Trends
Sci.
2026;
23(8):
12865
Biodegradation Potential of Glyphosate by Rhizobacteria Isolated from
Tithonia diversifolia: Characterization, Glyphosate Degrading,
and Molecular Identification
Agustian1,*, Chairin Nisa1, Parwanto1, Hermansah2 and Syafrimen Yasin2
1Soil Biology Laboratory, Department of Soil Science and Land Resources, Faculty of Agriculture,
Universitas Andalas, West Sumatra 25163, Indonesia
2Chemistry and Soil Fertility Laboratory, Department of Soil Science and Land Resources, Faculty of Agriculture, Universitas Andalas, West Sumatra 25163, Indonesia
(*Corresponding author’s e-mail: [email protected])
Received: 2 December 2025, Revised: 23 December 2025, Accepted: 30 December 2025, Published: 10 March 2026
Abstract
Glyphosate is a widely used broad-spectrum systemic herbicide that effectively controls weeds by inhibiting the synthesis of specific amino acids for the formation of plant proteins. Improper and repeated use can lead to the accumulation of residue in the soil, which may remain strongly absorbed over an extended period. Investigations into the biodegradation of glyphosate in soil under various environmental conditions are crucial for bioremediation efforts. Recently, a further step has been taken considering the use of rhizobacteria for the removal of glyphosate herbicide. Our research aimed to isolate naturally occurring rhizobacteria from Tithonia diversifolia (as a source of green manure) and assess the critical ecological factors influencing their growth and glyphosate degradation. Screening began by cultivating a one gram of T. diversifolia rhizosphere soil sample in Nutrient Broth (NB) media containing 15 mg mL−1 glyphosate for a week. The six isolates initially obtained were further subcultured in Nutrient agar (NA) containing nutrient agar and 15 mg mL−1 glyphosate. The morphological and physiological properties, phosphate solubilization, and IAA production were used in isolate characterization, in addition to glyphosate biodegradation. Only two isolates, TBr1 and TBr12, have shown the capacity to survive and biodegrade glyphosate. Both isolates exhibit optimal pH levels above 6.0 and optimal activity at 30 °C, demonstrating the fastest growth rates and abilities to break down glyphosate by 67% and 76%, respectively, within 7 days. Tested in a medium minus carbon, nitrogen, and phosphorus sources, both isolates showed the ability to hydrolyze glyphosate through CN and CP bonds. However, they had different CP lyase efficacy in metabolizing glyphosate. Based on 16S rDNA gene sequence analysis, TBr1 was identified as Burkholderia cepacia strain TBr1, and TBr12 as the Bacillus velezensis strain TBr12. Therefore, the ability of both isolates to degrade glyphosate, produce IAA, and dissolve phosphates makes them promising candidates for removing these emerging contaminants from the environment.
Keywords: Bioremediation, Glyphosate, Isolation, Rhizobacteria, Tithonia diversifolia
Introduction
Glyphosate, with the active ingredient N-[phosphonomethyl]glycine, is the most widely used herbicide in the world [1]. This water-soluble, non-selective compound demonstrates broad-spectrum efficacy. Glyphosate is commonly used in no-till agricultural practices before planting, during plant
growth, and after harvest [2] to control various types of weeds, including herbaceous and non-herbaceous plants, as well as specific shrubs, annual and perennial plants, brush, and broadleaf trees [3].
The herbicide glyphosate acts by inactivating the enzyme EPSPS (EC 2.5.1.19, 5-enolpyruvylshikimate-3-phosphate synthase), which is located in chloroplasts that play an essential role in the plant’s shikimic acid pathway [4]. Disruption of the shikimic pathway will inhibit the synthesis of proteins and secondary metabolites, such as phytoalexins, folic acid, cinnamic acid, lignin precursors, flavonoids, plastoquinone, and other phenolic substances, which contain the basic ingredients of aromatic amino acids [5].
Glyphosate is mainly hydrolyzed by microbial metabolism in the soil. However, significant soil adsorption can halt this process and extend the half-life of glyphosate from a few weeks to several months or even one year [6]. As a result, it negatively impacts the population and performance of soil microorganisms participating in the soil nutrient cycle [7], particularly in relation to soil dynamics, carbon, and phosphorus, as well as the activity of enzymes involved in organic phosphorus composition [8]. Lozano and Pizarro et al. [9] stated that the overuse and misuse of glyphosate harm the environment. The constant accumulation of glyphosate residues in various ecosystems has led to a chain of harmful effects, including plant damage [9,10] reduced soil biodiversity [11], and considerable human health risks [12]. Disruption of plant metabolism has also caused, along with the disruption of metabolic functions in human and aquatic organisms, as well as endocrine disturbances in the human population [2,7]. Thus, biodegradable-based strategies must be adopted to mitigate the impact of glyphosate on soils, as this chemical directly affects the microbial community and soil fauna that are crucial for both agriculture and human health [13].
Microbiology research results suggest that certain microorganisms can degrade glyphosate and utilize it as their source of carbon, phosphorus, and nitrogen [14]. Biodegradation of glyphosate microbially occurs in two primary biological ways: The first, CN bonds cleavage by an enzyme called glyphosate oxyreductase (Gox), which produces the intermediates glyoxylic acid and aminomethylphosphonic acid (AMPA) [15] and the second CP bonds cleavage, yielding sarcosine, glycine, and formaldehyde, through the action of enzyme CP lyase [16,17].
The investigation of soil pollutant bioremediation has declared that the application of rhizobacteria in the bioremediation process is of utmost importance [18], as is the research of [19,20] which showed that plant rhizospheres play an essential role in supporting and creating the microbiome and increasing the ability of rhizobacteria to metabolize various pollutants. Classifying these differences in microbiomes, Jia et al. [21] noted that there are differences between the plant microbiomes from one to another, such as those between cultivated and uncultivated soils and the complexity of plant-supporting interactions between rhizobacteria and plants in the rhizosphere, as shown by Macek et al. [22] and Montreemuk et al. [23]. Recently, further advances have focused on the use of plant-interacting bacteria, either individually or in combination with plants, for removing the glyphosate herbicide. Jia et al. [21] and Mohy-Ud-Din et al. [24] believe that the best way to mitigate the impact of glyphosate is to bio-inoculate plants with the bacteria. Hakim et al. [25], were the first to report the potential occurrence of this interaction in Tithonia diversifolia (Mexican sunflower). According to Jama et al. [26], this plant plays a unique and strategic role in agroecosystems, functioning as a nutrient-rich green manure source, an effective hedge or boundary plant, and a biological platform [27] for enhancing rhizobacteria-mediated bioremediation of glyphosate residues in soil. Nevertheless, the rhizobacterial potential associated with this plant remains insufficiently explored.
Therefore, the isolation and characterization of effective glyphosate-degrading bacteria in the rhizodegradation process of glyphosate residues is a strategic step to eliminate the negative impact of glyphosate in an environmentally friendly manner. This study aims to isolate and characterize the ability of the isolates obtained from the rhizosphere of T. diversifolia in degrading glyphosate and evaluate the environmental conditions required for the growth and biodegradation of glyphosate in soil.
Materials and methods
Chemicals and reagents used
Monsanto’s brand, Roundup herbicide (based on isopropylamine salts of glyphosate), with a content of 360 g of active ingredient glyphosate per liter, is obtained from stores that provide agricultural production needs. All other chemicals used are analytical grade products with high purity levels. In addition, the standard stock solution of glyphosate 1,000 mg L⁻¹ (Supelco 44690-U) used was obtained from Sigma-Aldrich.
The rhizospheric soil samples in this study were collected from the rhizospheres of T. diversifolia, which were grown at the experimental field station of the Agriculture Faculty of Universitas Andalas (00º54'36"S and 100º28'18"E in Limau Manis, Padang, West Sumatra, Indonesia) (Figure 1). The soil was classified as Ultisols with a clay texture (pH 5.48 and base saturation of 19.1%). The roots of the plant, along with the soil attached to the root hairs, were randomly collected from the field and then taken to the laboratory. All the soil grains from each root hair are mixed until homogeneous, put in sterile plastic bags, and then stored at 4 °C until use.

Figure 1 The sampling location (A) of rhizospheric soil from T. diversifolia, which is used as a hedgerow in the experimental field station (B) of the Faculty of Agriculture, Universitas Andalas.
The collection of soil samples
The rhizospheric soil samples in this study were collected from the rhizospheres of T. diversifolia, which were grown at the experimental field station of the Agriculture Faculty of Universitas Andalas (00º54'36"S and 100º28'18"E in Limau Manis, Padang, West Sumatra, Indonesia) (Figure 1). The soil was classified as Ultisols with a clay texture (pH 5.48 and base saturation of 19.1%). Composite soil samples from the rhizosphere of T. diversifolia were taken at random on three clumps of each of three rows of T. diversifolia, a hedge row plant. All the soil grains from each root hair are mixed until homogeneous, put in sterile plastic bags, and then stored at 4 °C until use.
Screening and isolation of glyphosate-degrading rhizobacteria
In this study, a glyphosate concentration of 15 mg mL⁻¹ was employed as a screening level to evaluate the tolerance and degradation potential of rhizobacterial isolates under high selective pressure. The glyphosate concentration applied was set at twice the lowest concentration previously reported by Moneke et al. [16] for Acetobacter sp. and Pseudomonas fluorescens isolated from paddy soil, and by Elarabi et al. [28] for Bacillus aryabhattai FACU. A multi-stage dilution of the rhizosphere soil suspension is prepared and used to screen the population of rhizobacteria that degrade glyphosate. One gram of T. diversifolia rhizosphere soil sample was mixed with 9 mL of 0.5% Nutrient Broth (NB) medium containing glyphosate (15 mg mL−1) in a 15 mL test tube. The mixture was then incubated on a rotary shaker at 28 °C and 120 rpm for seven days. As many as 0.1 mL (100 μL) of culture with 0.260 OD at 660 nm was cultured in a petri dish containing 3% Nutrient Agar (NA) medium with 15 mg mL−1 glyphosate and incubated at 28 °C for three days. After the incubation period, colonies that emerge with a variety of shapes and morphological characters are assumed to be glyphosate-resistant bacteria. All colonies of different shapes and colors are then isolated and stored in a medium to be maintained at 4 °C as a stock culture, and their glyphosate-degrading capacity is further verified. Each isolate is coded TBr, which refers to the rhizospheric bacterium from T. diversifolia.
The growth of isolates and glyphosate biodegradation activity
Ten milliliters of 0.5% NB containing 15 mg mL−1 glyphosate was prepared and placed into seven 15 mL volume test tubes to observe the growth of the isolates for a week. For this reason, a 0.1 mL aliquot of OD 0.265 bacterial cultures from previously prepared screened isolates was added to test tubes containing 10 mL of NB medium with 15 mg mL−1 glyphosate and replicated three times. Bacterial growth was tracked daily by taking culture samples, and culture growth was monitored turbidimetrically at 660 nm, following the method used by [24]. Isolates that showed high growth and endurance until the seventh day were selected and further characterized for their ability to degrade glyphosate.
The selected isolates obtained from the previous assay were recultured in different new media using a 0.5% Kingʼs B broth base medium with a glyphosate concentration of 15 mg mL−1. The decrease in glyphosate in the medium was monitored spectrophotometrically according to Jan et al. [29] using a standard prepared with a stock solution of glyphosate (1,000 mg L−1). The same medium was used to test the influence of pH and temperature shifts on growth and the isolateʼs potential to hydrolyze glyphosate. The pH range of the medium is adjusted from 4.0 to 7.0 in steps of 0.5 using 0.5 N HCl solution. The temperature during the incubation is varied between 20 and 40 in 5 °C steps to observe the isolateʼs ability to withstand different environmental conditions.
Phosphorus dissolves and the IAA production assay
Blue molybdate method was utilized to determine inorganic phosphates in liquid media, while ascorbic acid acts as a reducing agent [30]. The color measured spectrometrically at a wavelength of 882 nm was the color intensity. Measurement of indole-3-acetic acid (IAA) production was carried out using Salkowski reagents (1 mL of 0.5 M FeCl3 and 50 mL of 50% HClO4 (v/v) according to Sarwar et al. [31]. The IAA concentrations were measured by readings on a spectrophotometer using a wavelength of 530 nm, and a standard curve was constructed with IAA concentrations (Sigma) expressed in μg mL−1.
An assay of glyphosate as a carbon, phosphorus, and nitrogen source
To further test the ability of isolates to use glyphosate as an energy source and to determine the biodegradation pathway of the isolate, a series of treatments was arranged using the basic medium of Pikovskaya broth, with or without glucose (carbon), calcium phosphate (phosphorus), and nitrogen sources from Bacto. All treatment media contained glyphosate at a concentration of 15 mg mL−1. The combination of treatments assayed were: (A) complete Pikovskaya with glyphosate 15 mg mL−1; (B) Pikovskaya (–carbon,–phosphorus) with glyphosate 15 mg mL−1; (C) Pikovskaya media (–carbon,–nitrogen) with glyphosate 15 mg mL−1; (D) Pikovskaya media (–carbon,–phosphorus,–nitrogen) with glyphosate 15 mg mL−1. Ten milliliters of treatment medium was poured into a 15-mL test tube and inoculated with isolates of TBr1 or TBr12, with three replications. The test tube is incubated on a rotary shaker at 28 °C and 120 rpm for 7 days. The number of populations and the solubility of phosphorus in culture media were observed after 7 days of culture.
The results in this study were not statistically analyzed; therefore, they are presented descriptively to illustrate comparative trends among treatments and isolates.
Morphological and physiological characterization of isolates
The morphological and physiological characterization of the isolate was carried out by observing the color of the colony, its shape, thickness, and physiological properties through Gram staining. Gram staining is used to distinguish between Gram-positive and Gram-negative bacteria, following a standard procedure Krieg et al. [32]. A culture smear is first prepared, then stained with violet crystals, fixed with an iodine solution, rinsed until the color fades, and finally treated with fuchsin dye. The specimen is then dried and observed under a microscope. The results of characterizing the morphology and shape of the cell, as well as its metabolic system, which are commonly used in determining isolates, were subsequently confirmed by molecular identification.
Molecular identification and phylogenetic construction
DNA extraction is carried out using the heat shock method as has been shown to be a simple and efficacious in the DNA extraction of Gram-positive and Gram-negative bacteria [33-35]. Microbial colonies of selected isolates grown on Pikovskayaʼs agar were scraped with a loop and diluted in 200 μL of demineralized water, which was used for DNA extraction by heating at 90 °C for 1 min. After vortexing for 1 min at high speed, the mixture was centrifuged at 12.000 g for 30 s in an Eppendorf microcentrifuge. The supernatant was carefully transferred to a 1.5 mL microcentrifuge tube, and the extracts were stored at –20 °C until further processing. One microliter was pipetted as the template for PCR amplification, using the universal forward (338f) primer with the nucleotide sequence 5'-CCTACGGGAGGCAGCAG-3' and the reverse (518r) primer with the nucleotide sequence 5'-CGTATTACCGCGGCTGCTGG-3'. The 50 μL of PCR reagent mixture consisting of 1 µl of DNA template, 30,75 µl of ddH2O, 10 µl of buffer, 3 µl MgCl2 25mM, 1 µl of mixt PCR dNTP 10 mM, 2 µl of 338f and 518r primer of each, and 0.25 µl Go-Taq. The DNA amplification process is carried out for 20 cycles, with stages: Denaturation at 94 °C for 30 s, primary annealing at 55 °C for 30 s, and extension at 72 °C for 30 s. The PCR results were then run on a 1.5% agarose gel with a voltage of 120 V for 35 min, and then the formed DNA band was examined after being stained with etidium bromide. Direct sequencing of PCR products is carried out using a 518r reverse primer. The base sequences obtained from the sequencing results were searched for homology in the DNA database using the Basic Local Alignment Search Tool (BLAST) on the National Center for Biotechnology Information (NCBI) website (http://www.ncbi.nlm.nih.gov).
The species name of each isolate was then determined based on the results of matching the DNA sequence against the following criteria: The highest percentage of match (query cover), the highest degree of sequence similarity (at least 97%), and the minimum E-value. The trees were reconstructed using the neighbor-joining (NJ) technique [36] and the evolutionary distances were estimated based on 1,000 bootstrap values [37], computed using the Maximum Composite Likelihood method [38]. Evolutionary analyses were conducted in MEGA12 [39] utilizing up to 8 parallel computing threads to create phylogenetic trees between the nearest genetic sequence species obtained after blasting.
Results and discussion
Screening and isolation of glyphosate-degrading rhizobacteria
This study successfully isolated and characterized the rhizospheric bacterium T. diversifolia, which can degrade the glyphosate herbicide at very high concentrations (15 mg mL−1), representing a possible condition of glyphosate residues in tropical lands after multiple herbicide applications. Of the six isolates (TBr1, TBr2, TBr3, TBr5, TBr6 and TBr12) that were able to survive the screening process on various media with a glyphosate content of 15 mg mL−1, three isolates were TBr1, TBr2 and TBr12 -showing the most consistent degradation capacity (Figure 2).

Figure 2 Growth of six isolates in NB containing 15 mg mL−1 of glyphosate. Error bars indicate the standard deviation.
The paucity of isolates obtained is consistent with the findings of [16,24] that only a small fraction of the isolates from the initial screening were completely resistant to high concentrations of glyphosate. Based on the ability to survive in high concentrations of glyphosate, coupled with the ability to produce IAA and dissolve phosphate (Figure 3), which are characterized as Plant Growth Promoting Rhizobacteria, two isolates (TBr1 and TBr12) were subsequently identified and further characterized.

Figure 3 Halozone (arrow direction) of TBr1 (A) and TBr12 (B) isolates formed in Pikovskaya media.
The test results of the effect of pH and temperature variations on the growth of the 2 isolates showed that optimal growth conditions were achieved at a pH range of 6.0 - 7.0 (Figure 4(A)), as indicated by the OD values, which reached 0.910 for the TBr12 isolate and 0.430 for TBr1. Meanwhile, the 2 isolates at temperatures of 30 - 35 °C both reach OD of 0.950 and 0.972 for TBr1 and TBr12, respectively (Figure 4(B)). These conditions represent a general range of tropical soils, thus reinforcing the potential application of both isolates in field bioremediation. The biodegradation of glyphosate is strongly influenced by pH and ambient temperature, as found by Chen et al. [14] and Pipke and Amrhein et al.[40]. Thus, the success of degradation by TBr1 and TBr12 in conditions suitable for tropical agroecosystems lends high applicability value.
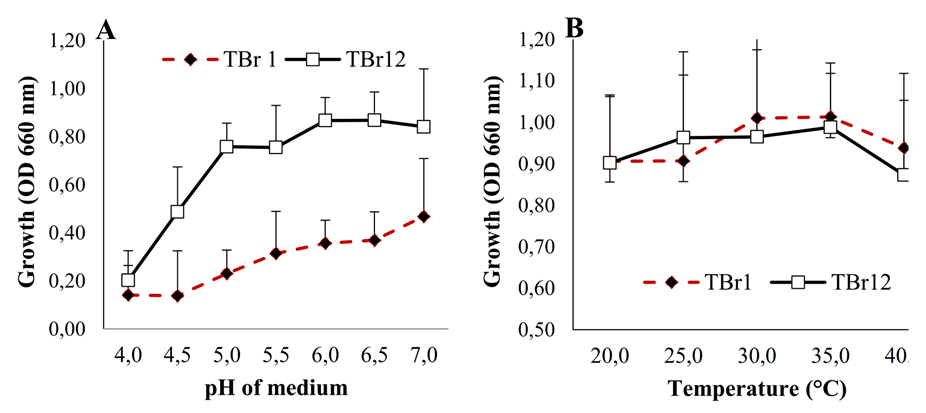
Figure 4 Effect of pH (A) and temperature (B) on the growth of isolates TBr1 and TBr12 in Kingʼs B broth containing 15 mg mL−1 of glyphosate. Error bars indicate the standard deviation.
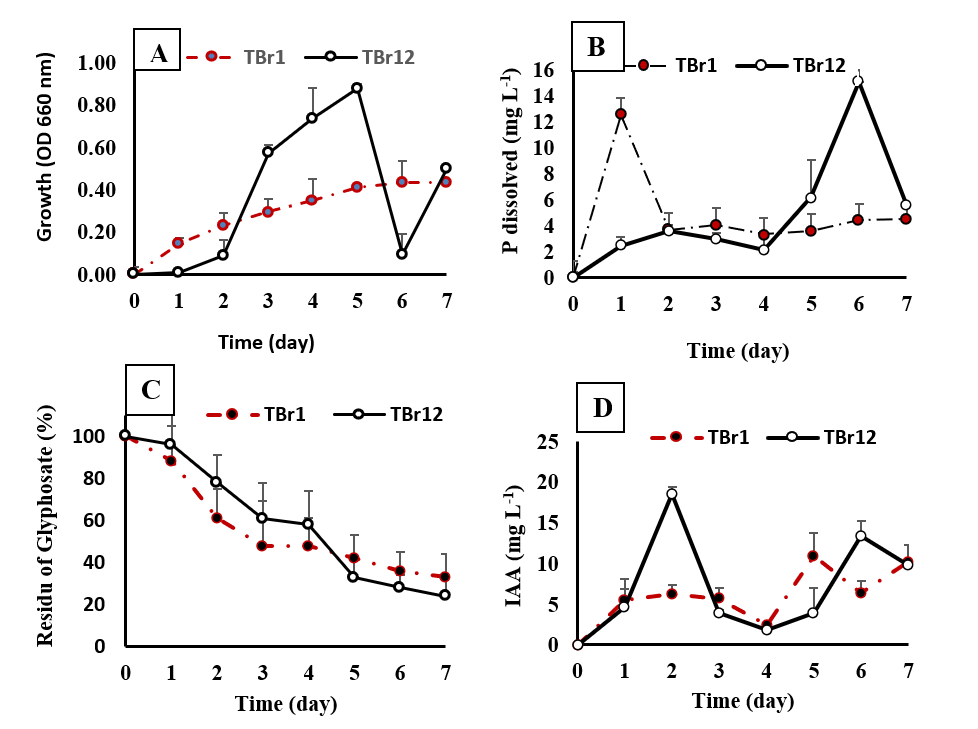
Figure 5 The Growth (A), P-solubilization (B), Glyphosate degradation (C), and IAA production (D) activity of isolates TBr1 and TBr12 during seven days of incubation in Kingʼs B broth containing 15 mg mL−1 of glyphosate. Error bars indicate the standard deviation.
The dissimilarities in morphological and physiological traits of the distinct isolates indeed reflect the differences in the metabolic pathways of glyphosate biodegradation by TBr1 and TBr12. For instance, TBr1 has a shorter lag phase but relatively slower growth rate only reached OD of 0.437 on the 7th day, whereas TBr12 has the opposite with longer lag phase followed by rapid growth after day two until the 5th day with an OD of 0.877. The growth decreased on the 6th day with OD of 0.093 but back up on day seven with OD 0.497 (Figure 5(A)). This growth pattern is indicative of the difference in the 2 isolatesʼ ability to adapt to high glyphosate concentration. The ability of both isolates to survive and grow in a medium composed solely of glyphosate implies that they have employed the pathways of Gox and CP lyase for their biodegradation [15,41]. Besides, the investigation also uncovered the release of inorganic phosphate differing between the 2 isolates (Figure 5(B)). The isolate TBr1 released the largest amount of phosphates on the very first day reaching 12.5 mg L−1 but then steadily decreased the release until the cultureʼs end. In contrast, the isolate TBr12 released phosphates at a slow but constant rate until it reached the peak at 15.1 mg L−1 release on day six. This variation indicates that there are differences in the extent of production of the enzymes Gox and CP lyase by the 2 strains. The findings of Firdous et al. [42] and Ibrahim et al. [17] corroborate the assertion that the bacterial species, type of enzyme as well as the efficiency of the enzymes produced, are the factors determining the rate and the pathway of glyphosate degradation.
When compared to the results of previous studies, the level of resistance of both isolates at a concentration of 15 mg mL−1 of glyphosate far exceeded many earlier reports, for example, Bacillus Albus F9D at 50 mg L−1 [43], Bacillus cereus at 20 mM or equivalent 3.5 mg mL−1 [44] or other isolates that generally survive below 10 mM or equivalent 1.75 mg mL−1 [19]. Nevertheless, the tolerance of isolates TBr1 and TBr12 was lower than that reported for Acetobacter sp. and Pseudomonas fluorescens by [16], as well as for Bacillus aryabhattai FACU by [28], which remained capable of growth at glyphosate concentrations up to 250 mg mL⁻¹. We found that TBr1 and TBr12 isolates showed the fastest growth rates and the ability to break down glyphosate by 67% and 76% in seven days, respectively (Figure 5(C)) at a level of 15 mg mL−1. This uniqueness indicates that both isolates have developed more complex molecular resistance mechanisms. Ecologically, the presence of this bacterium on the rhizosphere of T. diversifolia is also interesting, since this plant is known to thrive in marginal soils with minimal external input [27]; symbiosis with glyphosate-degrading microbes can be an essential adaptation factor. This variation indicates that there are differences in the extent of production of the enzymes Gox and CP lyase by the two strains.
In addition to being capable of breaking down glyphosate, the 2 isolates also continue to produce indole-3-acetic acid (IAA) at a higher glyphosate concentration (Figure 5(D)). Until the 7th day, in the culture media of TBr1 and TBr12 isolates, were still found IAA of 10.16 and 9.80 mg mL−1, respectively. This has physiological significance because glyphosate acts as an inhibitor of the shikimate pathway and, therefore, of the synthesis of tryptophan. Despite this, IAA is still present in the bacterial cultures, indicating that both isolates likely employ alternative routes, such as the tryptamine or indole-3-acetamide pathways, which do not rely on tryptophan [45]. Such a mechanism suggests that the isolate has adapted metabolically to the stress exerted by the herbicide, while also confirming its potential as a rhizobacterial agent that supports plant growth (PGPR) through this duality. In line with Shahid et al. [46], IAA production under environmental stress conditions can be viewed as a bacterial strategy to enhance survival while promoting the growth of host plants.
Utilization of glyphosate as a carbon, phosphorus, and nitrogen source
To evaluate the glyphosate biodegradation pathway used by each isolate, follow-up testing using growing media by removing carbon, phosphorus, and nitrogen sources from the base media used was conducted. The test results showed that both isolates could hydrolyze glyphosate through the CN pathway, producing glioxylic acid and AMPA. In addition, both also utilize the CP pathway to release sarcosine and phosphate in their metabolism. This ability is demonstrated by the growth of both isolates in the growing medium (B) without a source of carbon and phosphorus and (C) without a source of carbon and nitrogen (Figure 6(a)). The growth differences between the 2 isolates in medium (B) and (C) plus in the C, N, and P unsourced media (D), not only approving that the 2 isolates can break the glyphosate chain at the C-P and C-N bonds, but also suggest that they have different CP lyase efficacy in metabolizing glyphosate as demonstrated by the higher dissolved P results (Figure 6(b)) in the culture media of TBr1 compared to TBr12.
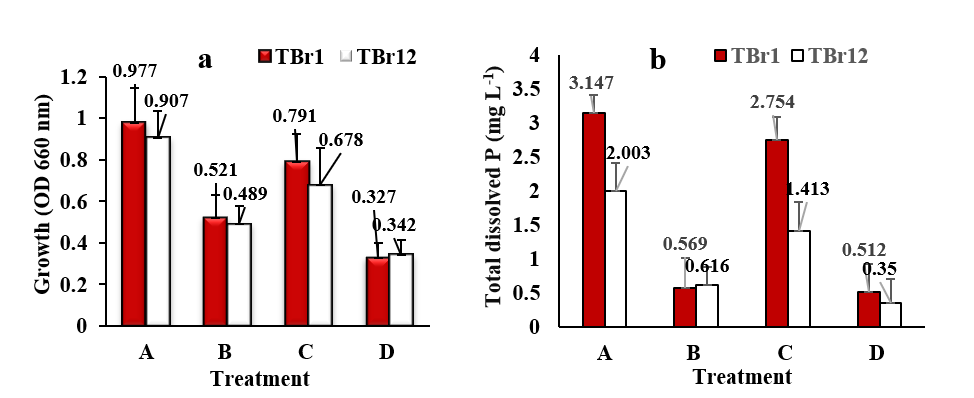
Figure 6 Comparison of population (a) and dissolved P (b) in Pikovskaya broth media of Burkholderia cepacia strain TBr1 and Bacillus velezensis strain TBr12 isolates with treatment: A, Complete Pikovskaya media + glyphosate 15 mg mL−1; B, without carbon and phosphorus sources + glyphosate 15 mg mL−1; C, without carbon and nitrogen sources + glyphosate 15 mg mL−1; D, without carbon, nitrogen, and phosphorus sources + glyphosate 15 mg mL−1 after 7 days culture. Error bars indicate the standard deviation.
TBr1 and Bacillus velezensis strain TBr12 isolates with treatment: A, Complete Pikovskaya media + glyphosate 15 mg mL−1; B, without carbon and phosphorus sources + glyphosate 15 mg mL−1; C, without carbon and nitrogen sources + glyphosate 15 mg mL−1; D, without carbon, nitrogen, and phosphorus sources + glyphosate 15 mg mL−1 after 7 days culture. Error bars indicate the standard deviation.
Based on growth performance and phosphorus release patterns, CP lyase-mediated degradation appears to play a dominant role; however, discrimination between CP lyase I and II remains speculative without targeted enzymatic or genomic analyses, as reported by Sviridov et al. [15] in Achromobacter sp. MPS 12A and Ochrobactrum anthropi GPK3. Thus, TBr1 appears to be more rapidly utilizing glyphosate, whereas TBr12 exhibits a slower but still effective degradation mechanism. The two rhizobacterial isolates obtained are special because very few can use glyphosate as a carbon, nitrogen, and phosphorus source compared to isolates that only metabolize it as a source of nitrogen or phosphorus.
Morphological and physiological characterization
Isolate TBr1 has the characteristics of yellow colour, circular colony shape, mucoid texture, smooth surface, flat edges, raised elevation, and a Gram-negative rod. The isolate TBr12, a Gram-positive, rod-shaped bacterium, exhibits a circular colony shape and smooth-textured colonies. Isolates TBr1 and TBr12 have a small size (Table 1). Both have the colony morphology characteristics of Isolate TBr1 and Isolate TBr12, which resemble those of the corresponding genus.
Table 1 Morphological and physiological characteristics of TBr1 and TBr12 isolates of glyphosate-degrading rhizobacteria from the rhizosphere of T. diversifolia.
Characteristic |
Isolates |
|
|
TBr1 |
TBr12 |
Gram stain |
‒ |
+ |
Cell form |
rod |
rod |
Motility |
+ |
+ |
Cell growth in agar media |
|
|
a)NA |
+ |
+ |
Kingʼs B |
+ |
+ |
b)Pikovskaya |
+/+ |
+/+ |
Malic acid |
‒ |
‒ |
Optimum pH |
6.5 - 7.0 |
6.0 - 6.5 |
Colony morphology |
|
|
Size |
small |
small |
Surface |
Smooth and shiny |
smooth |
Color |
yellow |
white |
Elevation |
raised |
flat |
Sugar use |
glucose |
glucose |
Biochemical tests |
|
|
catalase |
+ |
+ |
oksidase |
+ |
‒ |
glucose fermentation |
‒ |
+ |
IAA |
+ |
+ |
Optimum Temperature |
30 - 35 °C |
30 - 35 °C |
a) + = slow, ++ = fast, +++ = very fast.
b) +/+ = grew/ formed halozone; +/‒ = grew/not formed halozon.
Molecular identification and phylogenetic construction
Identification was carried out molecularly on 2 rhizobacteria isolates (TBr1 and TBr12) with the highest glyphosate degrading activity index. Molecular identification was carried out on the conserved region of ribosomal DNA. The identification of closely related species of isolates TBr1 and TBr12 was performed using BLAST to compare their DNA sequences with those stored in the GenBank database of the National Center for Biotechnology Information (NCBI). The BLAST results showed that the rhizobacteria TBr1 isolate had the closest homology to Burkholderia cepacia strain A21CJ with an identity value of 100%. In comparison, the rhizobacteria TBr12 isolate had the closest homology to Bacillus velezensis strain TH1 with a value of 100% identity (Table 2). Furthermore, the names of the 2 isolates used in this article are Burkholderia cepacia strain TBr1 and Bacillus velezensis strain TBr12.
Table 2 BLAST results of isolates TBr1 and TBr12 based on analysis of rDNA conserve region sequences.
Isolate number |
Taxon identified from Blast |
Max Score |
Total Score |
Query cover |
E-value |
Per. Identity |
Accession |
TBr1 |
Burkholderia cepacia strain A21CJ |
296 |
296 |
76% |
9e-76 |
100% |
EU481409.1 |
TBr12 |
Bacillus velezensis strain TH1 |
298 |
298 |
63% |
3e-76 |
100% |
OQ473003.1 |
Phylogenetic analysis based on 16S rRNA sequences indicates that the B. cepacia strain TBr1 belongs to the B. cepacia complex (Bcc) species (Figure 7). It exhibits the strongest phylogenetic affinity with B. cenocepacia (EU481409.1) and B. multivorans (EF427758.1). High bootstrap support (81%) indicates that this relationship is stable and consistent. The relatively short branch length of this node reflects a higher level of genetic similarity than other nodes in the tree. In contrast, the difference in branch length with the B. cepacia group (0.0065) and the B. contaminans-lata-seminalis group (0.0100) suggests that B. cepacia strain TBr1 is evolutionarily further away from the clade. Thus, B. cepacia strain TBr1 can be characterized as an isolate belonging to the B. cepacia complex (Bcc) group, with the best affinity to the cenocepacia–multivorans subclade.

Figure 7 Neighbor-joining phylogenetic tree constructed from the 16S rDNA sequence of B. cepacia strain TBr1 and related species with close genetic kinship using Clustal W and MEGA 12 software.
Phylogenetic trees constructed using the Neighbor-Joining method showed that B. velezensis strain TBr12 formed separate branches with relatively large branch lengths (0.6781), indicating a significant degree of genetic divergence compared to the main clade of Bacillus (Figure 8). The majority of reference species, including B. velezensis, B. subtilis, B. amyloliquefaciens, B. tequilensis, and B. atrophaeus, form a major clade supported by a bootstrap value of 58%. Although this support is moderate, the grouping of these species is consistent with the kinship relationships reported in the B. subtilis group, which generally exhibit high genomic proximity. The length of the branch to this clade (0.5794) is shorter than in the TBr12 isolate, thus confirming a closer evolutionary relationship among the members of the clade.
The phylogenetic comparison of TBr1 Isolate and TBr12 Isolate is shown in Figures 7 and 8. This figure shows the difference between the strain TBr1 of the neighboring species B. cepacia (Figure 7) and the strain TBr12 of the neighboring species B. velezensis (Figure 8). Previous studies have reported that Bacillus sp. are types of bacteria that can effectively degrade glyphosate [43,44]. Meanwhile, B. cepacia has also been reported as an active degrader of glyphosate in the environment [47]. However, information on B. cepacia and B. velezensis as degraders of glyphosate is still very limited. Therefore, this bacterium presents a significant opportunity in the bioremediation of glyphosate contamination, but a study of its biosecurity aspects is needed before further exploring its potential both at the ex-situ and in-situ levels in restoring glyphosate-soil contaminated areas.

Figure 8 Neighbor-joining phylogenetic tree constructed from the 16S rDNA sequence of B. velezensis strain TBr12 and related species with close genetic kinship using Clustal W and MEGA 12 software.
Conclusions
Pollutant-degrading rhizobacteria are essential, considering microorganisms in the rhizosphere can degrade various contaminants through diverse enzymatic processes. In our study, the rhizobacteria Burkholderia cepacia strain TBr1 and Bacillus velezensis strain TBr12 can live and thrive at the roots of T. diversifolia, effectively degrading glyphosate. The success of glyphosate degradation by both isolates in conditions suitable for tropical agroecosystems (pH 6.0 to 6.5) and temperatures of 30 - 35 °C lends high applicability value. Their ability to adapt to high concentrations of glyphosate and metabolize approximately 67% and 76% of 15 mg mL⁻¹ glyphosate within 7 days of incubation, serving as a source of carbon, nitrogen, and phosphorus, indicates their potential as valuable agents for glyphosate biodegradation. In addition, the ability of the 2 isolates to dissolve phosphates and produce IAA is a unique characteristic for both isolates. Biostimulation or bioaugmentation strategies, combined with T. diversifolia as a hedge-growing plant, could be highly effective for bioremediation in environments contaminated with glyphosate. However, further research is needed to confirm the potential and biosafety of both isolates in bioremediation before field-scale application.
Acknowledgements
The author would like to express his sincere gratitude to the Institute for Research and Community Service of the Faculty of Agriculture, Andalas University, West Sumatra 25163, Indonesia for the Research Grant given to the author (Grant No. 05/SPK/PLK/RKAT Faperta Unand/2025).
Declaration of Generative AI in Scientific Writing
The authors acknowledge the use of generative AI tools (e.g., ChatGPT by OpenAI and Grammarly) in the preparation of this manuscript, specifically for language editing and grammar correction. AI performed no content generation or data interpretation. The authors take full responsibility for the content and conclusions of this work.
CRediT Author Statement
Agustian: Conceptualization, Resources, Methodology, Supervision, Validation, Funding acquisition, and Writing-review & editing. Chairin Nisa: Data curation, Formal analysis, Investigation, Validation, and Visualization. Parwanto: Data curation, Formal analysis, Investigation, Validation, and Visualization. Hermansah: Methodology, Supervision, Validation, and Writing –original draft. Syafrimen Yasin: Methodology, Supervision, Validation, and Writing –original draft.
References
[1] F Maggi, DL Cecilia, FHM Tang and A McBratney. The global environmental hazard of glyphosate use. Science of The Total Environment 2020; 717, 137167.
[2] JP Myers, MN Antoniou, B Blumberg, L Carroll, T Colborn, LG Everett, M Hansen, PJ Landrigan, BP Lanphear, R Mesnage, LN Vandenberg, FS vom Saal, WV Welshons and CM Benbrook. Concerns over use of glyphosate-based herbicides and risks associated with exposures: A consensus statement. Environmental Health 2016; 15, 19.
[3] CM Benbrook. Trends in glyphosate herbicide use in the United States and globally. Environmental Sciences Europe 2016; 28, 3.
[4] PM Dewick. The biosynthesis of shikimate metabolites. Natural Product Reports 1995; 12(6), 579-607.
[5] AR Knaggs. The biosynthesis of shikimate metabolites. Natural Product Reports 1999; 16(4), 525-560.
[6] JT Padilla and HM Selim. Chapter one - Environmental behavior of glyphosate in soils. Advances in Agronomy 2020; 159, 1-34.
[7] AHC van Bruggen, MR Finckh, M He, CJ Ritsema, P Harkes, D Knuth and V Geissen. Indirect effects of the herbicide glyphosate on plant, animal and human health through its effects on microbial communities. Frontiers in Environmental Science 2021; 9, 763917.
[8] P Chávez-Ortiz, Y Tapia-Torres, J Larsen and F Garcia-Olivia. Glyphosate-based herbicides alter soil carbon and phosphorus dynamics and microbial activity. Applied Soil Ecology 2022; 169, 104256.
[9] VL Lozano and HN Pizarro. Glyphosate lessons: Is biodegradation of pesticides a harmless process for biodiversity? Environmental Sciences Europe 2024; 36, 55.
[10] R Kanissery, B Gairhe, D Kadyampakeni, O Batumen and F Alferez. Glyphosate: Its environmental persistence and impact on crop health and nutrition. Plants 2019; 8(11), 499.
[11] JA García-Pérez, E Alarcón, Y Hernández and C Hernandez. Impact of litter contaminated with glyphosate-based herbicide on the performance of Pontoscolex corethrurus, soil phosphatase activities and soil pH. Applied Soil Ecology 2016; 104, 31-41.
[12] Y Sang, JC Mejuto, J Xiao and J Simal-Gandara. Assessment of glyphosate impact on the agrofood ecosystem. Plants 2021; 10(2), 405.
[13] S Singh, V Kumar, JPK Gill, JPK Datta, S Singh, V Dhaka, D Kapoor, AB Wani, DS Dhanjal, M Kumar, SL Harikumar and J Singh. Herbicide glyphosate: Toxicity and microbial degradation. International Journal of Environmental Research and Public Health 2020; 17(20), 7519.
[14] Y Chen, WJ Chen, Y Huang, J Li, J Zhong, W Zhang, Y Zou, S Mishra, P Bhatt and S Chen. Insights into the microbial degradation and resistance mechanisms of glyphosate. Environmental Research 2022; 215(1), 114153.
[15] AV Sviridov, TV Shushkova, NF Zelenkova, NG Vinokurova, IG Morgunov, IT Ermakova and AA Leontievsky. Distribution of glyphosate and methylphosphonate catabolism systems in soil bacteria Ochrobactrum anthropi and Achromobacter sp. Applied Microbiology and Biotechnology 2012; 93, 787-796.
[16] AN Moneke, GN Okpala and CU Anyanwu. Biodegradation of glyphosate herbicide in vitro using bacterial isolates from four rice fields. African Journal of Biotechnology 2010; 9(26), 4067-4074.
[17] NE Ibrahim, V Sevakumaran and F Ariffin. Preliminary study on glyphosate-degrading bacteria isolated from agricultural soil. Environmental Advances 2023; 12, 100368.
[18] ML Castrejón-Godínez, E Tovar-Sánchez, L Valencia-Cuevas, ME Rosas-Ramírez, A Rodríguez and P Mussali-Galante. Glyphosate pollution treatment and microbial degradation alternatives, a review. Microorganisms 2021; 9(11), 2322.
[19] YV Kryuchkova, GL Burygin, NE Gogoleva, YV Gogolev, MP Chernyshova, OE Makarov, EE Fedorov and OV Turkovskaya. Isolation and characterization of a glyphosate-degrading rhizosphere strain, Enterobacter cloacae K7. Microbiological Research 2014; 169(1), 99-105.
[20] G Saxena and RN Bharagava. Bioremediation of industrial waste for environmental safety. Springer, Singapore, 2020.
[21] L Jia, Z Wang, L Ji, SD Neve, PC Struik, Y Yao, J Lv, T Zhou and K Jin. Keystone microbiome in the rhizosphere soil reveals the effect of long-term conservation tillage on crop growth in the Chinese Loess Plateau. Plant and Soil 2022; 473, 457-472.
[22] T Macek, M Macková and J Káš. Exploitation of plants for the removal of organics in environmental remediation. Biotechnology Advances 2000; 18(1), 23-34.
[23] J Montreemuk, TN Stewart and B Prapagdee. Bacterial-assisted phytoremediation of heavy metals: Concepts, current knowledge, and future directions. Environmental Technology & Innovation 2024; 33, 103488.
[24] W Mohy-Ud-Din, MJ Akhtar, S Bashir, HN Asghar, MF Nawaz and F Chen. Isolation of glyphosate-resistant bacterial strains to improve the growth of maize and degrade glyphosate under axenic condition. Agriculture 2023; 13(4), 886.
[25] N Hakim, R Alfina, Agustian, Hermansah and Yulnafatmawita. Bacterial inoculants to increase the biomass and nutrient uptake of Tithonia cultivated as hedgerow plants in ultisols. Malaysian Journal of Soil Science 2014; 18 115-123.
[26] B Jama, CA Palm, RJ Buresh, A Niang, C Gachengo, G Nziguheba and B Amadalo. Tithonia diversifolia as a green manure for soil fertility improvement in western Kenya: A review. Agroforestry Systems 2000; 49, 201-221.
[27] ST Partey. Effect of pruning frequency and pruning height on the biomass production of Tithonia diversifolia (Hemsl) A. Gray. Agroforestry Systems 2011; 83, 181-187.
[28] NI Elarabi, AA Abdelhadi, RH Ahmed, I Saleh, IA Arif, G Osman and DS Ahmed. Bacillus aryabhattai FACU: A promising bacterial strain capable of manipulate the glyphosate herbicide residues. Saudi Journal of Biological Sciences 2020; 27(9), 2207-2214.
[29] MR Jan, J Shah, M Muhammad and B Ara. Glyphosate herbicide residue determination in samples of environmental importance using spectrophotometric method. Journal of Hazardous Materials 2009; 169(1-3), 742-745.
[30] J Murphy and JP Riley. A modified single solution method for the determination of phosphate in natural waters. Analytica Chimica Acta 1962; 27, 31-36.
[31] M Sarwar, M Arshad, DA Martens and WT Frankerberger. Tryptophan-dependent biosynthesis of auxins in soil. Plant and Soil 1992; 147, 207-215.
[32] NR Krieg, JT Staley, DR Brown, BP Hedlund, BJ Paster, NL Ward, W Ludwig and WB Whitman. Bergey’s manual of systematic bacteriology. 2nd ed. Springer, New York, 2010.
[33] B Bearzatto, JF Durant, J Ambroise and JL Gala. Rapid, user-friendly, cost-effective DNA and library Preparation methods for whole-genome sequencing of bacteria with varying cell wall composition and GC content using minimal DNA on the illumina platform. BMC Genomics 2025; 26, 396.
[34] AA Abdulla. Optimization of DNA extraction of Lactobacillus spp for Identification by tuf B gene –Based polymerase chain reaction. Journal of Biology, Agriculture and Healthcare 2014; 4(8), 122-127.
[35] AA Dashti, MM Jadaon, AM Abdulsamad and HM Dashti. Heat treatment of bacteria: A simple method of DNA extraction for molecular techniques. Kuwait Medical Journal 2009; 41(2), 117-122.
[36] N Saitou and M Nei. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Molecular Biology and Evolution 1987; 4(4), 406-425.
[37] J Felsenstein. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985; 39(4), 783-791.
[38] K Tamura, M Nei and S Kumar. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proceedings of the National Academy of Sciences of the United States of America 2004; 101(30), 11030-11035.
[39] S Kumar, G Stecher, M Suleski, M Sanderford, S Sharma and K Tamura. MEGA12 : Molecular evolutionary genetic analysis version 12 for adaptive and green computing. Molecular Biology and Evolution 2024; 41(12), msae263.
[40] R Pipke and N Amrhein. Isolation and characterization of a mutant of Arthrobacter sp. strain GLP-1 which utilizes the herbicide glyphosate as its sole source of phosphorus and nitrogen. Applied and Environmental Microbiology 1988; 54(11), 2868-2870.
[41] N Stosiek, M Talma and M Klimek-Ochab. Carbon-phosphorus Lyase—the state of the art. Applied Biochemistry and Biotechnology 2020; 190, 1525-1552.
[42] S Firdous, S Iqbal and S Anwar. Optimization and modeling of glyphosate biodegradation by a novel Comamonas odontotermitis P2 through response surface methodology. Pedosphere 2020; 30(5), 618-627.
[43] WJ Chen, M Liu, SF Chen, Y Zhang, H Song, MH Abdoulahi, K Bhatt, S Mishra, MA Ghorab, W Zhang and S Chen. Glyphosate bioremediation using a newly isolated Bacillus albus strain F9D: Mechanisms and kinetic studies. Microbial Cell Factories 2025; 24, 195.
[44] J Fan, G Yang, H Zhao, G Shi, Y Geng, T Hou and K Tao. Isolation, identification and characterization of a glyphosate-degrading bacterium, Bacillus cereus CB4, from soil. The Journal of General and Applied Microbiology 2012; 58(4), 263-271.
[45] CL Patten, AJC Blakney and TJD Coulson. Activity, distribution and function of indole-3-acetic acid biosynthetic pathways in bacteria. Critical Reviews in Microbiology 2013; 39(4), 395-415.
[46] M Shahid, B Ahmed and MS Khan. Evaluation of microbiological management strategy of herbicide toxicity to greengram plants. Biocatalysis and Agricultural Biotechnology 2018; 14, 96-108.
[47] R Hertel, K Schöne, C Mittelstädt, J Meißner, N Zschoche, M Collignon, C Kohler, I Friedrich, D Schneider, M Hoppert, R Kuhn, I Schwedt, P Scholz, A Poehlein, M Martienssen, T Ischebeck, R Daniel and FM Commichau. Characterization of glyphosate‐resistant Burkholderia anthina and Burkholderia cenocepacia isolates from a commercial Roundup® solution. Environmental Microbiology Reports 2022; 14(1), 70-84.