Trends
Sci.
2026;
23(9):
12909
Diversity of Bioactive Compounds from Sphagnum junghuhnianum and Their Antimicrobial Potential: An In-Vitro and In-Silico Assessment
Sinta R Pardosi, Etti Sartina Siregar* and Isnaini Nurwahyuni
Department of Biology, Faculty of Mathematics and Natural Sciences, Universitas Sumatera Utara,
Medan 20155, Indonesia
(*Corresponding author’s e-mail: [email protected])
Received: 6 December 2025, Revised: 15 January 2026, Accepted: 22 January 2026, Published: 30 March 2026
Abstract
Sphagnum junghuhnianum is a moss species that has attracted attention as a potential natural source of antimicrobial agents. This study aimed to evaluate the antimicrobial activity of the methanolic extract of S. junghuhnianum and to explore its chemical profile using LC-HRMS combined with in silico approaches. Antimicrobial activity was assessed using disk diffusion assays against Gram-positive and Gram-negative bacteria, as well as pathogenic fungi. The extract exhibited notable antibacterial activity in preliminary screening, particularly against Streptococcus pyogenes (42.01 ± 0.33 mm) and Staphylococcus aureus (34.62 ± 2.21 mm), while no antifungal activity was observed under the tested conditions. LC-HRMS profiling revealed 514 putatively identified metabolites, of which 35 compounds were selected for further biological activity prediction and molecular docking analysis. PASS prediction suggested that several compounds may possess antimicrobial relevance. Molecular docking against Penicillin-Binding Protein 1 (PBP1) indicated that ursolic acid showed the most favorable binding affinity (−9.8 kcal/mol) compared to the reference ligand, suggesting a possible antibacterial mode of action. Overall, this study provides preliminary evidence supporting the antibacterial potential of S. junghuhnianum and highlights its relevance for future investigations involving quantitative bioassays and compound validation.
Keywords: Antimicrobial activity, Bioactive metabolites, Molecular docking, Penicillin-Binding Protein 1, Sphagnum junghuhnianum
Introduction
Bryophytes, particularly mosses belonging to the genus Sphagnum, represent an ecologically important plant group that plays a crucial role in maintaining ecosystem stability, especially in wetlands and humid forest environments. In addition to functioning as natural water reservoirs and long-term carbon sinks, Sphagnum species are known to produce a wide range of secondary metabolites that contribute to their ecological resilience. Previous studies have reported that Sphagnum contains diverse classes of bioactive compounds, including polysaccharides such as sphagnan, amino acids, carotenoids, fatty acids, triterpenes, sterols, and phenolic acids [1,2]. Identified phenolic acids include dihydroxybenzoic, gallic, vanillic, salicylic, caffeic, chlorogenic, p-coumaric, and
cinnamic acids [3]. In addition, Sphagnum species contain unique flavonoids, including flavonols and flavanones, present in both aglycone and glycosylated forms [4]. The dominant phenolic compound, sphagnic acid, plays an important ecological role in peat formation by inhibiting microbial decomposition and lowering environmental pH [5,6], while sphagnan exhibits cation-exchange properties that influence nutrient retention in peatland ecosystems [7]. Several studies have further demonstrated that phenolics, flavonoids, and saponins in Sphagnum are associated with antimicrobial activity [8,9].
Sphagnum junghuhnianum is a moss species widely distributed in humid tropical regions, including Indonesia. Despite its ecological significance, studies focusing on its chemical composition and biological properties remain limited. Previous investigations have reported that ethanol extracts of S. junghuhnianum exhibit antimicrobial activity against selected Gram-positive bacteria and fungi, suggesting the presence of secondary metabolites with potential antimicrobial relevance [10]. More recent studies have indicated that S. junghuhnianum exhibits biochemical activity related to enzyme production and secondary metabolite biosynthesis, further supporting its value as a subject for chemical and biological exploration [11]. However, comprehensive characterization of its metabolite profile remains scarce.
To date, no studies have examined the bioactive constituents of S. junghuhnianum from Indonesia, particularly from North Sumatra. Most existing research on Sphagnum species has focused primarily on taxonomy rather than microbiological or biochemical characterization. Given the growing threats to tropical forests caused by deforestation and other anthropogenic pressures, the investigation of bioactivity properties in S. junghuhnianum has become increasingly urgent and relevant.
Penicillin-Binding Protein 1 (PBP1) from Staphylococcus aureus is a key transpeptidase involved in the final stages of bacterial cell wall biosynthesis. The Protein Data Bank structure 7O4B provides detailed structural information on the enzyme’s active site and its interaction with β-lactam antibiotics, which inhibit transpeptidase activity by acylating the catalytic serine residue [12]. This structural model has been widely used to investigate protein–ligand interactions and to explore mechanisms associated with β-lactam resistance in S. aureus, including methicillin-resistant strains (MRSA) [13]. Consequently, PBP1 represents a relevant molecular target for computational screening approaches aimed at identifying compounds with potential antibacterial relevance [14]. Natural secondary metabolites derived from mosses, including S. junghuhnianum, may possess structural features that enable interaction with this enzyme, although such interactions remain to be experimentally validated [15].
This study aims to investigate the metabolite profile of Sphagnum junghuhnianum using LC-HRMS and to evaluate its antimicrobial activity through preliminary in vitro screening. In addition, in silico approaches, including PASS prediction and molecular docking, were employed to explore the potential biological relevance of selected metabolites and their predicted interactions with bacterial protein targets. These computational analyses are intended to generate hypotheses and guide future experimental studies, rather than to establish definitive mechanisms or therapeutic applications. Overall, this work seeks to contribute to the growing body of knowledge on Sphagnum-derived metabolites and their possible relevance in antimicrobial research.
Materials and methods
Research materials and tools
Preparation of Sphagnum junghuhnianum extracts
A total of 2,000 g of Sphagnum junghuhnianum samples were collected from the Sicike-cike Nature Tourism Park, North Sumatra, Indonesia (coordinates: 2.652462°N, 380944°E). The samples were thoroughly rinsed with distilled water to remove soil particles and other impurities, then air-dried at room temperature for 7 - 10 days. The dried material was cut into small fragments and ground into a fine powder, yielding 100 g of dried sample. The powdered material (100 g) was placed into an Erlenmeyer flask and extracted with 3,000 mL of 70% methanol. The flask was sealed with plastic film and secured with a rubber band to prevent solvent evaporation. Maceration was carried out for 72 h at room temperature with occasional stirring. After extraction, the mixture was filtered, and the combined filtrates were concentrated under reduced pressure at 40 °C using a rotary evaporator, producing a viscous methanolic extract of S. junghuhnianum.

Figure 1 Population of Sphagnum junghuhnianum on the substrate.
Antibacterial assays of Sphagnum junghuhnianum extracts
Pathogens used for antibacterial activity
The antimicrobial activity of the extract was tested against selected pathogenic microorganisms, including Gram-positive and Gram-negative bacteria, as well as fungal strains. The test microorganisms included three Gram-positive bacteria: Staphylococcus aureus ATCC 25923, Streptococcus pyogenes ATCC 35041, and Streptococcus mutans ATCC 35668; and three Gram-negative bacteria: Klebsiella pneumoniae ATCC 700603, Salmonella typhi ATCC 14028, and Porphyromonas gingivalis ATCC 33277. In addition, two fungal strains were included, Candida albicans ATCC 32476 and Aspergillus niger ATCC 793772.
Antibacterial activity
The evaluation of antibacterial efficacy was conducted using the disc diffusion method [16]. Twenty mL of Mueller-Hinton Agar (MHA) medium was dispensed into sterile Petri dishes and allowed to solidify Bacterial suspensions were adjusted to the turbidity of a 0.5 McFarland standard and uniformly spread on the agar surface using sterile cotton swabs. Filter paper disks were impregnated with 100 µL methanolic extract of S. Junghuhnianum and placed onto the inoculated agar surface. Chloramphenicol served as the positive control, whereas DMSO was used as the negative control. Plates were incubated at room temperature for 24 h. After incubation, inhibition zone diameters were measured. Although Minimum Inhibitory Concentration (MIC) values were not determined in this study, the disk diffusion assay provided preliminary evidence of antibacterial activity. All antimicrobial assays were performed in triplicate, and the results are presented as mean ± standard deviation. Although minimum inhibitory concentration (MIC) values were not determined in this study, the disk diffusion assay provided preliminary qualitative evidence of antibacterial activity.
Antifungal and anti-candida activity
Antifungal assays were conducted using the disk diffusion method on Potato Dextrose Agar (PDA). A total of 20 mL of PDA medium was poured into Petri dishes and allowed to solidify. Candida albicans suspensions were standardized to the McFarland turbidity scale and spread evenly across the agar surface. Disks loaded with 100 µL of methanolic extract were placed onto the medium. Plates were incubated at room temperature for 48 h. Following incubation, inhibition zones were measured to assess antifungal activity against C. albicans and A. niger.
Liquid Chromatography-high Resolution Mass Spectrometry (LC-HRMS) analysis
Metabolite profiling of S. junghuhnianum extract was performed using LC-HRMS. Compound separation was carried out using a Thermo Scientific™ Vanquish™ Horizon UHPLC system equipped with an Accucore™ Phenyl Hexyl column (100×2.1 mm2, 2.6 µm). The mobile phase consisted of water with 0.1% formic acid (A) and acetonitrile with 0.1% formic acid (B), applied at a flow rate of 0.3 mL/min. A gradient elution from 5% to 90% B was performed over 25 min. Column temperature was maintained at 40 °C, and the injection volume was 5 µL. Detection was performed using a Thermo Scientific™ Orbitrap™ Exploris 240 HRMS in Full MS/dd-MS² mode under both positive and negative ionization modes. The mass resolution was set at 60,000 FWHM, with a scan range of 70 - 800 m/z and collision energies between 30 - 70 NCE. Ionization was conducted using a Heated Electrospray Ionization (H-ESI) source at spray voltages of 3,500 V (positive mode) and 2,500 V (negative mode), with a transfer tube temperature of 300 °C and a vaporizer temperature of 320 °C. For sample preparation, 50 mg of extract was dissolved in 1 mL of HPLC-grade methanol, vortexed for 1 min, sonicated for 30 min, centrifuged at 1,400×g for 5 min, and filtered through a 0.2 µm membrane prior to injection. Data acquisition and processing were carried out using Thermo Scientific™ Compound Discoverer 3.3 software with compound identification based on mzCloud, MassList Database, and ChemSpider libraries.
In-silico analysis
Protein-ligan preparation
The bioactive compounds identified from Sphagnum junghuhnianum and used for this study are listed below along with their respective PubChem Compound Identifiers (CID): 4-(3-Dodecanyl)benzenesulfonic acid (CID: 29249), Gluconic acid (CID: 10690), 4-Methoxycinnamaldehyde (CID: 641294), Guttiferone E (CID: 5352088), Dilaurylmethylamine (CID: 76205), Ursolic acid (CID: 64945), 6-Gingerol (CID: 442793), Bilobetin (CID: 5315459), Xylarioic acid B (CID: 46832764), Brosimacutin C (CID: 10936790), Ceriporic acid C (CID: 9819908), Glycerophospho-N-palmitoyl ethanolamine (CID: 53393933), Garcinol (CID: 5490884), (−)-Desoxygambogenin (CID: 16078252), 3,4-Dihydroxyphenylacetic acid (CID: 547), 32-Hydroxy-ent-guttiferone M (CID: 139031624), Nootkatone (CID: 1268142), Questiomycin C (CID: 146682678), (+)-ar-Turmerone (CID: 160512), Polygodial (CID: 72503), Ginkgetin (CID: 139587180), Caerulomycin F (CID: 25192237), Momilactone (CID: 162644), 18β-Glycyrrhetinic acid (CID: 10114), Punctaporonin K (CID: 139584880), Aqabamycin A (CID: 46846125), Malyngolide dimer (CID: 46211780), Pulcherriminic acid (CID: 3083664), Mimosamycin (CID: 4198), Asperglaucide (CID: 10026486), Helquinoline (CID: 10466080), 5-O-Desosaminyl-6-O-methylerythronolide A (CID: 10579285), and Carbazomycin B (CID: 166449).
Prediction of biological activity using PASS online
The biological activities of the selected compounds were predicted using the Prediction of Activity Spectra for Substances (PASS) tool available on the Way2Drug server (http://way2drug.com/PassOnline/). The SMILES structures of each compound were obtained from PubChem (https://pubchem.ncbi.nlm.nih.gov) and submitted to the server for activity prediction. The results were interpreted based on the Pa (probability to be active) values, with the following criteria: Pa > 0.70 indicates high probability of exhibiting biological activity. 0.50 ≤ Pa ≤ 0.70 indicates moderate to low probability of activity. Pa < 0.50: Low or negligible probability of activity. These predictions were used solely for hypothesis generation.
Drug-likeness profiles of the compounds were evaluated using SwissADME (http://www.swissadme.ch/) according to Lipinski’s Rule of 5, including molecular weight, hydrogen bond donors and acceptors, and lipophilicity. This evaluation was applied as an early-stage screening tool, not as confirmation of oral bioavailability.
Molecular docking analysis
The study employed Autodock Vina within PyRx 8.0.0 to perform docking analysis, treating the protein as the macromolecule and examining bioactive compounds and capivasertib as ligands. Docking utilized specific grid parameters centered at (1.3057×1.4901×0.3121) with dimensions (36.7569×19.5379×18.1727 Å). Visualization of the docking outcomes was accomplished using Discovery Studio 2024 software. The docking analysis was conducted as a supportive, hypothesis-generating approach to explore potential ligand-target interactions and does not constitute experimental confirmation of inhibitory activity.
Antimicrobial activity of Sphagnum junghuhnianum
Based on Table 1, the methanolic extract of Sphagnum junghuhnianum exhibited antibacterial activity in disk diffusion screening against several pathogenic microorganisms, indicating its potential as a source of antibacterial agents. The largest inhibition zone was observed against S. pyogenes ATCC 35041 (42.01 ± 0.33 mm) (Figure 2), reflecting notable antibacterial activity under the tested conditions. This value was higher than the inhibition zones recorded for S. aureus ATCC 25923 (34.62 ± 2.21 mm) and S. mutans ATCC 65064 (11.53 ± 0.11 mm), suggesting differential susceptibility among Gram-positive bacteria. Although the antibacterial effect of the extract remained lower than that of the reference antibiotic chloramphenicol, the observed inhibition zones indicate that S. junghuhnianum contains secondary metabolites with potential antibacterial relevance. These findings are consistent with previous reports describing mosses as sources of bioactive compounds capable of inhibiting bacterial growth.
The comparatively lower inhibition observed against S. mutans suggests reduced susceptibility relative to other Gram-positive bacteria. This may be associated with intrinsic factors such as differences in cell wall architecture, biofilm-forming capacity, or other protective mechanisms that limit the effectiveness of certain antimicrobial compounds. Similar patterns have been reported previously, where extracts from moss species, including S. junghuhnianum, showed stronger antibacterial activity against selected Gram-positive bacteria than against other tested strains [17].
Among Gram-negative bacteria tested, inhibition was detected only against P. gingivalis, with inhibition zones of 9.03 ± 0.03 mm (50% extract) and 9.73 ± 0.51 mm (100% extract). No measurable inhibition was detected against K. pneumoniae, indicating limited antibacterial activity against Gram-negative bacteria overall. This observation is consistent with the known resistance mechanisms of Gram-negative bacteria, particularly the presence of an outer lipopolysaccharide membrane that restricts the penetration of many antimicrobial compounds [18].
In contrast, the methanolic extract did not exhibit any antifungal or anti-yeast activity. No inhibition was observed against A. niger or C. albicans, while the positive control (ketoconazole) produced clear inhibition zones of 10.22 ± 0.07 mm and 17.16 ± 0.05 mm, respectively. These results indicate that the active metabolites in S. junghuhnianum are likely bactericidal or bacteriostatic primarily targeting Gram-positive bacteria rather than broad-spectrum antimicrobial compounds
These findings are supported by [19], who demonstrated that moss-derived secondary metabolites such as phenolics, flavonoids, and tannins are closely associated with antimicrobial properties. Similarly, [20] reported that flavonoids inhibit bacterial growth through mechanisms such as membrane disruption, enzyme inhibition, and protein binding. Furthermore, the phenolic and flavonoid content of S. junghuhnianum has been shown to vary with ecological conditions such as altitude [21], suggesting that environmental factors at the collection site may influence antimicrobial potency.
Table 1 Antimicrobial activity of the methanolic extract of Sphagnum junghuhnianum.
Extract |
Test microorganisms |
Inhibition zone (mm) |
Positive control |
|||
25% |
50% |
100% |
Chloramphenicol (1000 ppm)/Nystatin (10,000 mg/L) |
|||
Sphagnum junghuhnianum |
Gram-negative bacteria |
Porphyromonas gingivalis ATCC 33277 |
0 |
9.03 ± 0.03 |
9.73 ± 0.51 |
18.05 ± 0.41 |
|
|
Klebsiella pneumoniae ATCC 700603 |
0 |
0 |
0 |
12.15 ± 0.68 |
|
|
Salmonella typhi ATCC 14028 |
0 |
0 |
0 |
22.36 ± 0.84 |
|
Gram-positive bacteria |
Streptococcus mutans ATCC 65064 |
0 |
0 |
0 |
20.37 ± 0.59 |
|
|
Streptococcus pyogenes ATCC 35041 |
0 |
11.98 ± 1.08 |
17.02 ± 2.58 |
42.01 ± 0.33 |
|
|
Staphylococcus aureus ATCC 25923 |
0 |
8.96 ± 0.32 |
9.99 ± 0.71 |
34.62 ± 2.21 |
|
Fungi |
Aspergillus niger ATCC 793772 |
0 |
0 |
0 |
10.22 ± 0.07 (Nystatin) |
|
Yeast |
Candida albicans ATCC 32476 |
0 |
0 |
0 |
17.16 ± 0.05 (Nystatin) |
C
B
A![]()
![]()
![]()
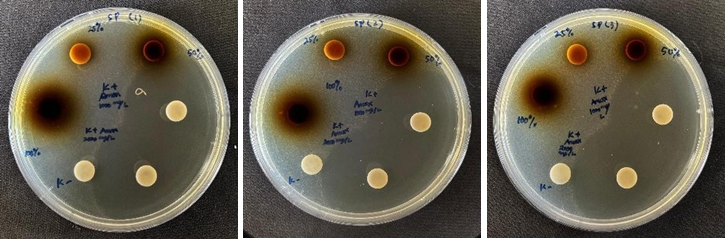
Figure 2 Inhibition zones of the methanolic extract of Sphagnum junghuhnianum against S. pyogenes ATCC 35041 (replicates 1, 2, and 3).
LC-HRMS Analysis
Bioactive compounds of Sphagnum junghuhnianum Dozy & Molk
LC-HRMS profiling of S. junghuhnianum Dozy & Molk revealed a total of 514 putatively annotated metabolites, indicating a high level of chemical diversity. To explore compounds with potential biological relevance, particularly those associated with antimicrobial-related activities, a targeted compound selection approach was applied. This selection was based on literature surveys and database mining using KEGG, ChEBI, PubChem, and NPAtlas. Compounds previously reported to exhibit antibacterial, antifungal, or general antimicrobial activities were prioritized for further computational evaluation. A total of 35 compounds met the criteria and were selected for further in silico screening. Selection criteria included MS/MS fragmentation data, peak intensity (relative abundance), and association with known biosynthetic pathways of antimicrobial metabolites. The selected compounds include: 4-(3-Dodecanyl)benzenesulfonic acid (CID: 29249), Gluconic acid (CID: 10690), 4-Methoxycinnamaldehyde (CID: 641294), Guttiferone E (CID: 5352088), Dilaurylmethylamine (CID: 76205), Ursolic acid (CID: 64945), 6-Gingerol (CID: 442793), Bilobetin (CID: 5315459), Xylarioic acid B (CID: 46832764), Brosimacutin C (CID: 10936790), Ceriporic acid C (CID: 9819908), Glycerophospho-N-palmitoyl ethanolamine (CID: 53393933), Garcinol (CID: 5490884), (−)-Desoxygambogenin (CID: 16078252), 3,4-Dihydroxyphenylacetic acid (CID: 547), 32-Hydroxy-ent-guttiferone M (CID: 139031624), Nootkatone (CID: 1268142), Questiomycin C (CID: 146682678), (+)-ar-Turmerone (CID: 160512), Polygodial (CID: 72503), Ginkgetin (CID: 139587180), Caerulomycin F (CID: 25192237), Momilactone (CID: 162644), 18-β-Glycyrrhetinic acid (CID: 10114), Punctaporonin K (CID: 139584880), Aqabamycin A (CID: 46846125), Malyngolide dimer (CID: 46211780), Pulcherriminic acid (CID: 3083664), Mimosamycin (CID: 4198), Asperglaucide (CID: 10026486), Helquinoline (CID: 10466080), 5-O-Desosaminyl-6-O-methylerythronolide A (CID: 10579285), and Carbazomycin B (CID: 166449). Retention times for these compounds are presented in Table 2, while the chromatographic profile is shown in Figure 3.
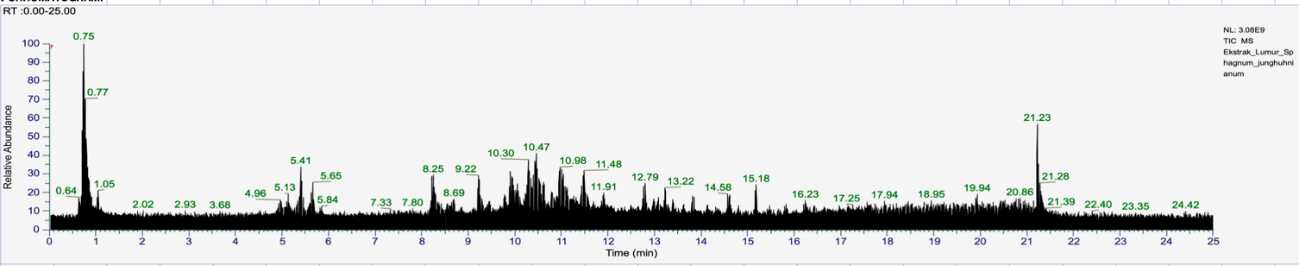
Figure 3 Chromatogram of LC-HRMS analysis of Sphagnum junghuhnianum Dozy & Molk.
Table 2 Identified compounds from Sphagnum junghuhnianum Dozy & Molk. based on LC-HRMS analysis.
Compound name |
Mol. Formula |
R. time (min) |
Smiles |
PubChem ID |
4-(3-Dodecanyl) Benzenesulfonic Acid |
C18H30O3S |
10.613 |
CCCCCCCCCC(CC)C1=CC=C(C=C1)S(=O)(=O)O |
|
Gluconic Acid |
C6 H12 O7 |
0.719 |
C(C(C(C(C(C(=O)O)O)O)O)O)O |
|
4-Methoxycinnamaldehyde |
C10 H10 O2 |
11.697 |
COC1=CC=C(C=C1)/C=C/C=O |
|
Guttiferone E |
C38 H50 O6 |
15.323 |
CC(=CCC1CC2(C(=O)C(=C(C3=CC(=C(C=C3)O)O)O)C(=O)C(C2=O)(C1(C)C)CC=C(C)C)CC(CC=C(C)C)C(=C)C)C |
|
Dilaurylmethylamine |
C25H53 N |
14.929 |
CCCCCCCCCCCCN(C)CCCCCCCCCCCC |
|
Ursolic Acid |
C30H48 O3 |
12.79 |
CC1CCC2(CCC3(C(=CCC4C3(CCC5C4(CCC(C5(C)C)O)C)C)C2C1C)C)C(=O)O |
|
6-Gingerol |
C17H26O4 |
10.165 |
CCCCCC(O)CC(=O)CCc1ccc(O)c(OC)c1 |
|
Bilobetin |
C31H20O10 |
8.781 |
COC1=C(C=C(C=C1)C2=CC(=O)C3=C(C=C(C=C3O2)O)O)C4=C(C=C(C5=C4OC(=CC5=O)C6=CC=C(C=C6)O)O)O |
|
Xylarioic acid B |
C11H22O5 |
6.75 |
CCC(C)C(C(C)(C(C(C)C(=O)O)O)O)O |
|
Brosimacutin C |
C20H22O5 |
11.695 |
CC(C)(CCC1=C(C=CC2=C1OC(CC2=O)C3=CC=C(C=C3)O)O)O |
|
Ceriporic acid C |
C21H36 O4 |
1 |
CCCCCCCC/C=C\CCCCCCC(C(=C)C(=O)O)C(=O)O |
|
Glycerophospho-N-palmitoyl ethanolamine |
C21H44N7P |
10.235 |
CCCCCCCCCCCCCCCC(=O)NCCOP(=O)(O)OCC(CO)O |
53393933 |
Garcinol |
C38 H50O6 |
14.17 |
C=C(C)C(CC=C(C)C)CC12CC(CC=C(C)C)C(C)(C)C(CC=C(C)C)(C(=O)C(=C(O)c3ccc(O)c(O)c3)C1=O)C2=O |
5490884 |
(−)-Desoxygambogenin |
C38 H48O6 |
13.907 |
CC(C)=CCCC(C)=CCc1c(O)c(CC=C(C)C)c2c(c1O)C(=O)C1=CC3CC4C(C)(C)OC(CC=C(C)C)(C3=O)C14O2 |
|
3,4-Dihydroxyphenylacetic acid |
C8H8O4 |
7.814 |
C1=CC(=C(C=C1CC(=O)O)O)O |
|
32-hydroxy-ent-guttiferone M |
C38H50O7 |
13.055 |
CC(C)=CCC(O)C(C)=CCC12CC(CC=C(C)C)C(C)(C)C(CC=C(C)C)(C(=O)C(=C(O)c3ccc(O)c(O)c3)C1=O)C2=O |
|
Nootkatone |
C15H22O |
10.785 |
C=C(C)C1CCC2=CC(=O)CC(C)C2(C)C1 |
|
Questiomycin C |
C13H10N2OS |
2.325 |
CS(=O)C1=C(C(=O)C=C2C1=NC3=CC=CC=C3O2)N |
|
(+)-ar-Turmerone |
C15H20O |
9.762 |
CC(C)=CC(=O)CC(C)c1ccc(C)cc1 |
|
Polygodial |
C15H22O2 |
10.455 |
CC1(C)CCCC2(C)C(C=O)C(C=O)=CCC12 |
|
Ginkgetin |
C32H22O10 |
10.122 |
COC1=C(C=C(C=C1)C2=CC(=O)C3=C(C=C(C=C3O2)OC)O)C4=C(C=C(C5=C4OC(=CC5=O)C6=CC=C(C=C6)O)O)O |
5271805 |
Thermolide G |
C34H63NO9 |
12.29 |
CC(=O)OC(C(C)CC(C)C(O)C(C)C(C)O)C(C)CC(C)C1OC(=O)C(C(C)C)NC(=O)CC(O)CC(O)C(C)CC1C |
|
Caerulomycin F |
C12H12NO2 |
2.825 |
COC1=CC(=NC(=C1)C2=CC=CC=N2)CO |
|
Momilactone |
C20H26O3 |
10.425 |
C=CC1(C)CCC2C(=CC3OC(=O)C4(C)C(=O)CCC2(C)C34)C1 |
|
18-β-Glycyrrhetinic acid |
C30 H46O4 |
10.922 |
CC1(C(=O)O)CCC2(C)CCC3(C)C(=CC(=O)C4C5(C)CCC(O)C(C)(C)C5CCC43C)C2C1 |
10114 |
C21H33NO5 |
10.307 |
C=C1CCC2(O)C(CC2(C)C)C(CO)=CC(O)C1NC(=O)C=C(C)CCO |
||
C16H11NO3 |
6.628 |
C1=CC=C(C=C1)C2=C(C(=O)NC2=O)C3=CC=C(C=C3)O |
||
Farinomalein |
C10 H13NO4 |
1.981 |
CC(C)C1=CC(=O)N(C1=O)CCC(=O)O |
|
C32H60O6 |
15.272 |
CCCCCCCCCC1(CO)CCC(C)C(=O)OC(CO)(CCCCCCCCC)CCC(C)C(=O)O1 |
||
C12H20N2O4 |
4.626 |
CC(C)CC1=C([N+](=C(C(=O)N1O)CC(C)C)[O-])O |
||
C12H11NO4 |
2.805 |
CC1=C(C(=O)C2=CN(C(=O)C=C2C1=O)C)OC |
||
C27H28 N2O4 |
9.972 |
CC(=O)OCC(Cc1ccccc1)NC(=O)C(Cc1ccccc1)NC(=O)c1ccccc1 |
||
C12H15NO3 |
1.938 |
COC1CC(C)Nc2c(C(=O)O)cccc21 |
||
C30H55NO10 |
11.726 |
CCC1OC(=O)C(C)C(O)C(C)C(OC2OC(C)CC(N(C)C)C2O)C(C)(OC)CC(C)C(=O)C(C)C(O)C1(C)O |
||
C15 H15 N O2 |
8.988 |
CC1=C(C(=C(C2=C1NC3=CC=CC=C32)O)OC)C |
Prediction of compound biological activities
PASS Online prediction analysis (Table 3) was employed to provide a preliminary estimation of the potential biological activities of the compounds identified from S. junghuhnianum. The results indicated that several compounds were associated with predicted activities related to antimicrobial, antiviral, antiprotozoal, and enzyme-inhibitory functions, including activities linked to processes such as cell-wall biosynthesis [22]. A number of metabolites exhibited moderate to high Pa values, suggesting potential biological relevance within these activity categories. Compounds such as gluconic acid, 3,4-dihydroxyphenylacetic acid, thermolide G, 18-β-glycyrrhetinic acid, and farinomalein displayed predicted activity across multiple categories, indicating chemical diversity in their possible biological roles.
Among the analyzed compounds, 5-O-desosaminyl-6-O-methylerythronolide A showed relatively high Pa values in several predicted activity classes, including general anti-infective, anti-Helicobacter pylori, and antiprotozoal categories. While these results suggest a broad predicted bioactivity profile, they should be interpreted cautiously, as PASS predictions do not confirm biological efficacy and require experimental validation.
Several other compounds, including ursolic acid, bilobetin, nootkatone, ginkgetin, and caerulomycin F, were predicted to possess antiprotozoal-related activities, particularly against Leishmania and Trypanosoma. These predictions may reflect possible interactions with protozoal cellular targets; however, specific mechanisms of action cannot be inferred solely from PASS analysis. In addition, a subset of compounds was predicted to be associated with enzyme-related activities, including glucan endo-1,3-β-D-glucosidase and peptidoglycan glycosyltransferase, which may be relevant to antimicrobial processes but remain speculative at this stage.
Predicted antiviral activities were also observed for several metabolites, particularly in categories associated with influenza viruses, rhinoviruses, and picornaviruses. Compounds such as 18-β-glycyrrhetinic acid, 6-gingerol, and brosimacutin C exhibited moderate to high Pa values in selected antiviral categories, suggesting potential relevance for further exploratory studies. In contrast, compounds including helquinoline, momilactone, and mimosamycin consistently showed low predicted activity scores and may therefore be considered lower-priority candidates for subsequent investigation.
Overall, the PASS prediction results indicate that a subset of the analyzed compounds may possess potential antimicrobial and anti-infective relevance, serving as a basis for hypothesis generation and compound prioritization. These computational predictions provide preliminary guidance for future experimental screening, but in vitro and in vivo validation will be essential to confirm the biological significance of the predicted activities.
Table 3 Prediction of the biological activity of compounds from Sphagnum junghuhnianum via the pass online test.
Compound name |
Biological activity |
Pa |
Pi |
Criteria |
4-(3-Dodecanyl) Benzenesulfonic Acid |
Antiseptic |
0.787 |
0.004 |
High |
|
Membrane integrity antagonist |
0.775 |
0.009 |
High |
|
Antiinfective |
0.727 |
0.006 |
High |
Gluconic Acid |
Macrophage stimulant |
0.914 |
0.001 |
Very high |
|
Membrane integrity agonist |
0.926 |
0.005 |
Very high |
|
Levansucrase inhibitor |
0.964 |
0.000 |
Very high |
4-Methoxycinnamaldehyde |
Membrane integrity agonist |
0.833 |
0.028 |
Very high |
|
Polyporopepsin inhibitor |
0.768 |
0.025 |
High |
|
Antiprotozoal (Leishmania) |
0.502 |
0.024 |
High |
Guttiferone E |
Antibacterial |
0.621 |
0.008 |
High |
|
Antiprotozoal (Trypanosoma) |
0.591 |
0.009 |
High |
|
Antiviral (Rhinovirus) |
0.488 |
0.029 |
Low |
Dilaurylmethylamine |
Antiviral (Poxvirus) |
0.630 |
0.013 |
High |
|
Antiviral (Picornavirus) |
0.547 |
0.033 |
High |
|
Antiviral (Adenovirus) |
0.515 |
0.00 |
High |
Ursolic Acid |
Antiprotozoal (Leishmania) |
0.915 |
0.003 |
Very high |
|
Antiviral (Influenza) |
0.761 |
0.004 |
High |
|
Glucanendo-1,3-beta-D-glucosidase inhibitor |
0.572 |
0.034 |
High |
6-Gingerol |
Antiviral (Rhinovirus) |
0.553 |
0.012 |
High |
|
Antiviral (Influenza) |
0.466 |
0.029 |
Low |
|
Antiprotozoal (Leishmania) |
0.401 |
0.048 |
Low |
Bilobetin |
Antiprotozoal (Leishmania)
|
0.535 |
0.020 |
High |
|
Antimycobacterial |
0.525 |
0.016 |
High |
|
Antiviral (Herpes) |
0.477 |
0.013 |
Low |
Xylarioic acid B |
Protein synthesis inhibitor |
0.797 |
0.002 |
High |
|
Protein 50S ribosomal subunit inhibitor |
0.711 |
0.001 |
High |
|
Peptidoglycan glycosyltransferase inhibitor |
0.523 |
0.018 |
High |
Brosimacutin C |
Antifungal |
0.523 |
0.027 |
High |
|
Antiviral (Rhinovirus) |
0.619 |
0.005 |
High |
|
Antiviral (Influenza) |
0.462 |
0.029 |
Low |
Ceriporic acid C |
Antiviral (Rhinovirus) |
0.606 |
0.006 |
High |
|
Peptidoglycan glycosyltransferase inhibitor |
0.470 |
0.029 |
Low |
|
Pediculicide |
0.455 |
0.012 |
Low |
Glycerophospho-N-palmitoyl ethanolamine |
Antiprotozoal (Leishmania) |
0.640 |
0.012 |
High |
|
Antiviral (Rhinovirus) |
0.585 |
0.008 |
High |
|
DNA polymerase I inhibitor |
0.475 |
0.010 |
Low |
Garcinol |
Antibacterial |
0.621 |
0.008 |
High |
|
Antiprotozoal (Trypanosoma) |
0.591 |
0.009 |
High |
|
Antiviral (Rhinovirus) |
0.488 |
0.029 |
Low |
(−)-Desoxygambogenin |
Antibacterial |
0.621 |
0.008 |
High |
|
Antiprotozoal (Trypanosoma) |
0.591 |
0.009 |
High |
|
Antiviral (Rhinovirus) |
0.488 |
0.029 |
Low |
3,4-Dihydroxyphenylacetic acid |
Membrane integrity agonist |
0.918 |
0.007 |
Very high |
|
Penicillin amidase inhibitor |
0.912 |
0.002 |
Very high |
|
Antiseborrheic |
0.868 |
0.007 |
Very high |
32-hydroxy-ent-guttiferone M |
Antibacterial |
0.630 |
0.007 |
High |
|
Antifungal |
0.509 |
0.029 |
High |
|
Antimycoplasmal |
0.179 |
0.068 |
Very low |
Nootkatone |
Antiprotozoal (Leishmania) |
0.523 |
0.022 |
High |
|
Antifungal |
0.346 |
0.064 |
Low |
|
Antibacterial |
0.316 |
0.054 |
Low |
Questiomycin C |
Antimycobacterial |
0.500 |
0.019 |
High |
|
Antituberculosic
|
0.284
|
0.080
|
Very low |
|
Antibacterial |
0.273 |
0.071 |
Very low |
(+)-ar-Turmerone |
Antifungal |
0.557 |
0.023 |
High |
|
Antiviral (Rhinovirus) |
0.498 |
0.025 |
Low |
|
Antiparasitic |
0.463 |
0.020 |
Low |
Polygodial |
Antifungal |
0.444 |
0.040 |
Low |
|
Peptidoglycan glycosyltransferase inhibitor |
0.458 |
0.032 |
Low |
|
Antiacne |
0.442 |
0.009 |
Low |
Ginkgetin |
Antiprotozoal (Leishmania) |
0.544 |
0.019 |
High |
|
Antimycobacterial
|
0.523 |
0.016 |
High |
|
Antifungal |
0.514 |
0.028 |
High |
Thermolide G |
Antifungal |
0.887 |
0.002 |
Very high |
|
Antibacterial |
0.782 |
0.003 |
High |
|
Antiparasitic |
0.682 |
0.006 |
High |
Caerulomycin F |
Antiviral (Picornavirus) |
0.532 |
0.038 |
High |
|
Antiprotozoal (Amoeba) |
0.525 |
0.008 |
High |
|
Anti-Helicobacter pylori |
0.482 |
0.005 |
Low |
Momilactone |
Antifungal |
0.467 |
0.036 |
Low |
|
Antiprotozoal (Leishmania) |
0.485 |
0.026 |
Low |
|
Antibacterial |
0.338 |
0.046 |
Low |
18-β-Glycyrrhetinic acid |
Antiviral (Influenza) |
0.892 |
0.002 |
Very high |
|
Antifungal |
0.570 |
0.022 |
High |
|
Antibacterial |
0.352 |
0.043 |
Low |
Punctaporonin K |
Antiviral (Rhinovirus) |
0.471 |
0.037 |
Low |
|
Antibacterial |
0.392 |
0.032 |
Low |
|
Antifungal |
0.319 |
0.074 |
Low |
Aqabamycin A |
Glucan endo-1,6-beta-glucosidase inhibitor |
0.776 |
0.010 |
High |
|
Antiviral (Picornavirus) |
0.631 |
0.013 |
High |
|
Antibacterial |
0.188 |
0.130 |
Low |
Farinomalein |
Muramoyltetrapeptide carboxypeptidase inhibitor |
0.889 |
0.004 |
Very high |
|
Glucan endo-1,3-beta-D-glucosidase inhibitor |
0.885 |
0.003 |
Very high |
|
Tpr proteinase (Porphyromonas gingivalis) inhibitor |
0.842 |
0.003 |
Very high |
Malyngolide dimer |
Beta-mannosidase inhibitor |
0.752 |
0.004 |
High |
|
Glucan endo-1,3-beta-D-glucosidase inhibitor |
0.593 |
0.029 |
High |
|
Antiviral (Picornavirus) |
0.490 |
0.055 |
Low |
Pulcherriminic acid |
RNA-directed RNA polymerase inhibitor |
0.588 |
0.003 |
High |
|
Glucan endo-1,6-beta-glucosidase inhibitor |
0.585 |
0.037 |
High |
|
Glucan endo-1,3-beta-D-glucosidase inhibitor |
0.453 |
0.077 |
Low |
Mimosamycin |
Antimycobacterial |
0.426 |
0.033 |
Low |
|
Antituberculosic |
0.413 |
0.028 |
Low |
|
Antibacterial |
0.372 |
0.037 |
Low |
Asperglaucide |
Antiviral (Picornavirus) |
0.434 |
0.085 |
Low |
|
Antiviral (Adenovirus) |
0.296 |
0.092 |
Very low |
|
Antiviral (Poxvirus) |
0.232 |
0.114 |
Very low |
Helquinoline |
Antituberculosic |
0.274 |
0.087 |
Very low |
|
Isopenicillin-N epimerase inhibitor |
0.275 |
0.089 |
Very low |
|
Glucan 1,3-beta-glucosidase inhibitor |
0.204 |
0.014 |
Very low |
5-O-Desosaminyl-6-O-methylerythronolide A |
Antiinfective |
0.980 |
0.002 |
Very high |
|
Anti-Helicobacter pylori |
0.957 |
0.001 |
Very high |
|
Antiprotozoal (Leishmania) |
0.953 |
0.002 |
Very high |
Carbazomycin B |
Anti-Helicobacter pylori |
0.254 |
0.058 |
Very low |
|
Antiprotozoal (Leishmania) |
0.387 |
0.054 |
Low |
|
Antiprotozoal (Trypanosoma) |
0.258 |
0.139 |
Very low |
Prediction of drug-likeness
The analysis revealed varying degrees of compliance with drug-likeness criteria among the compounds derived from S. junghuhnianum (Table 4). According to Lipinski’s Rule of 5, compounds with a molecular weight (MW) ≤ 500 g/mol, logP ≤ 5, no more than 10 hydrogen bond acceptors (N and O atoms), and no more than 5 hydrogen bond donors (NH and OH groups), and with no more than one rule violation tend to exhibit good permeability and absorption in the gastrointestinal tract [23]. Most compounds, such as 4-methoxycinnamaldehyde, 6-gingerol, xylarioic acid B, nootkatone, polygodial, and momilactone, showed no violations, indicating balanced lipophilicity and hydrogen-bonding capacity that support membrane permeability. Recent studies further support these observations, showing that compounds with excessively high logP or MlogP values often face limitations in drug development and require optimization to improve their pharmacokinetic behavior and overall drug-likeness [24,25]. These findings highlight the importance of fine-tuning molecular properties to enhance both efficacy and safety profiles of lead candidates.
Several other compounds, including guttiferone E, ursolic acid, bilobetin, and ginkgetin, exhibited a single Lipinski rule violation, typically related to MW or logP. A single violation is generally considered acceptable, as many bioactive natural products share similar physicochemical characteristics and nevertheless display potent biological activity. In contemporary drug discovery, such molecules remain suitable for further optimization through structural modification or formulation strategies. Conversely, compounds such as malyngolide dimer and 5-O-desosaminyl-6-O-methylerythronolide A showed 2 violations, mainly due to their high molecular weight and excessive numbers of hydrogen bond donors or acceptors. Compounds with more than one violation are more likely to exhibit low membrane permeability and limited oral bioavailability, and therefore may require advanced strategies such as nanoformulations, prodrug approaches, or targeted delivery systems to improve their pharmacokinetic profiles.
Overall, these results indicate that most metabolites listed in Table 4 possess physicochemical properties that are compatible with preliminary drug-likeness assessment. Compounds with no Lipinski violations may be prioritized for further exploratory studies, while those with one or more violations remain relevant within a natural-product-oriented discovery framework, particularly considering that many bioactive natural compounds fall within the “beyond the rule of five” (bRO5) chemical space.
Table 4 Physicochemical properties of compounds from Sphagnum junghuhnianum based on Lipinskiʼs Rule of 5.
Compound name |
Lipinski |
||||
MW |
MlogP ≤ 4.15 |
NorO ≤ 10 |
NHorOH ≤ 5 |
Violation |
|
326.49 |
4.49 |
3 |
1 |
||
Gluconic Acid |
196.16 |
‒2.90 |
7 |
6 |
Yes (1) |
4-Methoxycinnamaldehyde |
162.19 |
1.66 |
2 |
0 |
Yes (0) |
Guttiferone E |
602.80 |
3.78 |
10 |
6 |
Yes (1) |
Ursolic Acid |
456.70 |
5.82 |
3 |
2 |
Yes (1) |
6-Gingerol |
294.39 |
2.14 |
4 |
2 |
Yes (0) |
Bilobetin |
552.48 |
0.44 |
10 |
5 |
Yes (1) |
Xylarioic acid B |
234.29 |
0.31 |
5 |
4 |
Yes (0) |
Brosimacutin C |
342.39 |
1.63 |
5 |
3 |
Yes (0) |
(−)-Desoxygambogenin |
600.78 |
4.12 |
6 |
2 |
Yes (1) |
3,4-Dihydroxyphenylacetic acid |
168.15 |
0.47 |
4 |
3 |
Yes (0) |
32-hydroxy-ent-guttiferone M |
618.80 |
2.98 |
7 |
4 |
Yes (1) |
Nootkatone |
218.33 |
3.46 |
1 |
0 |
Yes (0) |
Questiomycin C |
274.30 |
‒0.00 |
4 |
1 |
Yes (0) |
(+)-ar-Turmerone |
216.32 |
3.68 |
1 |
0 |
Yes (0) |
Polygodial |
234.33 |
2.54 |
2 |
0 |
Yes (0) |
Ginkgetin |
566.51 |
0.63 |
10 |
4 |
Yes (1) |
Thermolide G |
629.87 |
2.06 |
9 |
5 |
Yes (1) |
Caerulomycin F |
216.24 |
0.02 |
4 |
1 |
Yes (0) |
Momilactone |
314.42 |
3.57 |
3 |
0 |
Yes (0) |
18-β-Glycyrrhetinic acid |
470.68 |
4.87 |
4 |
2 |
Yes (1) |
Aqabamycin A |
265.26 |
2.05 |
3 |
2 |
Yes (0) |
Farinomalein |
211.21 |
0.55 |
4 |
1 |
Yes (0) |
Malyngolide dimer |
540.82 |
4.52 |
6 |
2 |
No (2) |
Mimosamycin |
233.22 |
‒0.34 |
4 |
0 |
Yes (0) |
Asperglaucide |
444.52 |
3.41 |
4 |
2 |
Yes (0) |
5-O-Desosaminyl-6-O-methylerythronolide A |
589.76 |
‒0.16 |
11 |
4 |
No (2) |
Carbazomycin B |
241.29 |
2.35 |
2 |
2 |
Yes (0) |
Notes: MW: Molecular Weight (measured in g/mol); MlogP: Calculated octanol/water partition coefficient (lipophilicity); NorO: Total oxygen atoms; NhorOH: Total nitrogen + hydroxyl groups.
Molecular interactions of Sphagnum junghuhnianum bioactive compounds with PBP1 (7O4B)
Molecular docking analysis was conducted using Penicillin-Binding Protein 1 (PBP1; PDB ID: 7O4B), a transpeptidase involved in peptidoglycan biosynthesis in Staphylococcus aureus. The docking results (Table 5) demonstrated variation in predicted binding energies, ranging from −5.7 to −9.8 kcal/mol, suggesting differential ligand accommodation within the PBP1 active-site region. Among the analyzed compounds, ursolic acid showed the most favorable binding affinity (−9.8 kcal/mol), exceeding that of the reference inhibitor (7OPB; −7.3 kcal/mol).
Ursolic acid was predicted to interact predominantly through hydrophobic contacts with residues ILE348, LEU416, PHE423, TYR527, and TYR534, along with a π-sigma interaction involving TYR534. These interactions suggest that the triterpenoid scaffold of ursolic acid may fit favorably within a hydrophobic region of the PBP1 binding pocket. Other compounds with comparatively low binding energy values included bilobetin and thermolide G (both −9.4 kcal/mol), which formed multiple predicted hydrogen bonds with residues such as SER349, ARG353, SER368, THR514, THR516, and TYR534 (Figures 4 and 5). The presence of hydroxyl and phenolic functional groups in these molecules may contribute to their predicted interaction patterns.
Additional metabolites, including 18-β-glycyrrhetinic acid, brosimacutin C, nootkatone, and (−)-desoxygambogenin, were also predicted to interact with residues frequently associated with the PBP1 binding site, such as SER314, SER368, ASN370, THR514/516, TRP351, PHE423, TYR534, and TYR566. The recurrence of these interactions across multiple ligands suggests a degree of structural compatibility with the PBP1 active-site environment.
Overall, the docking results indicate that hydrogen bonding involving polar residues (e.g., SER, THR, ASN) and hydrophobic or aromatic interactions involving residues such as TRP, PHE, and TYR may contribute to ligand stabilization within the PBP1 binding region. However, these interaction patterns should be interpreted as computational predictions that provide preliminary, hypothesis-generating insights, rather than direct evidence of enzymatic inhibition or disruption of peptidoglycan biosynthesis. Experimental validation through quantitative enzymatic or cellular assays will be necessary to clarify the functional relevance of these predicted interactions.
Table 5 Binding residues and binding energies of bioactive compounds from Sphagnum Junghuhnianum docked with 7O4B (Pbp1).
Ligand |
Interaction with |
Binding Affinity (kcal/mol) |
|||
Conventional H-Bond |
Carbon H-Bond |
Pi-Alkyl |
Pi-Sigma |
||
Control |
SER A:314 SER A:368 ASN A:370 GLN A:425 THR A:516 |
|
TYR A:566
|
TRP A:351 |
‒7.3 |
Bioactive components of the S. junghuhnianum |
|||||
4-(3-Dodecanyl) Benzenesulfonic Acid |
SER A:314 ASN A:370 |
|
HIS A:499 |
TRP A:351 |
‒6.9 |
Gluconic Acid |
SER A:314 SER A:368 ASN A:370 THR A:516 |
TRP A:351 |
|
|
‒5.7 |
4-Methoxycinnamaldehyde |
SER A:314 SER A:368 LYS A:317 |
|
|
|
‒5.7 |
Guttiferone E |
SER A:314 ASN A:370 |
|
ILE A:348 PHE A:423 PRO A:533 |
TYR A:566 |
‒8.2 |
Ursolic Acid |
|
|
ILE A:348 LEU A:416 PHE A:423 TYR A:527 |
TYR A:534 |
‒9.8 |
6-Gingerol |
SER A:314 SER A:368 ASN A:370 TYR A:534 |
THR A:514 |
PHE A:423 |
|
‒6.1 |
Bilobetin |
SER A:349 ARG A:353 SER A:368 THR A:514 |
THR A:516 TYR A:534 |
|
|
‒9.4 |
Xylarioic acid B |
TYR A:503 THR A:516 GLY A:515 SER A:537 |
GLY A:569 |
TYR A:566 |
TRP A:351 |
‒6.5 |
Brosimacutin C |
SER A:314 LYS A:317 SER A:368 TYR A:534 |
|
|
|
‒8.2 |
(−)-Desoxygambogenin |
SER A:314 ASN A:370 TYR A:534 |
|
LYS A:529 PRO A:533 TYR A:566 |
|
‒7.8 |
3,4-Dihydroxyphenylacetic acid |
ARG A:353 ASN A:370 TYR A:534 |
|
TYR A:566 |
|
‒7.9 |
32-hydroxy-ent-guttiferone M |
SER A:314 ASN A:370 THR A:514 THR A:516 |
|
|
|
‒6.1 |
Nootkatone |
SER A:349 |
|
ILE A:348 TRP A:351 PHE A:423 TYR A:527 TYR A:566 |
|
‒8.1 |
Questiomycin C |
THR A:514 |
|
|
TRP A:351 TYR A:566 |
‒6.9 |
(+)-ar-Turmerone |
SER A:314 THR A:516 TYR A:566 |
|
|
|
‒7.9 |
Polygodial |
SER A:314 LYS A:317 SER A:368 |
|
HIS A:499 |
TRP A:351 |
‒6.8 |
Ginkgetin |
SER A:314 LYS A:317 SER A:368 |
|
TYR A:534 TYR A:566 |
|
‒6.9 |
Thermolide G |
SER A:349 ARG A:353 SER A:368 THR A:514 |
THR A:516 TYR A:534 TYR A:566
|
|
|
‒9.4 |
Caerulomycin F |
SER A:314 LYS A:317 SER A:349 SER A:368 THR A:514 |
ALA A:500 |
TRP A:351 PHE A:423 TYR A:534 TYR A:566 |
|
‒8.1 |
Momilactone |
TRP A:351 THR A:516 |
|
|
|
‒6.9 |
18-β-Glycyrrhetinic acid |
THR A:514 THR A:516 |
|
TRP A:351 ALA A:500 TYR A:566 |
|
‒8.7 |
Aqabamycin A |
SER A:314 SER A:368 THR A:514 THR A:516 TYR A:566 |
|
TRP A:351 |
|
‒7.6 |
Farinomalein |
THR A:514 |
|
|
|
‒7.8 |
Mimosamycin |
SER A:314 SER A:368 TYR A:566 |
|
PHE A:423 |
TRP A:351 |
‒6.5 |
Helquinoline |
SER A:314 SER A:368 LYS A:317 ASN A:370 TYR A:566 |
PHE A:423 THR A:516 |
|
TRP A:351 |
‒7.4 |
Carbazomycin B |
TYR A:503 THR A:514 GLY A:515 THR A:516 SER A:537 |
|
ALA A:500 |
|
‒7.4 |
The in-silico analyses were evaluated in relation to the in vitro antimicrobial screening results presented in Table 1. The methanolic extract of S. junghuhnianum exhibited concentration-dependent antibacterial effects in disk diffusion assays, with the largest inhibition zones observed at 100% extract concentration, particularly against S. pyogenes (42.01 ± 0.33 mm) and S. aureus (34.62 ± 2.21 mm). These Gram-positive pathogens rely on penicillin-binding proteins (PBPs) for cell wall biosynthesis, which may partly explain their higher susceptibility under the tested conditions. Moderate inhibition was also observed against S. typhi (22.36 ± 0.84 mm) and P. gingivalis (18.05 ± 0.41 mm), whereas no inhibition was detected against K. pneumoniae or A. niger, indicating a selective antibacterial spectrum of the extract.
Consistent with the antibacterial screening results, the extract did not exhibit antifungal activity against C. albicans, as no inhibition zone was observed, while the positive control produced clear antifungal effects. Molecular docking analysis suggested that several moss-derived metabolites may interact favorably with the PBP1 active site, exhibiting binding poses and affinity values comparable to or lower than those of the reference ligand. These predicted interactions provide preliminary insights into a possible molecular basis for the preferential antibacterial activity observed against Gram-positive bacteria. However, such interactions should be regarded as hypothesis-generating, as molecular docking alone does not confirm enzymatic inhibition or downstream effects on peptidoglycan biosynthesis.
Overall, this study indicates that S. junghuhnianum is a chemically rich source of secondary metabolites with potential antibacterial relevance. LC-HRMS profiling revealed 514 putatively identified metabolites, of which 35 compounds were selected for further computational evaluation based on database annotation and PASS predictions. By integrating in vitro antibacterial screening with metabolomic profiling and in silico analyses, this work provides preliminary, multi-level evidence supporting the antibacterial potential of S. junghuhnianum metabolites. Further studies involving quantitative antimicrobial assays, compound isolation, and target-based validation are required before their potential application in antimicrobial drug development can be fully assessed.
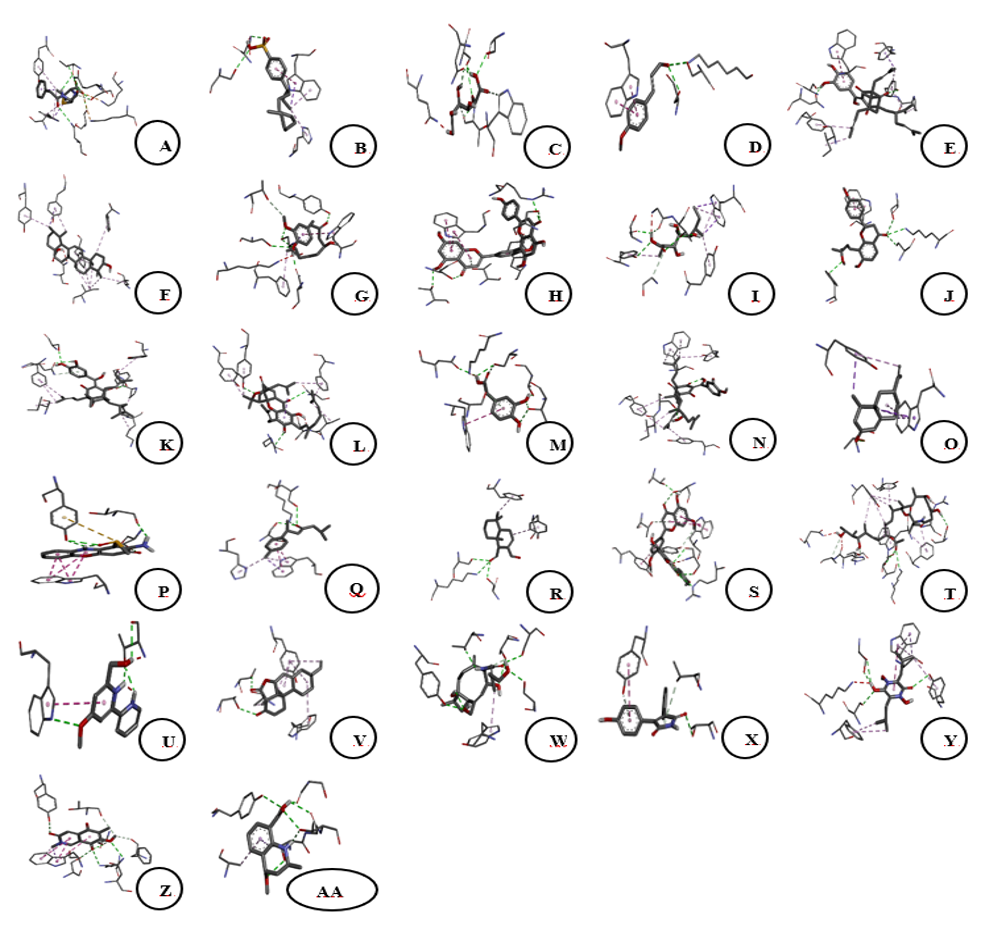
Figure 4 Three-dimensional binding interactions of 7OPB (control) and bioactive compounds from Sphagnum junghuhnianum with the 7O4B protein (PBP1). A. 7OPB; B. 4-(3-Dodecanyl) Benzenesulfonic Acid; C. Gluconic Acid; D. 4-Methoxycinnamaldehyde; E. Guttiferone E; F. Ursolic Acid; G. 6-Gingerol; H. Bilobetin; I. Xylarioic acid B; J. Brosimacutin C; K. (−)-Desoxygambogenin; L. 3,4-Dihydroxyphenylacetic acid; M. 32-hydroxy-ent-guttiferone N; L. Nootkatone; O. Questiomycin P; N. (+)-ar-Turmerone; Q. Polygodial; R. Ginkgetin; S. Thermolide G; T. Caerulomycin U; S. Momilactone; V. 18-β-Glycyrrhetinic acid; W. Aqabamycin A; X. Farinomalein; Y. Mimosamycin; Z. Helquinoline; AA. Carbazomycin B.
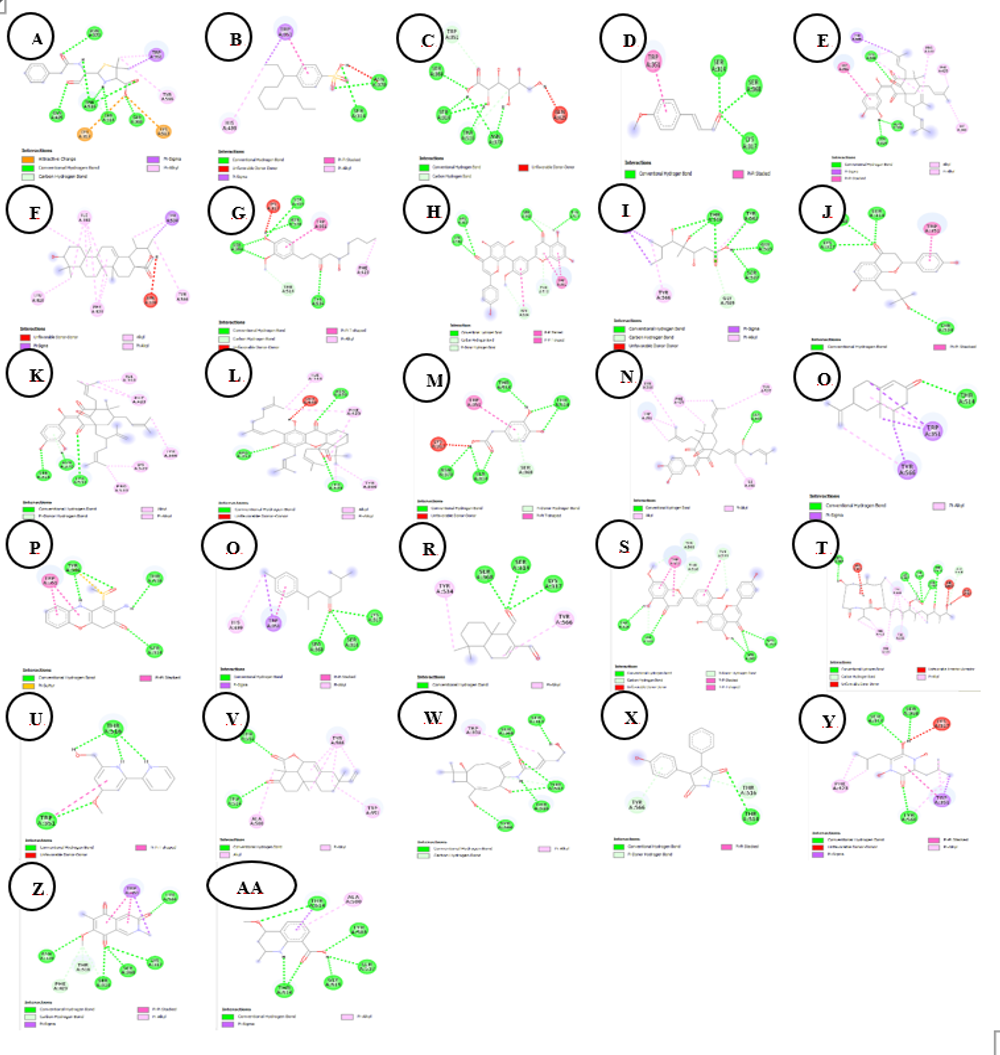
Figure 5 Two-dimensional binding interactions of 7OPB (control) and bioactive compounds from Sphagnum junghuhnianum against the 7O4B protein. A. 7OPB; B. 4-(3-Dodecanyl) Benzenesulfonic Acid; C. Gluconic Acid; D. 4-Methoxycinnamaldehyde; E. Guttiferone E; F. Ursolic Acid; G. 6-Gingerol; H. Bilobetin; I. Xylarioic acid B; J. Brosimacutin C; K. (−)-Desoxygambogenin; L. 3,4-Dihydroxyphenylacetic acid; M. 32-hydroxy-ent-guttiferone N; L. Nootkatone; O. Questiomycin P; N. (+)-ar-Turmerone; Q. Polygodial; R. Ginkgetin; S. Thermolide G; T. Caerulomycin U; S. Momilactone; V. 18-β-Glycyrrhetinic acid; W. Aqabamycin A; X. Farinomalein; Y. Mimosamycin; Z. Helquinoline; AA. Carbazomycin B.
Conclusions
The methanolic extract of Sphagnum junghuhnianum demonstrated notable antibacterial activity in preliminary disk diffusion assays, particularly against Gram-positive bacteria, while no antifungal activity was observed under the tested conditions. LC-HRMS profiling revealed a chemically diverse metabolite composition, with several compounds putatively identified as having potential antimicrobial relevance. In silico molecular docking analysis suggested favorable interactions of ursolic acid, bilobetin, and thermolide G with Penicillin-Binding Protein 1 (PBP1), providing preliminary insights into a possible antibacterial mode of action. Overall, these findings indicate that S. junghuhnianum represents a promising source of antibacterial candidates, warranting further studies involving quantitative antimicrobial assays, compound isolation, and experimental validation to support its potential for future drug development.
Acknowledgements
This research was supported by the Ministry of Education, Culture, Research and Technology of the Republic of Indonesia, through the BIMA Research Grant (Contract Number: 112/C3/DT.05.00/PL/2025).
Declaration of Generative AI in Scientific Writing
The authors acknowledge the use of generative AI tools (such as QuillBot and ChatGPT by OpenAI) in thepreparation of this manuscript, specifically for language editing and grammar correction. No content generation or data interpretation was performed by AI. The authors take full responsibility for the content and conclusions of this work.
CRediT author statement
Sinta R Pardosi: Investigation, methodology, data collection, data analysis, original draft writing. Etti Sartina Siregar: Conceptualization, investigation, review, editing, corresponding author. Isnaini Nurwahyuni: Research design, project administration, data curation, review, editing.
References
[1] GV Naumova, AE Tomson, NA Zhmakova, NL Makarova and TF Ovchinnikova. Biologically active compounds of different sphagnum peat species. Solid Fuel Chemistry 2015; 49, 135-140.
[2] F Chen, A Ludwiczuk, G Wei, X Chen, B Crandall-Stotler and JL Bowman. Terpenoid secondary metabolites in bryophytes: Chemical diversity, biosynthesis and biological functions. Critical Reviews in Plant Sciences 2018; 37(2-3), 210-231.
[3] G Montenegro, MC Portaluppi, FA Salas and MF Díaz. Biological properties of the chilean native moss Sphagnum magellanicum. Biological Research 2009; 42(2), 233-237.
[4] D Stenitzer, R Mócsai, H Zechmeister, R Reski, EL Decker and F Altmann. O-methylated N-glycans distinguish mosses from vascular plants. Biomolecules 2022; 12(1), 136.
[5] M Hymas, I Casademont-Reig, S Poigny and VG Stavros. Characteristic photoprotective molecules from the sphagnum world: A solution-phase ultrafast study of sphagnic acid. Molecules 2023; 28(16), 6153.
[6] JD Fudyma, J Lyon, R AminiTabrizi, H Gieschen, RK Chu, DW Hoyt, JE Kyle, J Toyoda, N Tolic, HM Heyman, NJ Hess, TO Metz and MM Tfaily. Untargeted metabolomic profiling of Sphagnum fallax reveals novel antimicrobial metabolites. Plant Direct 2019; 3(10), e00179.
[7] L Bryan, R Shaw, E Schoonover, A Koehl, S DeVries-Zimmerman and M Philben. Sphagnan in Sphagnum-dominated peatlands: Bioavailability and effects on organic matter stabilization. Biogeochemistry 2024; 167, 665-680.
[8] RS Shete, HV Wangikar, JJ Chavan, MB Kanade and SJ Chavan. Bioactive compounds of bryophytes: Unveiling antimicrobial properties and therapeutic potential. International Journal of Plant and Environment 2024, 10(3), 1-15.
[9] TA Çelik, ÖS Aslantürk, G Aslan and M Kirmaci. Determination of phytochemical content and antioxidant activities of Sphagnum divinum Flatberg & K. Hassel and Sphagnum girgensohnii Russow (Sphagnopsida). Anatolian Bryology 2022; 9(2), 58-69.
[10] M Singh, AKS Rawat and R Govindarajan. Antimicrobial activity of some Indian mosses. Fitoterapia 2007; 78(2), 156-158.
[11] DR Chhetri, S Yonzone and R Mukhia. Bioprospecting for enzymes in bryophytes: Extraction of L-Myo-inositol-1-phosphate synthase from Sphagnum junghuhnianum Doz. et Molk. and its characterization. South African Journal of Botany 2023; 136, 692-702.
[12] K Wacnik, VA Rao, X Chen, L Lafage, M Pazos, S Booth, W Vollmer, JK Hobbs, RJ Lewis and SJ Foster. Penicillin-binding protein 1 (PBP1) of Staphylococcus aureus has multiple essential functions in cell division. mBio 2022; 13(4), 0066922.
[13] DPM Sethuvel, YD Bakthavatchalam, MKM Irulappan, R Sherivastava, H Periasamy and V Veeraghavan. β-Lactam resistance in ESKAPE pathogens mediated through modifications in penicillin-binding proteins: An overview. Infectious Diseases and Therapy 2023; 12, 829-841.
[14] S Martinez-Caballero, KV Mahasenan, C Kim, R Molina, R Feltzer, M Lee, R Bouley, D Hesek, JF Fisher, IG Muñoz, M Chang, S Mobashery and JA Hermoso. Integrative structural biology of the penicillin-binding protein-1 from Staphylococcus aureus, an essential component of the divisome machinery. Computational and Structural Biotechnology Journal 2021; 19, 5392-5405.
[15] L Wu, Y Zhang, J Yang, H Liu and S Wang. Diversity and correlation analysis of microbiomes and metabolites of Sphagnum palustre in various microhabitats. BMC Plant Biology 2025; 25, 761.
[16] J Hudzicki. Kirby-Bauer disk diffusion susceptibility test protocol. ASM 2019; 15(1), 1-23.
[17] LR Valeeva, AL Dague, MH Hall, AE Tikhonova, MR Sharipova, MA Valentovic, LM Bogomolnaya and EV Shakirov. Antimicrobial activities of secondary metabolites from moss: A review. Antibiotics 2022; 11, 1004.
[18] L Klavina, G Springe, V Nikolajeva, I Martsinkevich, I Nakurte, D Dzabijeva and I Steinberga Chemical composition analysis, antimicrobial activity and cytotoxicity screening of moss extracts (Moss phytochemistry). Molecules 2015; 20(9), 17221-17243.
[19] S Xu, Y Li, H Wang, J Chen, L Liu and F Zhao. Plant flavonoids with antimicrobial activity against methicillin‑resistant Staphylococcus aureus (MRSA). ACS Infectious Diseases 2024; 10(8), 1594-1612.
[20] L Krunova, L Valeeva, A Ivanova, D Petrova, S Smirnov and E Fedorova. Antimicrobial activities of secondary metabolites from model mosses. Antibiotics 2022; 11(8), 1004.
[21] ML Astolfi, A Bianchi, F Rossi, G Conti, L Marino, P Santini and R De Luca. Sphagnum moss and peat comparative study: Metal release, binding properties and antioxidant activity. PLoS One 2024; 19(8), e0307210.
[22] A Lagunin, A Stepanchikova, D Filimonov and V Poroikov. PASS: prediction of activity spectra for biologically active substances. Bioinformatics 2000; 16(8), 747-748.
[23] CA Lipinski, F Lombardo, BW Dominy and PJ Feeney. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Advanced Drug Delivery Reviews 1997; 23(1‑3), 3-25.
[24] V Ivanović, M Rančić, B Arsić and A Pavlović. Lipinski's rule of five, famous extensions and famous exceptions. Chemia Naissensis 2020; 3(1), 171-177.
[25] TK Karami, S Hailu, S Feng, R Graham and HJ Gukasyan. Eyes on Lipinski's rule of five: A new “rule of thumb” for physicochemical design space of ophthalmic drugs. Journal of Ocular Pharmacology and Therapeutics 2022; 38(1), 43-55.