Trends
Sci.
2026;
23(9):
12545
Melatonin Attenuates Triple Whammy-Induced Acute Kidney Injury: In Vivo, In Vitro, and In Silico Findings
Rini Indriani Juhardi1, Suriati Usman1, Sang Ayu Iraekawati2,
Yulia Yusrini Djabir2 and Aryadi Arsyad3,*
1Postgraduate Program, Faculty of Pharmacy, Hasanuddin University, Makassar 90245, Indonesia
2Laboratory of Clinical Pharmacy, Faculty of Pharmacy, Hasanuddin University, Makassar 90245, Indonesia
3Department of Physiology, Faculty of Medicine, Hasanuddin University, Makassar 90245, Indonesia
(*Corresponding author’s e-mail: [email protected])
Received: 5 November 2025, Revised: 4 December 2025, Accepted: 15 December 2025, Published: 20 March 2026
Abstract
This study assessed the nephroprotective potential of melatonin in a triple whammy-induced acute kidney injury (AKI) model using in vivo, in vitro, and in silico approaches. Thirty male rats were randomly assigned to 6 groups: Normal control, negative control, N-acetylcysteine (NAC) as a positive control, and 3 melatonin-treated groups (0.5, 1.0 and 1.5 mg/kg). Treatments were administered orally for 5 days, followed by AKI induction via the triple whammy regimen for 2 days. Renal function was evaluated using serum and urine biomarkers, and kidney tissues were examined histologically. In vitro antioxidant assays and molecular docking studies were performed to assess melatonin and NAC interactions with NF-κB and caspase-3. The triple whammy regimen significantly increased renal injury markers and caused structural kidney damage. High-dose melatonin (1.5 mg/kg) significantly improved serum creatinine, urea, and urinary protein levels (p < 0.05) and preserved glomerular and tubular architecture. In vitro, melatonin demonstrated strong antioxidant activity (IC₅₀ = 3.19 µg/mL), comparable to NAC (IC₅₀ = 3.10 µg/mL). Molecular docking revealed moderate to strong binding affinities of melatonin to caspase-3 (−6.0 kcal/mol) and NF-κB (−7.5 kcal/mol). In conclusion, melatonin confers nephroprotection in triple whammy-induced AKI potentially through antioxidant effects and interaction with apoptotic and inflammatory protein targets. Further studies are warranted to evaluate the translational potential of these findings in clinical settings.
Keywords: Acute kidney injury, Antioxidant activity, DPPH, Melatonin, Molecular docking, N-Acetylcysteine, Triple whammy
Introduction
Acute kidney injury (AKI) is a critical global health issue, affecting approximately 13.3 million individuals annually and contributing to around 2 million deaths worldwide [1]. Its prevalence ranges from 1% to 25% with mortality reaching 15% - 60%, and it places a substantial economic burden, accounting for up to 5% of hospital budgets and 1% of total healthcare expenditures [2]. Among the major causes of AKI, ischemia-reperfusion injury and drug-induced nephrotoxicity are well-characterized and are both driven by oxidative stress, inflammation, and apoptosis [3-6].
A well-known drug-related cause is the “triple whammy” combination, which consists of a renin-angiotensin-aldosterone system (RAAS) inhibitor, a non-steroid anti-inflammatory drug (NSAID), and a diuretic. This combination impairs renal autoregulation by reducing circulating volume, blocking prostaglandin-mediated afferent vasodilation, and limiting angiotensin II-mediated efferent arteriolar constriction [7,8]. Clinically, this combination markedly increases AKI risk, particularly in older adults and individuals with dehydration, heart failure, or pre-existing renal dysfunction [9]. A large-scale analysis reported 7,466 of 18,415 documented AKI cases were associated with triple whammy exposure [10].
Melatonin (N-acetyl-5-methoxytryptamine) is a pleiotropic hormone primarily secreted by the pineal gland, known for its role in regulating circadian rhythms and modulating immune responses [11]. Beyond its neuroendocrine functions, melatonin exerts broad systemic effects through its receptors [12]. Melatonin has been known to possess antioxidant and anti-inflammatory effects, acting through ROS reduction, enhancement of antioxidant enzymes, and suppression of proinflammatory cytokines [13,14]. These mechanisms support its nephroprotective activity in various models of ischemic-reperfusion injury and drug-induced nephrotoxicity [15,16].
However, whether melatonin can protect the kidney from damage due to impaired renal autoregulation is still undetermined. Therefore, this study aimed to evaluate the nephroprotective potential of melatonin against triple whammy–induced AKI using in vivo, in vitro, and in silico approaches. N-acetylcysteine (NAC), a thiol antioxidant and glutathione precursor, was used as a positive control due to its well-recognized nephroprotective effects [17,18].
Materials and methods
Drugs and chemicals
Melatonin supplements were sourced from SWANSON® distributors (Indonesia). The drug combination used to induce triple whammy-associated acute kidney injury consisted of ibuprofen 400 mg (TRIFA, Indonesia), ramipril 5 mg (NOVELL, Indonesia), and injectable furosemide (BERNOFARM, Indonesia). N-acetylcysteine (SAMPHARINDO, Indonesia) was obtained from licensed local pharmacies. Laboratory chemicals, including 70% ethanol, diethyl ether, and 10% neutral buffered formalin, were supplied by a certified chemical distributor in Makassar, Indonesia. 2,2-diphenyl-1-picrylhydrazyl (DPPH) for antioxidant analyses was purchased from Sigma Chemical Co. (USA).
Animal preparation
Male Wistar rats 8 - 10 weeks old and weighing 180 - 220 g (n = 30) were used in this study. The animals were housed in polycarbonate cages under controlled environmental conditions (22 ± 2 °C temperature, 55 ± 10% relative humidity, and a 12-hour light/dark cycle), with free access to a standard laboratory diet and drinking water ad libitum. Animals were acclimatized to the laboratory environment for 1 week prior to the initiation of the experiment to minimize stress-related variability. The study protocol was reviewed and approved by the Ethics Committee of the Faculty of Pharmacy, Hasanuddin University, Indonesia (Ethical Approval No. 1086/UN4.17/KP.06.05/2025).
Triple whammy combination
The triple whammy regimen consisted of ibuprofen (124 mg/kg body weight), ramipril (0.5 mg/kg), and furosemide (3.72 mg/kg). Ibuprofen and ramipril were suspended in 0.5% sodium carboxymethyl cellulose (Na. CMC) and administered orally twice daily, while furosemide was given via intraperitoneal injection once daily. The selected doses were determined based on preliminary experiments, which successfully induced AKI in rats while avoiding excessive systemic toxicity.
Experimental protocols
A total of 30 male Wistar rats were randomly assigned into 6 experimental groups (n = 5 per group): A normal control receiving vehicle only (Na CMC); a negative control receiving triple whammy administration without pre-treatment; a positive control pre-treated with NAC 41 mg/kg; and 3 melatonin pre-treated groups receiving 0.5 mg/kg (low dose), 1.0 mg/kg (medium dose), or 1.5 mg/kg (high dose) of melatonin. Melatonin doses were determined using standard interspecies conversion from typical human doses of 5 - 15 mg/day. Similarly, the NAC dose was calculated from the standard 400 mg/day human regimen. All pre-treatments (melatonin or NAC) were administered once daily for 5 consecutive days. On days 6 and 7, triple whammy drugs were administered to induce AKI. On day 8, animals were anesthetized for sample collection; blood was obtained via the orbital vein for serum urea and creatinine analysis, and urine samples were collected for evaluation of renal function markers.
Blood and urine analysis
Blood samples were centrifuged at 2,500 rpm for 30 min to obtain serum. Serum urea and creatinine were measured using commercial reagent kits following the manufacturer’s instructions, and analyzed with a semi-automated clinical chemistry analyzer (Humalyzer® 4000, Human Diagnostics, Germany). Urine samples were collected individually from each rat using metabolic cages. The collected urine was immediately analyzed using a Verify® U-120 urine analyzer (Urit Medical, China) to assess urinary protein levels.
Histopathological examination
Following euthanasia with cervical dislocation, the kidneys were surgically removed and immediately rinsed with normal saline to remove residual blood. The organs were subsequently fixed in 10% neutral buffered formalin for 48 h to preserve tissue architecture. Post-fixation, the kidneys were sectioned longitudinally and processed into paraffin blocks using standard histological techniques. Paraffin-embedded tissues were cut into 4 - 5 µm sections using a rotary microtome (Leica RM2235, Germany) and floated on a 40 °C water bath to flatten the sections. The sections were then mounted on glass slides and air-dried for 2 h at room temperature. Hematoxylin and eosin (H&E) staining was performed to evaluate general histological morphology. Histopathological evaluation was conducted under a light microscope (Olympus CX43, Japan) at a magnification of 40×. Representative photomicrographs were captured using a calibrated digital camera attached to the microscope. Blinded assessment of histological changes was performed by an experienced pathologist.
Antioxidant activity assay
The antioxidant activity of melatonin and N-acetylcysteine was evaluated using the DPPH radical scavenging assay. Stock solutions (1 mg/mL) were prepared in methanol, and serial dilutions (10 - 50 ppm) were tested in triplicate. In a 96- well plate, 100 μL of DPPH solution was mixed with 100 μL of each sample concentration. After incubation, absorbance was measured at 517 nm using a microplate reader. The percentage of radical scavenging was calculated, and IC₅₀ values were determined using Microsoft Excel.
Molecular docking procedure
The in-silico analysis was conducted to compare the binding potential of melatonin and NAC toward key inflammatory and apoptotic regulators of AKI. The 3D structures of melatonin (PubChem CID: 896) and NAC (PubChem CID: 12035) were retrieved from PubChem database. NF-κB (PDB ID: 4DN5) and caspase-3 (PDB ID: 3DEI) were selected as target proteins based on prior literature identifying them as central mediators of inflammation and apoptosis in AKI models [19,20]. Their crystallographic structures were obtained from the RCSB Protein Data Bank.
Protein and ligand preparation were performed using UCSF Chimera’s Dock Prep module, which included removal of crystallographic water molecules, addition of polar hydrogens, and assignment of Gasteiger charges. A validation step was carried out through redocking of native ligands into their respective binding sites, and RMSD values were calculated to confirm protocol reliability. RMSD values were 1.041 Å for NF-κB and 2.641 Å for caspase-3, both within acceptable ranges for docking reproducibility [21].
For NF-κB, the docking grid box was set to 28×12×20 points (XYZ) with center coordinates of −9.333, 29.083 and −4.444. For caspase-3, the grid dimensions were 30×30×30 points, centered at −46.620, 15.373, −22.195. Docking simulations were performed using AutoDock Vina integrated in UCSF Chimera. The exhaustiveness parameter was set to 8, which is the standard and recommended level for balanced speed and accuracy. A total of 10 docking poses (num_modes = 10) were generated for each ligand–protein pair, and the pose with the lowest binding energy was selected for further analysis.
Binding affinities (kcal/mol) were extracted from Vina output, where more negative values indicate stronger interactions. Key interactions, including hydrogen bonds, hydrophobic contacts, and proximity to catalytic residues, were visualized and analyzed using BIOVIA Discovery Studio Visualizer.
Statistical analysis
All results are expressed as mean ± standard deviation (SD). Statistical analyses were performed using GraphPad Prism version 10.4.2. Data normality was assessed using the Shapiro-Wilk test. For normally distributed data, differences between groups in biomarker levels were analyzed using 1-way analysis of variance (ANOVA), followed by Tukey’s post hoc test. Non-parametric data, including urinalysis were analyzed using the Kruskal-Wallis test, followed by Dunn’s Multiple comparison test for post hoc comparisons. A p-value of < 0.05 was considered statistically significant.
Results and discussion
Renal biomarkers
Acute kidney injury was successfully induced by the triple whammy drug combination, as reflected by marked elevations in serum creatinine and urea levels (Figure 1). The normal control group showed baseline creatinine and urea values of 1.08 ± 0.24 mg/dL and 51.5 ± 7.18 mg/dL, respectively. In contrast, rats receiving triple whammy combination exhibited more than a 2-fold increase in serum creatinine (2.34 ± 0.46 mg/dL) and a nearly 6- fold rise in serum urea (310.6 ± 57.85 mg/dL), confirming significant renal impairment. This pattern aligns with the known mechanism of triple whammy–induced AKI, in which combined hemodynamic disturbances compromise glomerular filtration and precipitate acute renal dysfunction [7,8].
Proteinuria further supported the occurrence of kidney injury. While urinary protein was undetectable in the normal group, the AKI group showed measurable protein levels (0.72 ± 0.38 mg/dL), indicating glomerular and tubular damage. This finding is consistent with reports identifying protein leakage as an early marker of nephron injury driven by oxidative stress, inflammation, and disruption of the filtration barrier [22,23].
Pretreatment with melatonin at the lower doses (0.5 and 1.0 mg/kg) did not significantly reduce serum creatinine, urea, or urinary protein levels, suggesting insufficient renal protection at these doses. In contrast, melatonin at 1.5 mg/kg markedly improved kidney function, with creatinine and urea values reduced to 1.41 ± 0.26 and 91.0 ± 16.1 mg/dL, respectively. Urinary protein was also nearly undetectable (0.06 ± 0.08 mg/dL), approaching levels observed in the normal control. These findings support a dose-dependent protective effect of melatonin, consistent with its known antioxidant, anti-inflammatory, and mitochondrial-stabilizing actions [24,25].
Interestingly, NAC did not significantly improve renal outcomes in this study, which possibly reflects both dose-related and model-specific factors. Although NAC is widely recognized for its antioxidant and glutathione-restoring properties [26], its renoprotective efficacy is highly dose-dependent and varies across AKI models [27,28]. The dose used in this study was selected through human-to-rat conversion from the commonly administered 400 mg/day regimen. however, attempts to increase the dose were not feasible because NAC-pretreated rats exhibited gastrointestinal bleeding after triple-whammy exposure. This prevented further dose escalation and limited our ability to evaluate NAC across a therapeutic range.
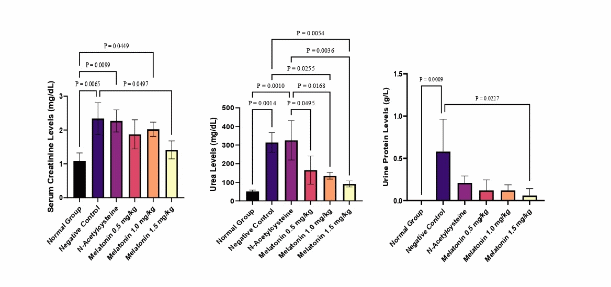
Figure 1 Comparison of serum and urinary biomarker levels among treatment groups. a) Creatinine, b) Urea, c) Urine Protein levels.
Renal histopathological changes
The kidneys of the normal control group exhibited well-preserved architecture, characterized by normal glomeruli, intact Bowman’s capsules, and organized tubular structures, confirming physiological renal morphology (Figures 2(a) and 2(b)). In contrast, rats in the negative control group demonstrated classical features of acute kidney injury, including Bowman’s capsule dilatation, tubular protein casts, interstitial hemorrhage, and inflammatory cell infiltration (Figures 2(c) and 2(d)). These morphological alterations are consistent with previous reports describing glomerular and tubular damage resulting from ischemic and oxidative stress-induced injury [29].
NAC treatment conferred partial histological protection, as evidenced by the attenuation of inflammatory cell infiltration and improved tubular preservation (Figure 2e-f). However, the persistence of glomerular dilatation and protein casts suggests that NAC’s antioxidant effect was not sufficient to fully prevent structural damage. Lower doses of melatonin (0.5 and 1.0 mg/kg) failed to confer significant histological protection, as indicated by persistent glomerular and tubular damage (Figures 2(g) - 2(j)). Conversely, treatment with 1.5 mg/kg melatonin markedly restored renal architecture, with the absence of glomerular dilatation, tubular casts, or hemorrhagic lesions (Figures 2(k) and 2(l)). These findings are consistent with melatonin’s known ability to preserve histological integrity through the suppression of oxidative stress, inflammation, apoptosis, and sirtuin (SIRT) family in renal tissues [30-32]. Melatonin’s lipophilic nature allows it to penetrate cellular and mitochondrial membranes [33], where it scavenges reactive oxygen species and stabilizes mitochondrial function. This feature of melatonin is more likely to underlie the improved histological and biochemical profiles observed at the highest dose.
\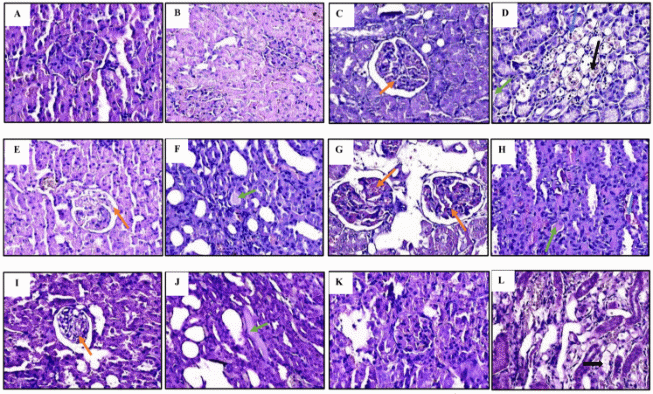
Figure 2 Histopathological features of kidney tissue. (A,B) normal control; (C,D) negative control; (E,F) NAC 41 mg/kg; (G,H) melatonin 0.5 mg/kg; (I,J) melatonin 1.0 mg/kg; (K,L) melatonin 1.5 mg/kg. Indicators of acute kidney injury include dilated Bowman’s capsules (red arrows), tubular protein casts (green arrows), inflammatory cells (blue arrows), and tubular degeneration (black arrows).
Predicted interaction with molecular targets
The docking analysis evaluated melatonin’s interaction with NF-κB and caspase-3, 2 central regulators of inflammation and apoptosis in AKI. Ligand interactions, including hydrogen bonding and key residue contacts, are summarized in Tables 1 and 2 and visualized in Figures 3 and 4.
For NF-κB, the native ligand AGS showed the strongest binding (−8.9 kcal/mol). Melatonin exhibited a comparatively strong affinity (−7.5 kcal/mol) and formed a hydrogen bond with Lys429, a residue involved in stabilizing the p65/p50 dimer and its DNA-binding interface [34]. Additional non-covalent interactions with Ala427, Val414, Leu406, and Cys533 indicate that melatonin engages residues important for maintaining NF-κB structural integrity. This interaction profile suggests that melatonin may hinder NF-κB activation and thereby dampen downstream inflammatory signaling. These computational results align with experimental evidence showing that melatonin inhibits IκBα degradation and p65 nuclear translocation, thereby reducing expression of inflammatory mediators such as TNF-α, IL-6, and COX-2 in kidney injury models [35].
NAC demonstrated weaker affinity for NF-κB (−4.2 kcal/mol), forming only limited interactions with Lys429 and Asn576. This is consistent with the established view that NAC modulates NF-κB activity mainly through redox-dependent pathways rather than direct protein binding [36]. Its minimal receptor engagement may partly explain the modest biological effects observed in this study.
For caspase-3, melatonin showed moderate affinity (−6.0 kcal/mol) relative to the native ligand (−8.1 kcal/mol). It formed multiple hydrogen bonds with Arg207, Arg64, Ser205, and Gly165 near the catalytic pocket, suggesting potential interference with caspase-3 activation [19]. Interactions with His121 and Met61, residues adjacent to the catalytic cysteine, further support the possibility of steric or allosteric modulation. These findings are consistent with prior in vivo studies reporting reduced caspase-3 expression and activity following melatonin treatment in renal injury models [37,38].
In comparison, NAC displayed weaker binding to caspase-3 (−4.6 kcal/mol), forming fewer hydrogen bonds and engaging less critical residues. This supports the notion that NAC’s anti-apoptotic activity occurs mainly through restoration of redox balance rather than direct inhibition of apoptotic enzymes [39,40].
Table 1 Binding affinities and interacting amino acid residues of melatonin and N-acetylcysteine with the NF-κB protein.
Compounds |
Binding Affinity (kcal/mol) |
Hydrogen Bond Residues |
Other Bonds/Interaction Residues |
Melatonin |
−7.5 |
Lys429 |
Ala427, Val414, Cys533, Leu522, Leu406, Gln479, Met469 |
N-Acetylcsytein |
−4.2 |
Lys429, Asn576 |
Asp534 |
Native ligand (AGS) |
−8.9 |
Leu406, Leu472, Glu470, Phe411, Gly412, Gly409, Ser410 |
Lys429, Lys517, Met469, Asp519, Ala427, Leu522, Cys533, Val414 |
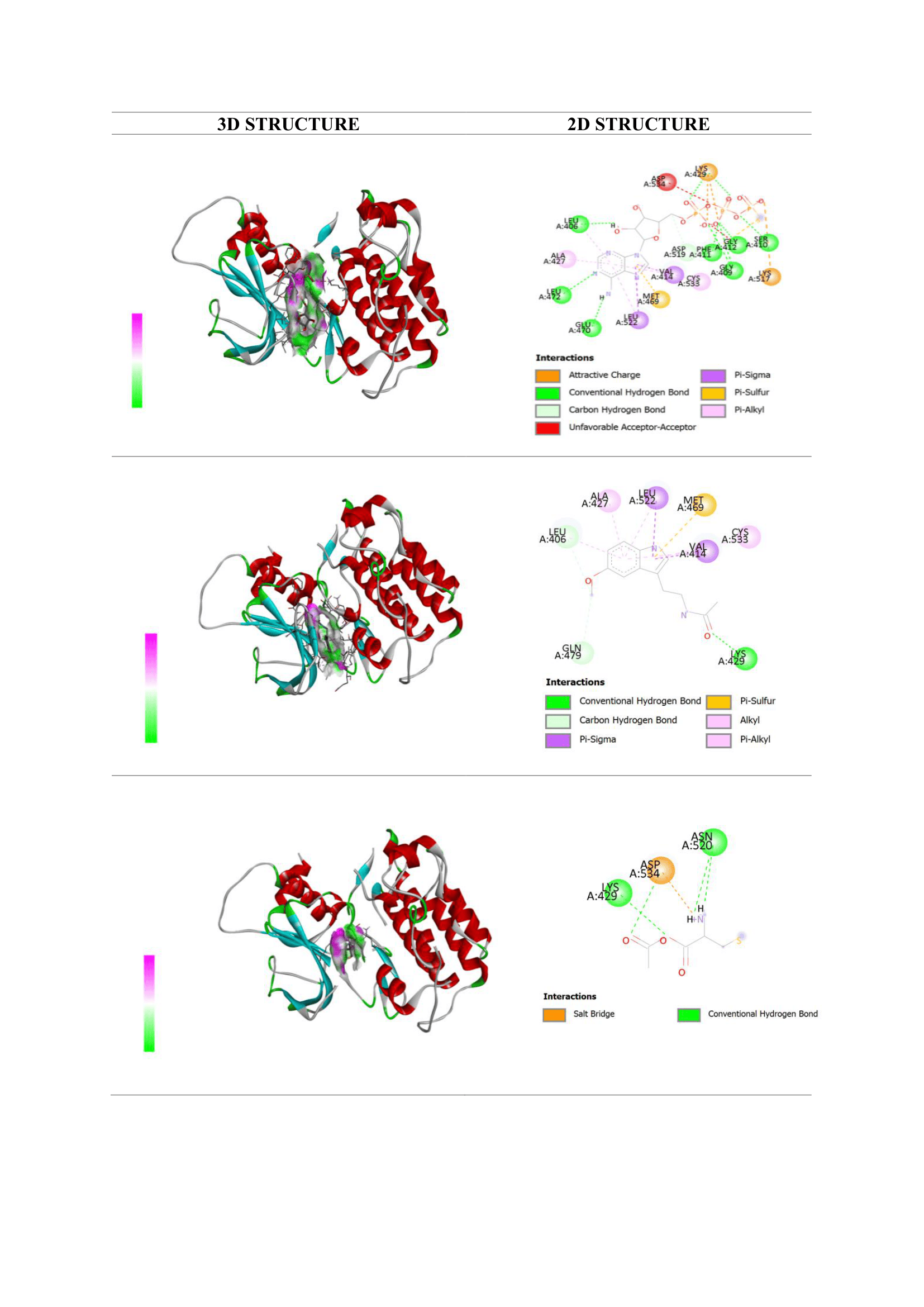
Figure 3 Visualization of molecular docking interactions with the NF-κB receptor: (a) native ligand AGS; (b) melatonin; and (c) N-acetylcysteine (NAC).
Table 2 Binding affinities and interacting amino acid residues of melatonin and N-acetylcysteine with the Caspase-3 protein.
Compounds |
Binding Affinity (kcal/mol) |
Hydrogen Bond Residues |
Other Bonds/Interaction Residues |
Melatonin |
−6.0 |
Arg207, Arg64, Ser205, Gly165 |
His121, Met61 |
N-Acetylcsytein |
−4.6 |
Gly122, Ser205 |
His121, Tyr204, Gly165 |
Native ligand (RXB) |
−8.1 |
Tyr204, Ser209 |
Arg207, Phe256, Trp206 |
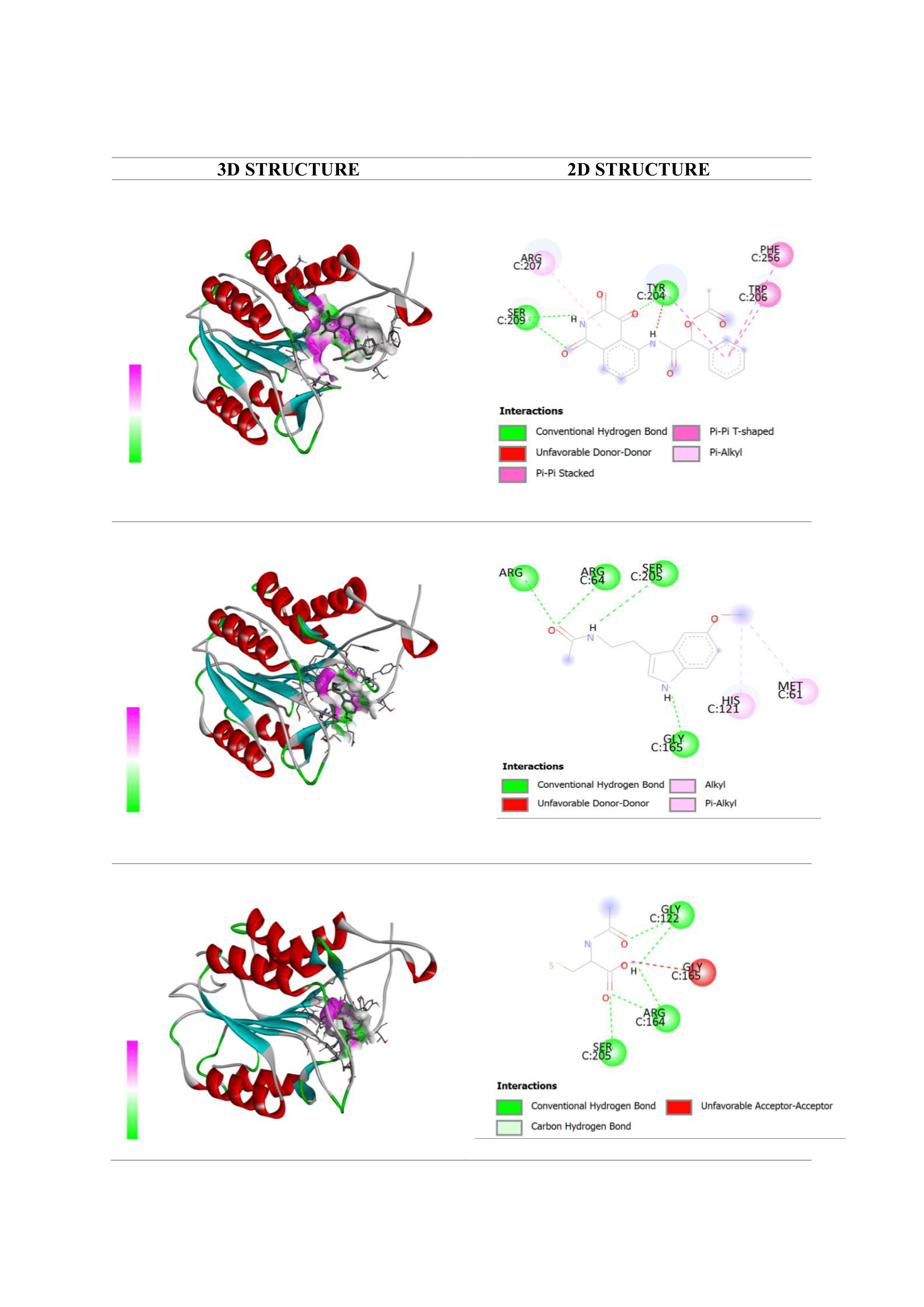
Figure 4 Visualization of molecular docking interactions with the caspase-3 protein: (a) native ligand AGS; (b) melatonin; and (c) N-acetylcysteine (NAC).
In vitro antioxidant activity
Based on DPPH radical scavenging assay, melatonin exhibited an IC₅₀ of 3.19 µg/mL, while N-acetylcysteine showed an IC₅₀ of 3.10 µg/mL (Table 3). Both melatonin and NAC demonstrated very strong antioxidant activity, with IC₅₀ < 50 µg/mL, indicating strong radical scavenging activity [41]. The antioxidant capacity of melatonin is consistent with its renoprotective effects, particularly in mitigating oxidative stress through multiple mechanisms. Melatonin has been shown to reduces ROS production in a sepsis-induced AKI model [13].
Table 3 The comparison between radical scavenging activities of Melatonin and N-Acetylcysteine.
Sample |
IC50 (µg/ml) |
R2 |
Equation |
Melatonin |
3.19 |
0.9497 |
y = 6.3219x + 37.617 |
N-Acetylcsytein |
3.10 |
0.9656 |
y = 18.917x + 27.439 |
Additionally, melatonin enhances endogenous antioxidant defenses by upregulating key enzymes such as superoxide dismutase, glutathione reductase, and glutathione peroxidase, while also lowering lipid peroxidation [42]. Moreover, melatonin can chelate transition metals involved in the Fenton and Haber- Weiss reactions, thereby inhibiting the formation of highly reactive hydroxyl radicals and further reducing oxidative damage [25].
Several studies have demonstrated pleiotropic activities of melatonin, highlighting its antioxidant, anti- inflammatory, and anti-apoptotic properties [43]. In cisplatin-induced AKI models, melatonin demonstrated nephroprotection by inhibiting both apoptosis and necroptosis [35]. Similarly, in sepsis-induced AKI models, melatonin reduced renal damage by modulating ROS levels [13]. Consistent with these findings, melatonin's nephroprotective effect was believed due to its ability to neutralize free radicals at the subcellular level and regulate inflammatory cytokines, restoring the balance between apoptosis and cell survival [16].
This study highlights melatonin’s renoprotective potential through multiple mechanisms, including potent free radical scavenging and inhibition of the NF-κB inflammatory cascade and caspase-3-mediated apoptosis. These effects support its promise as a therapeutic agent against drug-induced acute kidney injury, particularly under triple whammy conditions. However, limitations such as the short-term evaluation and translational gap between animal models and human pathology warrant further investigation.
Limitation
Several limitations should be considered when interpreting the findings of this study. The relatively small sample size may restrict the robustness of the statistical comparisons and limits the strength of the conclusions. Additionally, the short duration of observation allowed us to capture only the acute phase of kidney injury; therefore, potential long-term effects of melatonin or NAC could not be assessed.
Another important limitation is the use of a single NAC dose. Although the selected dose was based on clinical conversion, attempts to explore a broader dose range were not feasible due to gastrointestinal bleeding observed in NAC-pretreated rats following triple-whammy exposure. This restricted our ability to fully evaluate NAC’s therapeutic profile and may partly explain its limited renoprotective effect in this model.
Histopathological evaluation was performed descriptively, without a quantitative scoring system. While the images clearly demonstrated structural improvements, the absence of semi-quantitative scoring reduces objectivity. Future studies should incorporate validated semiquantitative or quantitative scoring methods to make direct comparison across treatment groups.
Finally, as with all animal studies, the rat model cannot fully replicate the complexity of human AKI, especially in the context of polypharmacy and comorbidities. Thus, while the findings offer meaningful insights, they should be interpreted with caution, and further studies with larger cohorts, longer follow-up, multiple dosing regimens, and standardized histological scoring are needed to confirm and extend these results.
Conclusions
Triple whammy-induced AKI represents a relevant model for studying nephrotoxicity. Administration of high-dose melatonin showed a tendency to ameliorate renal injury, as evidenced by improved biomarkers and histological protection of glomerular and tubular structures. These findings were supported by in vitro and in silico studies, which indicated the antioxidant potential and affinity of melatonin for key regulators of inflammation and apoptosis (NF-κB and caspase-3).
The results were interpreted cautiously due to limitations, including the small sample size, short observation period, and the use of a rat model. The renoprotective effect was primarily observed at high doses, and thus optimal dosing and safety require further investigation. Overall, melatonin shows potential for protecting against renal injury, but further studies are needed before considering clinical application.
Acknowledgements
The authors are grateful to the Metabolism and Oxidative Stress Research Group (MOST RG) for providing financial support to complete the pilot study [TRG Grant 00773/UN4.22/PT.01.03/2025]. The authors would like to thank Putri Nurfauziah and Muh. Arfandy Gunawan for their assistance in the molecular docking study.
Declaration of generative AI in scientific writing
During the preparation of this work, the authors used ChatGPT (OpenAI) for grammar checking and language refinement. The final content, interpretations, and conclusions are entirely the authors’ own and have been critically reviewed and approved by all co-authors.
CRediT author statement
Rini Indriani Juhardi: Writing Original draft, Formal analysis, Resources, Visualization. Suriati Usman: Investigation, Project administration. Sang Ayu Iraekawati: Investigation. Yulia Yusrini Djabir: Conceptualization, Supervision, and Writing-Review & Editing. Aryadi Arsyad: Conceptualization, Methodology, Supervision, and Writing-Review & Editing.
References
EAJ Hoste, JA Kellum, NM Selby, A Zarbock, PM Palevsky, SM Bagshaw, SM Bagshaw, SL Goldstein, J Cerdá and
LS Chawla. Global epidemiology and outcomes of acute kidney injury. Nature Reviews Nephrology 2018; 14, 607-625.
M Kerr, M Bedford, B Matthews and D O’Donoghue. The economic impact of acute kidney injury in England. Nephrology, dialysis, transplantation: Official publication of the European Dialysis and Transplant Association. European Renal Association 2014; 29(7), 1362-1368.
MØ Nørgård and P Svenningsen. Acute kidney injury by ischemia/reperfusion and extracellular vesicles. International Journal of Molecular Sciences 2023; 24(20), 15312.
YL Wu, HF Li, HH Chen and H Lin. MicroRNAs as biomarkers and therapeutic targets in inflammation- and ischemia- reperfusion-related acute renal injury. International Journal of Molecular Sciences 2020; 21(18), 6738.
YY Djabir, AArsyad, U Usmar, E Wahyudin, H Arwi and IS Rupang. The stages of development of liver and renal injuries in rats induced by fixed dose combination of antituberculosis regimen. FABAD Journal of Pharmaceutical Sciences 2020; 45, 29-35.
A Fiqardina, YY Djabir, A Santoso, NS Salsabil and I Ismail. The nephroprotective effect of clove oil (Oleum caryophylli) against levofloxacin toxicity in rats. Iranian Journal of Toxicology 2022; 16(1), 27-34.
KK Loboz and GM Shenfield. Drug combinations and impaired renal function - the ‘triple whammy’. The Journal of Clinical Pharmacology 2025; 59(2), 239-243.
DM Calvo, LC Saiz, L Leache, MC Celaya, M Gutiérrez‐Valencia, A Alonso, J Erviti, N Alzueta, A Echeverría, J Garjón, C Fontela, L Sanz, MT Acín, ML Fernández and N Gómez. Effect of the combination of diuretics, renin-angiotensin-aldosterone system inhibitors, and non-steroidal anti-inflammatory drugs or metamizole (triple whammy) on hospitalisation due to acute kidney injury: A nested case-control study. Pharmacoepidemiology and Drug Safety 2023; 32(8), 898-909.
MB Alsobaie and L Alsheikh. Incidence of the triple whammy phenomenon among cardiovascular diseases patients in Saudi Arabia and awareness among healthcare professionals. Frontiers Nephrology 2025; 5, 1494459.
Y Kunitsu, D Hira, A Morikochi, T Ueda, T Isono, S Morita and T Terada. Time until onset of acute kidney injury by combination therapy with “Triple Whammy” drugs obtained from Japanese Adverse Drug Event Report database. PLoS One 2022; 17(2), e0263682.
N Patel, XP Huang, JM Grandner, LC Johansson, B Stauch, JD McCorvy, Y Liu, B Roth and V Katritch. Structure-based discovery of potent and selective melatonin receptor agonists. Elife 2020; 9, e53779.
A Ariyandy, MA Arsyad, ARA Hasyar, MM Abdullah, N Nawir, AD Astuti, MI Basri, R Renaldi, ME Rachman, AZ Surur, W Usman and Fadliana Utami. Dynamic levels of hormonal, oxidants, insomnia, and stress in badminton athletes who practice morning and evening. Journal of Human Sport Exercise 2024; 19(2), 522-535.
L Chen, Z Han, Z Shi, C Liu and Q Lu. Melatonin alleviates renal injury in mouse model of sepsis. Frontiers in Pharmacology 2021; 12, 697643.
ML Wang, CH Wei, WD Wang, JS Wang, J Zhang and JJ Wang. Melatonin attenuates lung ischaemia-reperfusion injury via inhibition of oxidative stress and inflammation. Interact Cardiovascular Thoracic Surgery 2018; 26(5), 761-767.
R Dun, T Lan, J Tsai, J Mao, Y Shao, X Hu, W Zhu, G Qi and Y Peng. Protective effect of melatonin for renal ischemia- reperfusion injury: a systematic review and meta-analysis. Frontiers in Physiology 2022; 12, 791036.
Z Raza and Z Naureen. Melatonin ameliorates the drug induced nephrotoxicity: Molecular insights. Nefrologia 2020; 40(1), 12-25.
SH Jo. N-acetylcysteine for prevention of contrast-induced nephropathy: A narrative review. Korean Circulation Journal 2011; 41(12), 695.
P Yu, J Luo, H Song, T Qian, X He, J Fang, W Dong and X Bian. N-acetylcysteine ameliorates vancomycin-induced nephrotoxicity by inhibiting oxidative stress and apoptosis in the in vivo and in vitro models. International Journal of Medical Sciences 2022; 19(4), 740.
B Yang, S Lan, M Dieudé, JP Sabo-Vatasescu, A Karakeussian-Rimbaud, J Turgeon, S Qi, L Gunaratnam, N Patey, and MJ Hébert Caspase-3 is a pivotal regulator of microvascular rarefaction and renal fibrosis after ischemia-reperfusion injury. Journal of The American Society of Nephrology 2018; 29(7), 1900-1916.
L Markó, E Vigolo, C Hinze, JK Park, G Roël, A Balogh, M Choi, A Wübken, J Cording, IE Blasig, FC Luft, C Scheidereit, KM Schmidt-Ott, R Schmidt-Ullrich and DN Müller. Tubular epithelial Nf-κb activity regulates ischemic AKI. Journal of American Society of Nephrology 2016; 27(9), 2658-2669.
D Ramireza and J Caballero. Is it reliable to take the molecular docking top scoring position as the best solution without considering available structural data? Molecules 2018; 23(5), 1038.
D Zeng, B Wang, Z Xiao, X Wang, X Tang, X Yao, P Wang, M Li, Y Dai and X Yu. Early diagnosis and treatment of kidney injury: A focus on urine protein. International Journal of Molecular Sciences 2024; 25(20), 11171.
JE Toblli, P Bevione, F Di Gennaro, L Madalena, G Cao and M Angerosa. Understanding the mechanisms of proteinuria: Therapeutic implications. International Journal of Nephrology 2012; 2012(1), 546039.
P Loren, R Sánchez, ME Arias, R Felmer, J Risopatrón and C Cheuquemán. Melatonin scavenger properties against oxidative and nitrosative stress: Impact on gamete handling and in vitro embryo production in humans and other mammals. International Journal of Molecular Sciences 2017; 18(6), 1119.
KKAC Monteiro, ME Shiroma, LL Damous, MDJ Simões, RDS Simões, J Cipolla-Neto, EC Baracat and JM Soares. Antioxidant actions of melatonin: A systematic. Antioxidants 2024; 13(4), 439.
EY Hernández-Cruz, OE Aparicio-Trejo, FA Hammami, D Bar-Shalom, M Tepel, J Pedraza-Chaverri and A Scholze. N- acetylcysteine in kidney disease: Molecular mechanisms, pharmacokinetics, and clinical effectiveness. Kidney International Reports 2024; 9(10), 2883-2903.
DM Small, WY Sanchez, SF Roy, C Morais, HL Brooks, JS Coombes and DW Johnson. N-acetyl-cysteine increases cellular dysfunction in progressive chronic kidney damage after acute kidney injury by dampening endogenous antioxidant responses. American Journal of Physiology-Renal Physiology 2018; 314(5), F956-F968.
SN Heyman, M Goldfarb, A Shina, F Karmeli and S Rosen. N-acetylcysteine ameliorates renal microcirculation: Studies in rats. Kidney International 2003; 63(2), 634-641.
JV Bonventre and L Yang. Cellular pathophysiology of ischemic acute kidney injury. The Journal of Clinical Investigation 2011; 121(11), 4210-4221.
K Sevgin, T Sapmaz, E Erdem, S Topkaraoglu, KB Baki and S Aras. Comparative analysis of melatonin's protective role against flattening filter and flattening filter-free X-ray induced nephropathy: Focus on apoptosis and SIRT1 modulation. BMC Nephrology 2025; 26(1), 444.
TC Sun, XC Liu, SH Yang, LL Song, SJ Zhou, SL Deng, L Tian and LY Cheng. Melatonin inhibits oxidative stress and apoptosis in cryopreserved ovarian tissues via nrf2/ho-1 signaling pathway. Frontiers in Molecular Biosciences 2020; 7, 163.
A Atasever, S Teki̇n, I Bolat, M Bolat, Y Dağ, E Sengul, S Yıldırım, M Warda and F Çelebi. The effects of melatonin on oxidative stress, inflammation, apoptosis and Nrf2/HO-1 in acrylamide-induced lung injury in rats. Naunyn-Schmiedeberg’s Archives of Pharmacology 2025; 398, 15905-15922.
H Yu, EJ Dickson, SR Jung, DS Koh and B Hille. High membrane permeability for melatonin. The Journal of General Physiology 2016; 110(3), 605a.
AK Mankan, MW Lawless, SG Gray, D Kelleher and R McManus. NF-kappaB regulation: The nuclear response. Journal of Cellular and Molecular Medicine 2009; 10(1), 405-455.
JW Kim, J Jo, JY Kim, M Choe, M, J Leem and JH Park. Melatonin attenuates cisplatin-induced acute kidney injury through dual suppression of apoptosis and necroptosis. Biology 2019; 8(3), 64.
J Milara, F Martinez-Exposito, P Montero, I Roger, MA Bayarri, P Ribera, MN Oishi-Konari, JR Alba-García, E Zapater and J Cortijo. N-acetylcysteine reduces inflammasome activation induced by SARS-CoV-2 proteins in vitro. International Journal of Molecular Sciences 2023; 23(23), 14518.
YC Chang, SY Hsu, CC Yang, PH Sung, YL Chen, TH Huang, GS Kao, SY Chen, KH Chen, HJ Chiang, HK Yip and F Lee. Enhanced protection against renal ischemia- reperfusion injury with combined melatonin and exendin-4 in a rodent model. Experimental Biology and Medicine 2016; 241(14), 1588-1602.
S Barangi, R Asadi, S Mehri and G Karimi. The effect of melatonin on benzo(a)pyrene-induced renal toxicity in mice. Toxicology and Industrial Health 2023; 39(5), 248-257.
B Pedre, U Barayeu, D Ezeriņa and TP Dick. The mechanism of action of N-acetylcysteine (NAC): The emerging role of H2S and sulfane sulfur species. Pharmacology & Therapeutics 2021; 228, 107916.
Y Liu, K Liu, N Wang and H Zhang. N‑acetylcysteine induces apoptosis via the mitochondria‑dependent pathway but not via endoplasmic reticulum stress in H9c2 cells. Molecular Medicine Reports 2017; 16(5), 6626-6633.
I Gulcin and SH Alwasel. DPPH radical scavenging assay. Processes 2023; 11(8), 2248.
D Li, JH Pan, XF Huang,YQ Liao, YJ Ling and JY Luo. Effect of melatonin on oxidative stress indicators in animal models of fibrosis: A systematic review and meta-analysis. Free Radical Biology and Medicine 2023; 195, 158-177.
D Mahmood. Pleiotropic effects of melatonin. Drug Research 2019; 69(02), 65-74.