrene micro/nanoplastics across multiple animal species. Journal of Hazardous Materials Advances 2025; 20, 100938.
[6] A Hayati, M Pramudya, Aunurohim, AS Junaedi, FA Nurbani, WP Wilujeng, M Iqbal, FRP Dewi and V Lim. Micro-Nanoplastics pollution and its oxidative stress-induced effects on fish physiology in coastal waters of surabaya. Journal of Animal Health and Production 2025; 13(2), 435-444.
[7] M Yang and WX Wang. Differential cascading cellular and subcellular toxicity induced by two sizes of nanoplastics. Science of The Total Environment 2022; 829, 154593.
[8] J Pei, S Chen, L Li, K Wang, A Pang, M Niu, X Peng, N Li, H Wu and P Nie. Impact of polystyrene nanoplastics on apoptosis and inflammation in zebrafish larvae: Insights from reactive oxygen species perspective. Science of The Total Environment 2024; 948, 174737.
[9] M Iqbal, MNF Siregar, GZ Sofa, HR Saputra, M Pramudya, FRP Dewi, A Soegianto, F Saputra, Aunurohim and A Hayati. Nanoplastic-Induced immune modulation and histopathological changes in oreochromis niloticus at different SalinityNanoplastic-Induced immune modulation and histopathological changes in oreochromis niloticus at different salinity levels levels. Journal of Animal Health and Production 2025; 13(2), 235-242.
[10] RJK Susilo, M Pramudya, FRP Dewi, W Seftiarini, D Hidayati, A Aunurohim, V Lim, MA Herdiansyah and A Hayati. Adverse effect of polystyrene nanoplastics in impairing glucose metabolism in liver injury. International Journal of Molecular Sciences 2025; 26(10), 4870.
[11] J Li, H Liu and JP Chen. Microplastics in freshwater systems: A review on occurrence, environmental effects, and methods for microplastics detection. Water Research 2018; 137, 362-374.
[12] H Wu, J Guo, Y Yao and S Xu. Fish and shellfish immunology polystyrene nanoplastics induced cardiomyocyte apoptosis and myocardial inflammation in carp by promoting ROS production. Fish & Shellfish Immunology 2022; 125, 1-8.
[13] M Safitri, FA Nurbani, ML Hidayaturrohman, F Jamaluddin, KR Wulandari, M Pramudya, FRP Dewi, L Vuanghao and A Hayati. Potential of macang (Mangifera foetida) bark extract on antioxidant levels and pro-a poptotic proteins in rats (Rattus norvegicus) exposed to polystyrene nanoplastics. Sains Malaysiana 2025; 54(6), 1477-1487.
[14] FJ Emon, J Hasan, SIM Shahriar, N Islam, MS Islam and M Shahjahan. Increased ingestion and toxicity of polyamide microplastics in Nile tilapia with increase of salinity. Ecotoxicology and Environmental Safety 2024; 282, 116730.
[15] HM Pratiwi, T Takagi, S Rusni and K Inoue. Euryhaline fish larvae ingest more microplastic particles in seawater than in freshwater. Scientific Reports 2023; 13(1), 1-10.
[16] E Besseling, JTK Quik, M Sun and AA Koelmans. Fate of nano- and microplastic in freshwater systems: A modeling study. Environmental Pollution 2017; 220, 540-548.
[17] AJR Watts, MA Urbina, S Corr, C Lewis and TS Galloway. Ingestion of plastic microfibers by the crab carcinus maenas and its effect on food consumption and energy balance. Environmental Science & Technology 2015; 49(24), 14597-14604.
[18] B Li, W Liang, L Quan-Xing, S Fu, C Ma, Q Chen, L Su, NJ Craig and H Shi. Fish ingest microplastics unintentionally. Environmental Science & Technology 2021; 55(15), 10471.
[19] CT Walsh and BC Pease. The use of clove oil as an anaesthetic for the longfinned eel, Anguilla reinhardtii (Steindachner). Aquaculture Research 2002; 33(8), 627-635.
[20] R Siskandar, A Hendriana, J Ekasari and BR Kusumah. Automated Redox Monitoring System (ARMS): An instrument for measuring dissolved oxygen levels using a potential redox sensor (ORP) in a prototype of shrimp farming pond with an Internet-Based monitoring system. Journal of Aquaculture And Fish Health 2022; 11(2), 238-246.
[21] MRT Alifiansyah, MA Herdiansyah, RC Pratiwi, RP Pramesti, NW Hafsyah, AP Rania, ERPP Ju, PA Cahyono, Litazkiyyah, SK Muhammad, AAA Murtadlo, VD Kharisma, ANM Ansori, V Jakhmola, PK Ashok, JM Kalra, H Purnobasuki and IA Pratiwi. QSAR of acyl alizarin red biocompound derivatives of Rubia tinctorum roots and its ADMET properties as anti-breast cancer candidates againts MMP-9 protein receptor: In silico study. Food Systems 2024; 7(2), 312-320.
[22] AN Muhammad, A Ijaz, A Khalid, K Rafique, R Tabassum and A Zahra. Renal effects of graphite oxide sheets in albino rats (Rattus norvegicus): A preliminary study. BIO Integration 2024; 5(1), e966.
[23] MA Herdiansyah, R Rizaldy and MRT Alifiansyah. Molecular interaction analysis of ferulic acid (4-hydroxy-3-methoxycinnamic acid) as main bioactive compound from palm oil waste against MCF-7 receptors: An in silico study. Narra J 2024; 4(2), e775.
[24] Y Zhou, L Zhao, H Xu, EG Xu, M Li and Y Wang. Long-Term exposure to polystyrene nanoplastics impairs the liver health of medaka. Water 2022; 14(17), 2767.
[25] F Saputra, AD Pramata, A Soegianto and SY Hu. Polystyrene nanoplastics cause developmental abnormalities, oxidative damage and immune toxicity in early zebrafish development. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology 2025; 295, 110216.
[26] MN Monier, ASA El-Naby, RM Fawzy, F Samir, SHH Shady, YS Grana, NM Albaqami and M Abdel-Tawwab. Growth performance, antioxidant, and immune responses of Nile tilapia (Oreochromis niloticus) fed on low-fishmeal diets enriched with sodium chloride and its adaptability to different salinity levels. Fish Physiology and Biochemistry 2025; 51(1), 6.
[27] P Angadi, M Das and R Roy. Effect of high salinity acclimation on glucose homeostasis in Mozambique tilapia (Oreochromis mossambicus ). Fish Physiology and Biochemistry 2021; 47(6), 2055-2065.
[28] X Zhang, X Chen, L Gao, Z Hai-Ting, J Li, Y Ye, Z Qing-Ling, Z Jia-Lang and X Yan. Transgenerational effects of microplastics on Nrf2 signaling, GH/IGF, and HPI axis in marine medaka Oryzias melastigma under different salinities. Science of the Total Environment 2024; 906, 167170.
[29] Y Lu, Y Zhang, Y Deng, W Jiang, Y Zhao, J Geng, L Ding and H Ren. Uptake and accumulation of polystyrene microplastics in zebrafish (Danio rerio) and toxic effects in liver. Environmental Science & Technology 2016; 50(7), 4054-4060.
[30] Z Zhou, W Zhou, G Liu, C Zhu, M Han, T Zhu, Q Jiang and W Lv. Effects of polystyrene nanoplastics on apoptosis, digestive enzymes, and intestinal histological structure and flora of swamp eel (Monopterus albus). Environmental Science: Nano 2024; 11(3), 1085-1096.
[31] VP Vineetha, K Suresh and D Pillai. Impact of sub-chronic polystyrene nanoplastics exposure on hematology, histology, and endoplasmic reticulum stress-related protein expression in Nile tilapia (Oreochromis niloticus). Comparative Biochemistry and Physiology Part B: Biochemistry & Molecular Biology 2024; 273, 110982.
[32] A Agarwal and AD Bui. Oxidation-reduction potential as a new marker for oxidative stress: Correlation to male infertility. Investigative and Clinical Urology 2017; 58(6), 385-399.
[33] B Withyachumnarnkul, P Pongtippatee and J Ruangsri. Comparative proteomic profiling represents an inhibition of protein synthesis to regulate osmotic stress in Nile tilapia (Oreochromis niloticus) embryos. Comparative Biochemistry and Physiology Part D: Genomics and Proteomics 2024; 49, 101173.
[34] NS Aini, ANM Ansori, MA Herdiansyah, VD Kharisma, MH Widyananda, AAA Murtadlo, DDR Turista, TH Sucipto, S Sahadewa, FD Durry, V Jakhmola, M Rebezov, VP Kartashev, K Rachmawati, PA Wiradana, T Karpagam and H Purnobasuki. Antimalarial potential of phytochemical compounds from garcinia atroviridis Griff ex. T. Anders targeting multiple proteins of plasmodium falciparum 3D7: An in silico approach. BIO Integration 2024; 5(1), e967.
[35] A Azhagesan, N Chandrasekaran and A Mukherjee. Ecotoxicology and environmental safety multispectroscopy analysis of polystyrene nanoplastic interaction with diastase α-amylase. Ecotoxicology and Environmental Safety 2022; 247, 114226.
[36] MBD Ávila and WFDA Jr. Development of machine learning models to predict inhibition of 3-dehydroquinate dehydratase. Chemical Biology & Drug Design 2018; 92(2), 1468-1474.
[37] A Hyun-Kyu, KM Chung, H Park, J Hong, G Ji-Eun, H Choi, YW Lee, J Choi, JY Mun and Y Seong-Woon. CASP9 (caspase 9) is essential for autophagosome maturation through regulation of mitochondrial homeostasis. Autophagy 2020; 16(9), 1598-1617.
[38] SI Wanandi, A Limanto, E Yunita, RA Syahrani, M Louisa, AE Wibowo and S Arumsari. In silico and in vitro studies on the anti-cancer activity of andrographolide targeting survivin in human breast cancer stem cells. PLoS One 2020; 15(11), e0240020.
[39] Q Wang, F Huang, K Liang, W Niu, X Duan, X Jia, X Wu, P Xu and L Zhou. Polystyrene nanoplastics affect digestive function and growth in juvenile groupers. Science of the Total Environment 2022; 808, 152098.
[40] CP des Gachons and PAS Breslin. Salivary amylase: Digestion and metabolic syndrome. Current Diabetes Reports 2016; 16(10), 102.
[41] RM Wijaya, MA Hafidzhah, VD Kharisma and AA Parikesit. COVID-19 in silico drug with zingiber officinale natural product compound library targeting the mpro protein. Makara Journal of Science 2021; 25(3), 5.
[42] S Wang, X Sun, W Cui and S Yuan. MM/PB(GB)SA benchmarks on soluble proteins and membrane proteins. Frontiers in Pharmacology 2022; 13, 1018351.