Trends
Sci.
2026;
23(9):
13423
Empagliflozin Attenuates Cardiac Fibrosis via Suppression of TLR2/NF-κB Signaling in a Rat Model of Metabolic Syndrome
Muhammad
Gibran Fauzi Harmani Kalim1,2,*,
![]() ,
Mohammad Saifur Rohman3,4,
,
Mohammad Saifur Rohman3,4,
![]() ,
,
Djanggan
Sargowo3,
![]() ,
Sri Winarsih5,
,
Sri Winarsih5,
![]() ,
Dian Nugrahenny6,
,
Dian Nugrahenny6,
![]() ,
Hidayat Sujuti7,
,
Hidayat Sujuti7,
![]() ,
,
Siska
Nanda Widhaningrum8,
![]() and Janjte Wiliem Souhaly9,
and Janjte Wiliem Souhaly9,
![]()
1Doctoral Program of Medical Science, Faculty of Medicine, Universitas Brawijaya, East Java 65145, Indonesia
2Department of Internal Medicine, Faculty of Medicine, YARSI University, Jakarta 10510, Indonesia
3Department of Cardiology and Cardiovascular Medicine, Faculty of Medicine, Universitas Brawijaya,
Saiful Anwar General Hospital, East Java 65145, Indonesia
4Cardiovascular Research Center, Universitas Brawijaya, East Java 65145, Indonesia
5Department of Pharmacy, Faculty of Medicine, Universitas Brawijaya, East Java 65145, Indonesia
6Department of Pharmacology, Faculty of Medicine, Universitas Brawijaya, East Java, 65145, Indonesia
7Department of Ophthalmology, Faculty of Medicine, Universitas Brawijaya, East Java, 65145, Indonesia
8Department of Medical Laboratory Technology, Sekolah Tinggi Ilmu Kesehatan Maharani,
East Java 65143, Indonesia
9Department of Biology, Faculty of Biology, Gadjah Mada University, Yogyakarta 55281, Indonesia
(*Corresponding author’s e-mail: [email protected])
Received: 23 January 2026, Revised: 25 February 2026, Accepted: 11 March 2026, Published: 10 April 2026
Abstract
The global prevalence of metabolic syndrome (MetS) is rising at an alarming rate. A critical complication of MetS is cardiac fibrosis eventually leading to cardiac dysfunction and death. Therefore, mitigating cardiac fibrosis is essential. Although empagliflozin (EMP) has demonstrated potent cardioprotective and antihyperglycemic properties, it is not yet the primary treatment for MetS. This highlights a critical need to explore EMP’s molecular benefits in MetS populations. This study investigated whether empagliflozin (EMP) attenuates cardiac fibrosis through suppression of TLR2/NF-κB signaling in a rat model of metabolic syndrome (MetS). Sprague–Dawley rats were assigned to a normal diet group and to a high-fat, high-sucrose (HFHS) diet followed by streptozotocin injection to induce MetS. MetS animals were further divided into untreated MetS, MetS treated with EMP 1 mg/kgBW, and MetS treated with EMP 30 mg/kgBW for nine weeks. Cardiac histopathology was evaluated using Hematoxylin–Eosin and Masson’s Trichrome staining, with fibrosis quantified as collagen volume fraction (CVF). mRNA expression of TLR2 and NF-κB was assessed by RT-PCR. Immunofluorescence was performed to evaluate α-SMA expression and nuclear translocation of the NF-κB p65 subunit, a functional indicator of pathway activation. EMP treatment improved metabolic parameters, including fasting blood glucose, triglyceride, and high-density lipoprotein levels. MetS significantly increased TLR2 and NF-κB expression, enhanced nuclear localization of NF-κB p65, elevated α-SMA expression, and promoted interstitial collagen deposition. EMP treatment attenuated these changes, as demonstrated by reduced CVF, downregulation of TLR2 and NF-κB mRNA expression (P < 0.05), diminished p65 nuclear translocation, and decreased α-SMA expression (P < 0.05). Casp1 expression was not significantly altered. In conclusion, empagliflozin attenuates cardiac fibrotic remodeling in a rat model of metabolic syndrome via suppression of TLR2/NF-κB signaling, as supported by reduced p65 nuclear translocation and decreased myofibroblast activation.
Keywords: Empagliflozin, Cardiac fibrosis, Metabolic syndrome, Inflammation, TLR2, NFKB, CASP1
Introduction
Metabolic syndrome (MetS) is a multifaceted disorder characterized by a cluster of metabolic abnormalities, including central obesity, insulin resistance, hypertension, and dyslipidemia. These conditions significantly increase the risk of cardiovascular disease and type 2 diabetes mellitus complications, leading to higher mortality rates [1,2]. Globally, the prevalence of MetS is rising, affecting an estimated 25% of the population. This trend is particularly evident in low- and middle-income nations, driven by rapid industrialization, sedentary lifestyles, and high-calorie diets [3]. In Indonesia, the national prevalence of MetS has reached 21.66%, with some provinces reporting rates as high as 50% [4]. The primary drivers in the Indonesian population are poorly managed low-density lipoprotein (LDL) cholesterol, hypertension, and central obesity [5]. Collectively, these metabolic disturbances trigger pathological remodeling, notably cardiac fibrosis, which serves as a precursor to heart failure and other adverse cardiovascular events.
Cardiac fibrosis, defined by the excessive deposition of extracellular matrix (ECM) by cardiac fibroblasts, is a prominent clinical feature of many heart disorders [6]. The differentiation of fibroblasts into myofibroblasts is characterized by elevated expression of α-smooth muscle actin (α-SMA), which develops a pro-fibrotic phenotype that is induced by transforming growth factor-β (TGF-β) signaling and the innate immunological response [7,8]. The initial response of cardiac tissue to oxidative damage involves activation of nuclear factor kappa-B (NFKB), which triggers toll-like receptors (TLRs). TLR2 plays a crucial role in innate immunity and contributes to cardiac inflammation and fibrosis by activating NFKB signaling pathways [9,10]. These activations lead to the upregulation of pro-fibrotic factors and the production of proinflammatory cytokines that induce apoptosis and cell death by activating various caspases, including caspase-1 (CASP1) [11]. Cleavage from pro-CASP1 into mature CASP1 might promote the maturation and secretion of proinflammatory cytokines, such as interleukin (IL)-1β and IL-18, which lead to sustained inflammation and fibrosis in the heart [12,13].
Recent studies have identified sodium-glucose co-transporter 2 inhibitors (SGLT2Is), such as empagliflozin (EMP), as a novel class of glucose-lowering agents that effectively modulate cardiac fibrosis. Beyond their primary role in inhibiting renal tubular glucose reabsorption to promote glucosuria, SGLT2Is offer significant cardiovascular benefits. Recent research into the anti-fibrotic properties of EMP has primarily focused on its modulation of the TGF-β/SMAD3 signaling pathway [14,15]. However, despite the well-documented cardiovascular benefits of EMP, the specific mechanisms underlying its effects in Metabolic Syndrome (MetS) models remain incomplete. In particular, the potential involvement of the TLR2/NFκB signaling axis and CASP1 in EMP-mediated reduction of cardiac fibrosis has yet to be fully elucidated. While previous studies have utilized empagliflozin (EMP) doses ranging from 10 mg/kg/day to 30mg/kg/day in animal models, these concentrations far exceed clinically approved human dosage. According to ESC and ACC/AHA guidelines, the standard dose for heart failure is 10 mg/day, which translates to an animal equivalent dose (AED) of approximately 1 mg/kg/day in Sprague-Dawley rats. High-dose animal trials may yield results that are not achievable in clinical practice; therefore, this study utilized a translational dose of 1 mg/kg/day. Our objective was to evaluate the efficacy of this clinically relevant dose in reducing cardiac fibrosis and to investigate its potential association with the TLR2/NFκB signaling pathway.
Materials and methods
Experimental Animals
Male Sprague-Dawley rats (8 - 12 weeks old, weighing 250 - 300 g) were obtained from the National Agency of Drug and Food Control (BPOM), Jakarta, Indonesia. The animals were acclimated for seven days under controlled environmental conditions (25±2 °C; 50±5% humidity) with light/dark cycle and free access to standard laboratory chow and water ad libitum. Following the acclimation period, the rats were randomly assigned to two primary experimental groups: A normal healthy diet control group (N, n = 7) and a high-fat high-sucrose diet group (HFHS, n = 28). The HFHS diet was prepared according to the composition described by Rohman et al. [16]. The HFHS were given for 10 - 11 weeks. The HFHS groups then received a single intraperitoneal injection of streptozotocin (STZ, 30 mg/kg) (bioWORLD, GeneLinx International Inc., USA). Successful induction of Metabolic Syndrome (MetS) was confirmed based on the NCEP-ATP III criteria, requiring the presence of at least three primary pathological indicators such as fasting glucose (≥ 125 mg/dL), triglycerides (≥ 150 mg/dL), and high-density lipoprotein (HDL) (≤ 40 mg/dL), which remained steady for a minimum of 4 - 6 weeks [16].
Following the induction of Metabolic Syndrome (MetS), rats were assigned to four experimental groups (6 rats per group) using stratified randomization method based on the baseline data including body weight, body length, abdominal cirfumference, fasting blood glucose, triglyceride, high-density lipoprotein to avoid bias data. The final experimental groups were divided into: (i) normal healthy control (N); (ii) untreated MetS; (iii) MetS treated with 1 mg/kg/day empagliflozin (EMP1); and (iv) MetS treated with 30 mg/kg/day empagliflozin (EMP2) with the number of animals was 7 per group (Figure 1). The number of sample size followed Federer’s formula with equation:
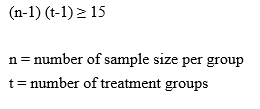
Empagliflozin was administered daily via oral gavage for nine weeks. All animals were treated and monitored with the same procedure as blind in vivo research. All animal procedures were conducted in accordance with the ARRIVE guidelines and EU Directive 2010/63/EU. The study protocol was reviewed and approved by the Health Research Ethics Commission of Saiful Anwar General Hospital, Malang, Indonesia (Ref. No: 400/211/K.3/302/2021).
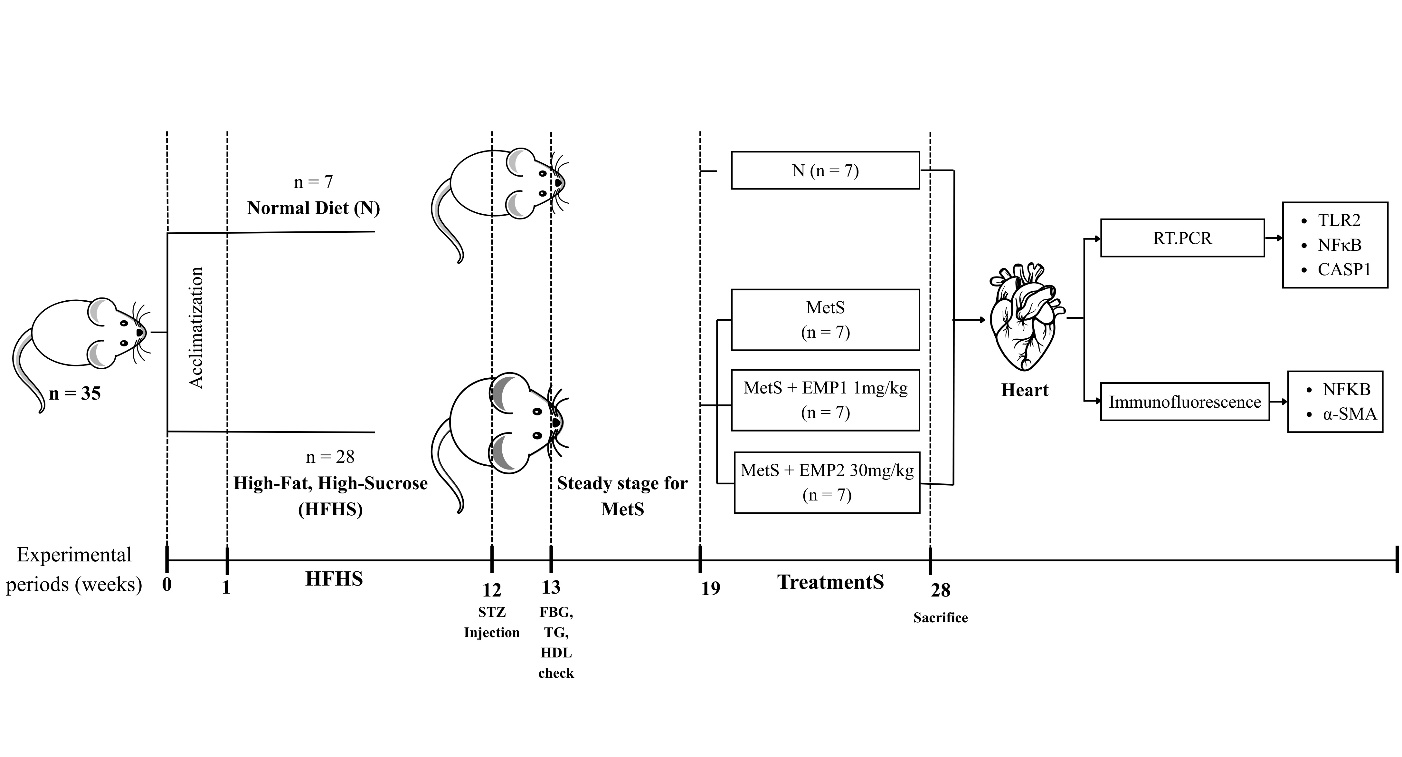
Figure 1 Diagram of animal experimental groups. HFHS induction for 11 weeks, continued with STZ injection (30 mg/kg BW). Fasting blood glucose (FBG), triglycerides (TG), high-density lipoprotein (HDL) were evaluated a week after STZ injection. Steady stage was performed to confirm the MetS state. After that, treatments were given for 9 weeks. Heart was removed and isolated for RT-PCT and immunofluorescence assay.
Measurement of fasting blood glucose, triglyceride, and high-density lipoprotein
Following an overnight fast, blood samples were collected from the tail vein. Serum was isolated via centrifugation at (equivalent to 4,500 rpm) for 15 min at 4 °C. Biochemical parameters, including fasting blood glucose (FBG), triglycerides (TG), and high-density lipoprotein (HDL), were quantified using commercial enzymatic kits (Biolabo, Maizy, France; Cat. Nos. 87409, LP80519, and 86516, respectively) according to the manufacturer's protocols. Absorbance was measured at 500 nm using a spectrophotometer. To ensure accuracy, all samples were analyzed in triplicate, and the mean values were used for subsequent statistical analysis [17].
Determination of collagen volume fraction
Cardiac tissues were harvested and rinsed three times with phosphate-buffered saline (PBS). The tissues were then fixed in 10% neutral buffered formalin, embedded in paraffin blocks, and sectioned at a thickness of 5 µm [18]. To evaluate interstitial fibrosis, the sections were stained using a Masson’s Trichrome kit (PanReac AppliChem, Barcelona, Spain). Digital images were captured using a light microscope (Olympus BX-51, Japan), and the collagen volume fraction (CVF) was quantified via ImageJ software. CVF was calculated as the ratio of the blue-stained collagen area to the total myocardial area, excluding endocardial and perivascular collagen. For each sample, fifteen random fields of view were analyzed in a blinded manner by two independent [19].
RNA isolation and reverse transcription
Cardiac tissues were isolated, minced, and immediately immersed in the RNA stabilization buffer to preserve transcript integrity. Total RNA was extracted using the Zymo Research Quick-RNATM MiniPrep Plus extraction kit (Zymo Research, Irvine, CA, USA) in accordance with the manufacturer’s protocol and stored at °C. This extraction kit is known to be effective in obtaining high quality RNA samples. The extracted RNA samples were then assessed for their quantity and purity using a Nanodrop spectrophotometer at the 260/280 nm absorbance ratio, with values between 1.8 and 2.0 signifying high-quality samples (a). The conversion of RNA into complementary DNA (cDNA) was performed via a reverse transcription (RT-PCR) process, utilizing reverse transcriptase enzyme to synthesize cDNA from the RNA template. The reverse transcription reaction using Toyobo ReverTra AceTM qPCR RT Master Mix (Toyobo, Osaka, Japan). This reaction was prepared in a mixture containing tissue sample, 4 x DNA master mix, gDNA remover, nucleus-free water, and 2 µL 5× RT master mix II. The synthesized cDNA products were kept at −20 °C until required for subsequent RT-PCR assays.
Genes Primer Design
The primer sequences for the TLR2 gene, NF-κB gene, Caspase-1 gene, and the housekeeping gene ACTB were designed based on sequences that obtained from GenBank database (https://www.ncbi.nlm.nih.gov). The primers were specifically designed to ensure the selective amplification on the target DNA fragments. The primer sequences were analyzed using the Molecular Evolutionary Genetics Analysis (MEGA) software version 11. Both forward and reverse primer design for TLR2 gene (GenBank accession number: NM_198769) were positioned in exon 2, ensuring that the targeted DNA fragment corresponded to the protein coding regions. Similarly, the primers for the NF-κB gene (GenBank accession number: NM_001276711) were positioned in exon 7 and exon 8. Whereas the primers for the Caspase-1 gene (GenBank accession number: NM_012762) were positioned in exon 4 and exon 5. On the other hand, the control gene ACTB (GenBank accession number: NM_031144) were positioned in exon 5 and exon 6. By strategically designing primers in the exon region facilitates the amplification process can selectively target the desired DNA fragments for subsequent RT-PCR assays (Table 1).
Table 1 Gene primer sequence for RT-PCR.
Gene |
Gene number |
Strand |
Sequence (5’→3’) |
Size (bp) |
Annealing temperature (°C) |
TLR2 |
NM_198769 |
Forward |
GGGAAATCCTTACAGGACAC |
143 |
60 |
|
|
Reverse |
CAAGTTCGTTGAGAGAGGTC |
|
|
NF-κB |
NM_001276711 |
Forward |
CCAACTTCTCCGACAGTTTC |
187 |
60 |
|
|
Reverse |
CCTGTGTTGGATTTAGTGGC |
|
|
Caspase-1 |
NM_012762 |
Forward |
CACGTCTTGCCCTCATTATC |
202 |
60 |
|
|
Reverse |
CTGTCAGAAGTCTTGTGCTC |
|
|
ACTB |
NM_031144 |
Forward |
CTATGTTGCCCTAGACTTCG |
172 |
59-60 |
|
|
Reverse |
GATGCCACAGGATTCCATAC |
|
|
Real-time PCR (RT-PCR)
RT-PCR was performed in a total reaction volume comprising 10 µL Toyobo ThunderBird, 8 µL nucleous-free water (NFW), 0.5 µL forward primer, and 0.5 µL reverse primer. Amplification was carried out with pre-denaturation at 95 °C for 1 min, denaturation at 95 °C for 15 s, annealing at 60 °C for 30 s, extension at 95 °C for 5 s, and final extension at 95 °C for 30 s. The amplification process on the RT-PCR machine lasted for 40 cycles. The experimental outputs included amplification graph, melt curve graph, melt peak graph, and cycle quantification values.
The quantification values of the TLR2 gene, NF-κB gene, and Caspase-1 gene expression were calculated using the formula 2-∆∆Ct which is used to compare the CT value of the target gene with control gene (ACTB) (b). The ∆CT value was calculated as the difference between the CT of the target gene and CT of the ACTB. Subsequently, ∆∆CT value was calculated as the difference between ∆CT of the target gene and ∆CT of the ACTB gene (c). The result of the differences between the target gene and control gene was calculated as fold change.
Immunofluorescence of α-SMA and NFKB subunit P65
Briefly, cardiac tissue sections were baked at 60 °C for 1 h, deparaffinized in xylene, and rehydrated through a graded ethanol series. Antigen retrieval was performed by immersing the slides in citrate buffer (M, pH 6.0) at 100 °C for 20 min, followed by three washes with TBS-T. Non-specific binding was blocked with 3% BSA for 2 h in the dark. The sections were then double-stained with FITC-conjugated anti-NFκB p65 (Santa Cruz, sc-8008) and Cy3-conjugated anti--smooth muscle actin (-SMA) (Sigma-Aldrich, c6198) and incubated overnight at 4 °C. After three subsequent washes in TBS-T, nuclei were counterstained with DAPI (Sigma, D9542). Images were captured using an Olympus IX51 fluorescence microscope (Olympus, Tokyo, Japan) equipped with Fluoview software. The exposure of fluorochrome light was same at all slides as blinded factor. The co-localization of NFκB and α-SMA was quantified using ImageJ [20]. As the blinded manner, the quantification of NFκB and α-SMA were evaluated at five area in the cardiac smooth muscle.
Data analysis
Statistical analysis was performed using GraphPad Prism 9 (GraphPad Software, LLC, San Diego, CA, USA). Data are presented as mean and standard deviation (SD). The normality of distribution and homogeneity of variance were assessed using Shapiro Wilk and Levene’s tests, respectively (P > 0.05). Differences between experimental groups were analyzed using one-way analysis of variance (ANOVA), followed by Tukey’s Honestly Significant Difference (HSD) post-hoc test for multiple comparisons. Statistical significance was defined with P < 0.05.
Results and discussion
Effect of empagliflozin on the anthropometric body and biochemistry of MetS rats
The anthropometric characteristics of the experimental groups are presented in Figures 2(A) - 2(C). Empagliflozin (EMP) administration at 1 mg/kg significantly attenuated body weight gain compared to the untreated MetS group (P < 0.05), whereas the 30 mg/kg dose showed no significant difference (Figure 2(A)). The body length differed significantly between the healthy control and MetS groups (Figure 2(B)), and the abdominal circumference was markedly increased in all MetS rats compared with healthy control. Notably, the high-dose EMP group exhibited the greatest abdominal circumference among all experimental groups (Figure 2(C)). Following the 13-week HFHS diet and STZ induction, the MetS group displayed significantly elevated fasting blood glucose and triglyceride levels compared to the healthy controls (P < 0.05); (Figures 2(D) - 2(E)). Conversely, HDL levels were significantly reduced in the MetS group; however, EMP treatment at both doses effectively restored HDL levels, exceeding those observed in both the MetS and healthy control groups (Figure 2(A)).
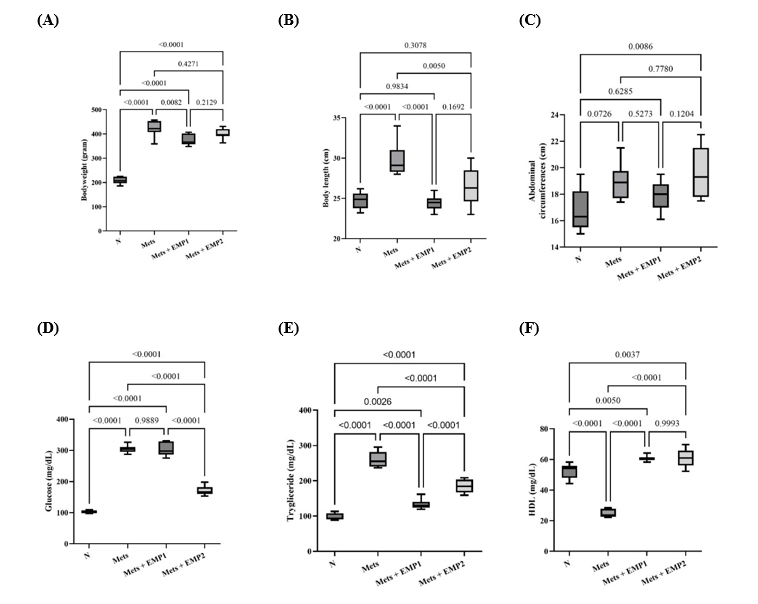
Figure 2 Anthropometric parameters and biochemical profiles of experimental groups. (A) Body weight, (B) body length, (C) abdominal circumference, (D) fasting blood glucose, (E) triglycerides, and (F) high-density lipoprotein (HDL) levels measured after 12 weeks of treatment. Data are presented as the mean standard deviation (per group). Statistical significance was determined using one-way ANOVA followed by Tukey HSD, with considered significant. N: normal healthy control; MetS: untreated metabolic syndrome; EMP1: MetS + 1 mg/kg empagliflozin; EMP2: MetS + 30 mg/kg empagliflozin.
A high-fat, high-sucrose (HFHS) diet is a primary driver of metabolic syndrome (MetS), characterized by central obesity and insulin resistance, which predisposes individuals to type 2 diabetes and adverse cardiac remodeling [21,22]. Chronic HFHS intake is inextricably linked to oxidative stress and a systemic inflammatory response. This metabolic stress triggers an excessive release of pro-inflammatory cytokines, facilitating the infiltration of inflammatory cells—a critical phase in the progression of tissue damage [23]. Previous studies have established that lipid and sugar overload significantly upregulates the TLR2/NFκB signaling pathway, which serves as a central orchestrator of the inflammatory cascade and subsequent tissue remodeling [24,25]. Thus, the MetS model developed in this study aligns with established protocols [16,17,26,27]. Crucially, the clinical burden of MetS extends beyond standard biomarkers like body weight or glucose levels; rather, it is the chronic inflammatory milieu that induces pathological cardiac fibrosis. Consequently, inhibiting the inflammatory mechanisms inherent in MetS is essential to attenuate fibrosis and mitigate the risk of major adverse cardiovascular events.
Our findings demonstrate that the HFHS diet combined with STZ induction effectively mirrors the dynamic phenotypic and microenvironmental shifts characteristic of metabolic syndrome. The observed changes in body weight, abdominal circumference, and metabolic parameters are consistent with established MetS criteria [19,31]. Notably, EMP administration exerted significant therapeutic effects across these markers. In line with previous reports, EMP reduced body weight and adipose tissue mass while improving the lipid profile [28]. The observed increase in HDL levels is particularly noteworthy; similar to findings with canagliflozin, this may reflect a shift toward larger HDL subclasses, which are associated with cardiovascular protection [29]. This suggests that HDL modulation may be a class-specific effect of SGLT2 inhibitors in metabolic syndrome.
Intriguingly, while the 30 mg/kg dose significantly reduced fasting plasma glucose (FPG), the 1 mg/kg dose—which reflects the human equivalent dose of 10 mg/day—showed no significant difference in FPG compared to the untreated MetS group. This indicates that the beneficial outcomes observed at 1 mg/kg are largely independent of its glucose-lowering capacity, suggesting potent pleiotropic effects. Furthermore, the higher dose provided no additional benefit in terms of blood pressure, body weight, or HDL levels. Most significantly, the lower dose (1mg/kg) outperformed the high dose (30 mg/kg) in reducing triglycerides, TLR2 expression, NFKB p65 translocation, and the collagen volume fraction (CVF). These results suggest that a clinically relevant dose may be more effective at targeting the inflammatory and fibrotic axis than supra-therapeutic doses.
Empagliflozin improves histopathological changes in MetS rats
Cardiac tissue examination using HE and MT staining suggested that cardiac disarrangements and fibrosis were found in the MetS group, while EMP ameliorates these pathological changes (Figure 3(A)). HE staining indicated an increase in fractured muscle fibers, loose muscle with wide gaps, necrosis, and hemorrhage in the MetS groups. Additionally, MT staining demonstrated greater fibrosis in the interstitial regions of the myocardium in the MetS groups than in normal healthy groups. Our findings showed that EMP administration reduce cardiac fibrosis significantly (P < 0.05) in MetS rats (Figure 3(A)). Histopathological examination using H&E and Masson’s Trichrome (MT) staining revealed significant structural disorganization and fibrosis within the myocardium of the MetS group, which were effectively attenuated by EMP treatment (Figure 3(A)). H&E staining in MetS rats showed characteristic pathological features, including fractured muscle fibers, myofibrillar disarray with widened interstitial gaps, focal necrosis, and areas of hemorrhage. Furthermore, MT staining confirmed significantly higher collagen deposition in the interstitial regions of the myocardium in the MetS group compared to healthy controls. Quantitative analysis of the MT sections demonstrated that EMP administration significantly reduced the cardiac collagen volume fraction (P < 0.05) in MetS rats (Figure 2(B)).
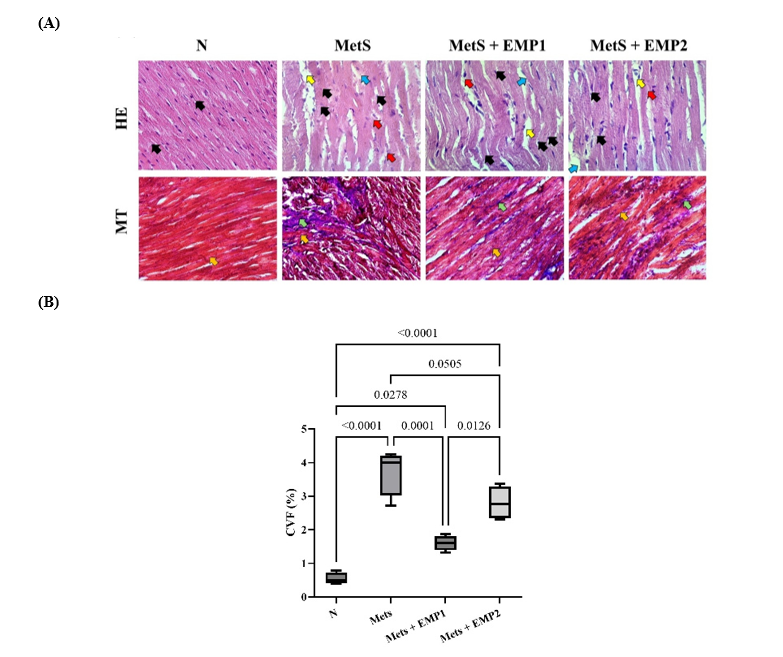
Figure 3 Histopathological changes and collagen deposition in myocardial tissue. (A) Representative H&E-stained sections showing myocardial morphology. Black arrows indicate fractured muscle fibers; yellow arrows point to widened interstitial gaps between myocytes; black arrows denote focal necrosis; and red arrows highlight areas of hemorrhage. Masson’s Trichrome (MT) staining was used to visualize collagen (blue, green arrows) against the cytoplasm (red, yellow arrows) and nuclei (black). (B) Quantitative analysis of the collagen volume fraction (CVF) expressed as a percentage. Data represent the mean standard deviation from six random fields of view per sample at 400x magnification. Statistical significance was determined using one-way ANOVA followed by Tukey HSD, where was considered significant. N: normal healthy control; MetS: untreated metabolic syndrome; EMP1: MetS + 1 mg/kg EMP; EMP2: MetS + 30 mg/kg EMP.
Histopathological analysis in the present study confirms that EMP treatment effectively ameliorates cardiac fibrosis induced by HFHS and STZ. While previous research has shown that EMP reduces the collagen volume fraction in non-diabetic myocardial infarction (MI) models via the TGF-/Smad3 [15], Nrf2/HO-1 [30] or mitochondrial pathways [31]. The pathophysiology of fibrosis in MI differs fundamentally from that in MetS. In MI, cardiac fibrosis is primarily a ‘replacement’ process - a localized wound-healing response to replace necrotic myocytes following an acute event. In contrast, fibrosis in our MetS model is 'reactive interstitial fibrosis,' characterized by a diffuse and disproportionate accumulation of collagen throughout the myocardium in response to chronic metabolic triggers.
Empagliflozin Downregulates TLR2, NFKB, CASP1 Expression in MetS Rats
To elucidate the molecular mechanisms underlying the anti-fibrotic effects of EMP, we quantified the mRNA expression of key components within the TLR2/NFκB signaling pathway. Our results demonstrated a significant upregulation of TLR2 and NFKB in the MetS group, suggesting pathway activation; however, EMP administration significantly suppressed the expression of these markers (Figure 4). We further examined CASP1, a central protein in the inflammasome pathway and a known mediator of fibrotic development. While CASP1 expression was elevated in MetS rats compared to healthy controls, EMP treatment failed to significantly reduce its expression (Figure 4), indicating that the drug's anti-fibrotic effect in this model may be independent of the CASP1-inflammasome axis.
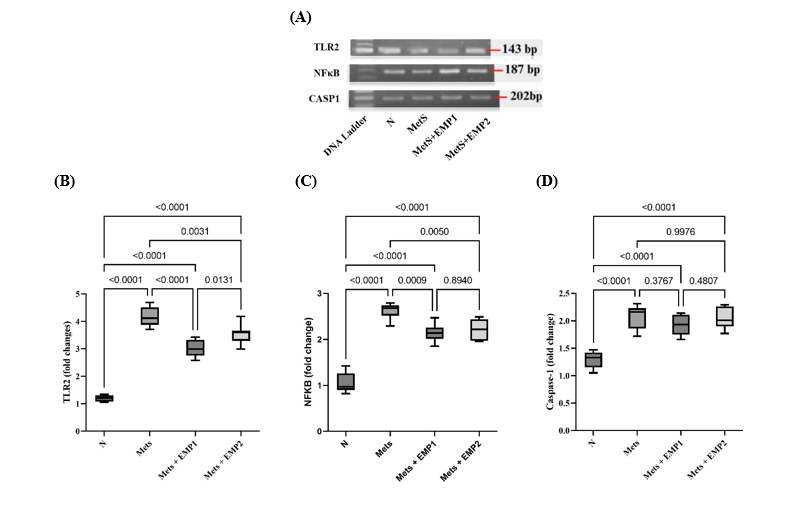
Figure 4 Effect of EMP on the expression of cardiac fibrosis-related genes in MetS rats. (A) Representative 1.5% agarose gel electrophoresis images of PCR products for TLR2 (143 bp), NFKB (187 bp), and CASP1 (202 bp). Relative mRNA expression levels of (B) TLR2, (C) NFKB, and (D) CASP1 were normalized to GAPDH as the internal control. Gel images were captured using the Glite 965 GW Gel Documentation System. Data are expressed as the mean standard deviation (per group). Statistical significance was determined using one-way ANOVA followed by Tukey HSD, with considered significant. N: normal health control; MetS: untreated metabolic syndrome; EMP1: MetS + 1 mg/kg EMP; EMP2: MetS + 30 mg/kg EMP.
Our findings suggest that targeting the TLR2/NFκB signaling axis is a viable strategy for improving this reactive fibrosis. The TLR2/NFκB pathway is a central mediator of inflammatory and fibrotic responses across various tissues [32,33]. For instance, inhibiting TLR2 activity has been shown to reduce cardiac dysfunction and inflammation-associated fibrosis by approximately 13% [34]. Furthermore, TLR2 deficiency has been reported to inhibit Angiotensin II-mediated remodeling via the MyD88 interaction [35,36]. Consistently, our data demonstrate that the downregulation of TLR2 leads to a concomitant decrease in NFKB activity, thereby alleviating the inflammation and hypertrophy that drive myocardial fibrosis [9].
To further investigate whether the observed structural alterations were driven by the TLR2/NFκB signaling axis, we performed dual immunofluorescence staining for α-smooth muscle actin (α-SMA) and the nuclear-translocated NFκB p65 subunit (Figure 5(A)). As a primary marker of myofibroblast differentiation, α-SMA expression serves as a definitive hallmark of active cardiac fibrosis. Our findings revealed a marked increase in both α-SMA expression and the nuclear translocation of NFκB p65 in the myocardium of MetS rats compared to healthy controls. Notably, EMP treatment significantly diminished the expression and co-localization of these markers within the cardiac tissue (Figure 5(B)), suggesting that EMP mitigates fibrosis by suppressing NFκB-mediated myofibroblast activation.
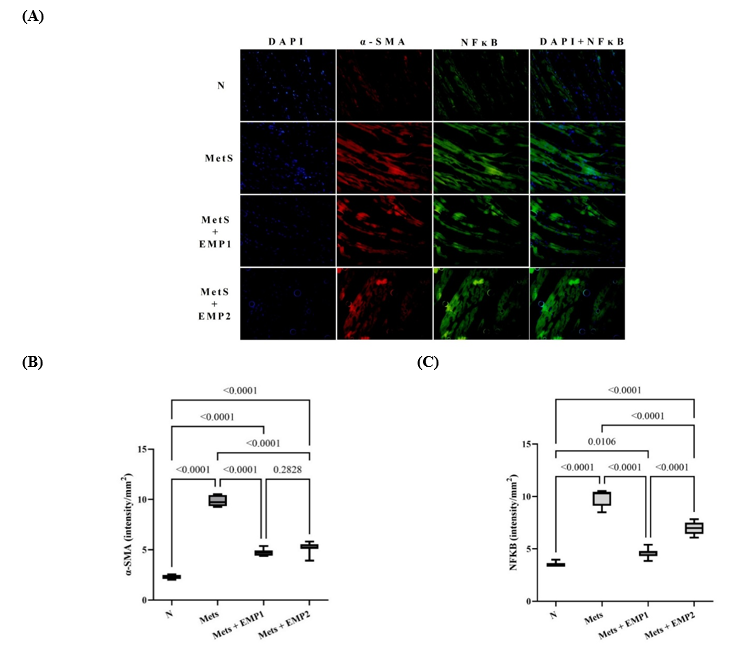
Figure 5 Effect of EMP on myocardial expression of -SMA and NFκB. (A) Representative immunofluorescence images of cardiac sections dual-stained for α-SMA (red) and NFκB p65 (green); nuclei were counterstained with DAPI (blue). Quantitative analysis of (B) α-SMA and (C) NFκB p65 fluorescence intensity. Empagliflozin (EMP) treatment significantly suppressed the expression of both markers (P < 0.05). Data are presented as mean standard deviation. Fluorescence intensity was quantified using ImageJ software from six random fields per section at 400× magnification (40× objective). Statistical significance was determined via one-way ANOVA followed by Tukey HSD (P < 0.05). N: normal healthy control; MetS: untreated metabolic syndrome; EMP1: MetS + 1 mg/kg EMP; EMP2: MetS + 30 mg/kg EMP.
Our findings showed that EMP did not significantly alter CASP1 expression in this specific MetS model. This suggests that the anti-fibrotic benefits of EMP observed in our study may be predominantly mediated through the TLR2/NFκB pathway rather than the canonical Caspase-1-dependent pyroptotic pathway. Caspase-1 (CASP1) is a well-established mediator of pyroptosis—a form of programmed cell death characterized by the excessive release of pro-inflammatory cytokines, which triggers a robust systemic inflammatory response. In type 2 diabetes, accelerated myocardial cell death is a fundamental driver of diabetic cardiomyopathy and coronary heart disease [37]. This discrepancy may be attributed to several regulatory factors. For instance, post-transcriptional modifications or allosteric regulation may serve as the dominant factors in determining Caspase-1 enzymatic activity, rather than changes at the transcript level alone. Furthermore, the activation of CASP1 via the NLRP3 inflammasome is a multifactorial process; beyond the TLR2/NFκB signaling axis, it is heavily influenced by endoplasmic reticulum (ER) stress, oxidative stress, and mitochondrial dysfunction [38]. Consequently, the inability of EMP to reduce CASP1 expression in this specific model suggests that these alternative pathways might maintain CASP1 levels despite the suppression of the TLR2/NFκB-mediated inflammatory response.
Cardiac fibrosis is fundamentally driven by the phenotypic differentiation of cardiac fibroblasts into active myofibroblasts. The expression of α-SMA serves as a definitive biomarker for these myofibroblasts and is an essential component in the progression of myocardial structural remodeling [39]. Our findings demonstrate that EMP treatment significantly attenuates α-SMA expression within the cardiac tissue. These results align with previous studies in renal models, where EMP was found to inhibit hyperglycemia-induced epithelial-mesenchymal transition (EMT) and restore epithelial markers such as E-cadherin, largely by suppressing the TGF-1/SMAD3 signaling pathway [40]. In the present study, NF-κB pathway activity was evaluated through immunofluorescence-based assessment of p65 nuclear translocation. Activation of NF-κB is primarily characterized by cytoplasmic-to-nuclear translocation of the p65 subunit rather than solely by changes in total protein abundance. Therefore, visualization of subcellular localization provides mechanistically relevant insight into pathway activation status. Immunofluorescence enables spatial resolution within cardiac tissue and direct evaluation of nuclear localization at the cellular level. Thus, the observed reduction in p65 nuclear translocation following empagliflozin treatment supports attenuation of NF-κB signaling activity in this experimental model. Intriguingly, our data suggests that EMP exerts a dual therapeutic role: while it effectively manages glycemic levels, it also possesses significant pleiotropic effects that provide cardiovascular protection by inhibiting the TLR2/NFκB inflammatory axis. The systemic blockade of NFκB activation has been well-documented to reduce key pro-inflammatory mediators, including TNF-α, IL-6, hs-CRP, and MCP-1, thereby mitigating the chronic inflammatory state associated with metabolic syndrome [41].
In this study, EMP at a dose of 1 mg/kg/day—the rat equivalent of the 10 mg/day human dose approved by the ACC/AHA for heart failure—yielded results that were largely comparable to the supra-therapeutic dose of 30 mg/kg/day (approximately 300 mg/day in humans). These findings suggest a 'ceiling effect' regarding the anti-inflammatory and anti-fibrotic properties of empagliflozin, where a 1 mg/kg/day dose appears sufficient to optimally inhibit the TLR2/NF-κB axis. A similar plateau has been observed in the pharmacodynamics of urinary glucose excretion, which reaches a ceiling at doses between 100 - 400 mg/day in humans [42]. This phenomenon may be explained by pharmacodynamic target saturation, wherein the receptors or signaling components targeted by EMP are fully occupied at lower concentrations, rendering further dose increases non-linear in their therapeutic impact. This aligns with the pharmacological principle that efficacy does not always scale proportionally with dose once the maximum biological response is achieved. Furthermore, factors such as saturated bioavailability or limited tissue distribution may play a role; specifically, the metabolic pathways or transport mechanisms for EMP may become saturated, preventing higher oral doses from translating into significantly increased concentrations within the myocardial tissue.
Interestingly, the improvements in collagen volume fraction (CVF) and the suppression of the TLR2/NFκB p65 axis were more pronounced at the 1mg/kg/day dose than at 30 mg/kg/day. These findings suggest that the anti-fibrotic properties of low-dose EMP (the human equivalent of 10 mg/day) may follow a non-monotonic or U-shaped dose-response curve. This phenomenon has been previously noted in human studies, where increasing EMP doses beyond 100 - 400 mg/day resulted in a plateau or a relative decline in urinary glucose excretion efficiency [42]. At supra-therapeutic doses, the disruption of metabolic homeostasis may counteract the drug’s primary benefits. For instance, high-dose EMP can induce excessive osmotic diuresis, leading to dehydration and significant plasma volume depletion. Such hypovolemia may trigger compensatory systemic stress responses, including the activation of the renin-angiotensin-aldosterone system (RAAS) and increased oxidative stress, which could ultimately attenuate the inherent anti-inflammatory and anti-fibrotic effects of the drug [43], An alternative explanation for the diminished efficacy of the 30mg/kg/day dose involves the potential loss of SGLT isoform selectivity. While empagliflozin is highly selective for SGLT2, supra-therapeutic doses (30-80 mg/kg/day) may lead to the off-target inhibition of SGLT1, which is prominently expressed in cardiomyocytes [44]. Under conditions of metabolic stress, such as hypoxia or hypoglycemia, SGLT1 expression is often up regulated as a compensatory mechanism to maintain glucose uptake and energy homeostasis within the heart. Consequently, the non-selective inhibition of SGLT1 by high-dose EMP may disrupt this protective energetic adaptation, thereby compromising the drug's anti-fibrotic potential. This mechanism likely explains why the reductions in TLR2, NFKB, and collagen volume fraction (CVF) were less significant at 30 mg/kg/day compared to the more targeted 1 mg/kg/day dose in our metabolic syndrome model.
Furthermore, our findings are consistent with current clinical guidelines from the American Heart Association (AHA), American College of Cardiology (ACC), and Heart Failure Society of America (HFSA), which recommend a standard human dose of 10 mg/day. This corresponds to the 1 mg/kg/day dose used in our rat model. Notably, the 1 mg/kg/day regimen demonstrated greater therapeutic benefit than the higher 30 mg/kg/day dose, suggesting that increasing the dose did not yield additional efficacy in this experimental setting. Clinical research by Wu et al. confirms that empagliflozin at daily doses of 10, 25, and 50 mg effectively reduces fasting plasma glucose (FPG) and HbA1c in patients with type 2 diabetes mellitus (T2DM). Moreover, Jiang et al. reported that standard doses of 10 or 25 mg/day significantly lower mortality rates compared to placebo and exhibit a superior safety and tolerability profile compared to other SGLT2 inhibitors [45]. Forycka et al. [47] similarly demonstrated that EMP-mediated reduction of cardiac fibrosis in T2DM patients effectively lowers mortality stemming from both renal and heart failure. By adhering to the recommended 10 mg dose, clinicians can achieve substantial reductions in heart failure hospitalizations and cardiovascular mortality while minimizing the risk of adverse side effects.
In summary, our findings demonstrate that nine-week empagliflozin (EMP) administration not only improved metabolic parameters but also attenuated cardiac fibrotic remodeling in a rat model of metabolic syndrome. Mechanistically, EMP reduced activation of the TLR2/NF-κB inflammatory signaling axis, as evidenced by downregulation of TLR2 and NF-κB gene expression and, critically, diminished nuclear translocation of the NF-κB p65 subunit within cardiac tissue. Because NF-κB activation is defined by p65 nuclear localization, these results provide functional evidence that EMP suppresses inflammatory signaling activity in the myocardium. The coordinated reduction in myofibroblast activation (α-SMA), and collagen deposition further supports a mechanistic link between NF-κB attenuation and reduced fibrotic remodeling.
Nevertheless, several limitations should be acknowledged. First, although nuclear translocation of p65 reflects functional NF-κB activation, complementary quantitative protein analyses (e.g., total or phosphorylated NF-κB levels) were not performed. Second, downstream inflammatory cytokines such as TNF-α, IL-1β, and IL-6 were not directly measured, limiting comprehensive characterization of the inflammatory cascade. Third, pathway-specific intervention approaches (e.g., genetic knockdown or pharmacological inhibition of TLR2/NF-κB) were not employed, and thus definitive causal hierarchy within the signaling network cannot be fully established. Fourth, functional cardiac assessments such as echocardiography were not conducted, precluding direct evaluation of whether reduced fibrosis translated into measurable improvements in cardiac performance. Finally, cardiac fibrosis is inherently multifactorial, and additional mechanisms—including oxidative stress modulation, metabolic remodeling, and non-canonical inflammatory pathways—may contribute to the observed effects.
Despite these limitations, the present data substantiate modulation of the TLR2/NF-κB axis as a key component of EMP’s cardioprotective action in this experimental model. Given the shared inflammatory and fibrotic mechanisms between metabolic syndrome and heart failure, these findings provide mechanistic insight into the broader cardiometabolic benefits of SGLT2 inhibition.
Conclusions
In conclusion, metabolic syndrome in this model is characterized by activation of the TLR2/NF-κB signaling axis and subsequent cardiac fibrotic remodeling. Empagliflozin attenuated these changes, as evidenced by reduced TLR2 and NF-κB expression, diminished nuclear translocation of the NF-κB p65 subunit, decreased myofibroblast activation, and lower collagen deposition. Given that NF-κB activation is defined by p65 nuclear localization, these findings provide functional mechanistic evidence that empagliflozin suppresses myocardial inflammatory signaling.
Notably, the clinically relevant low dose (1 mg/kg/day) demonstrated superior antifibrotic effects compared with the higher dose (30 mg/kg/day). Although complementary protein quantification, downstream cytokine profiling, and functional cardiac assessments were not performed, the coordinated reduction in inflammatory activation and fibrosis substantiates modulation of the TLR2/NF-κB axis as a central component of empagliflozin’s cardioprotective effects in metabolic syndrome. Future studies incorporating pathway-specific intervention strategies and functional cardiac evaluation are warranted to further define the causal hierarchy and translational impact of these findings.
Declaration of generative AI in scientific writing
The authors acknowledge that artificial intelligence (AI) tool was used solely for language editing (Google Gemini). This tool did not generate, analyze, or alter any scientific content, results, or conclusions presented in this manuscript.
CRediT author statement
Muhammad Gibran Fauzi Harmani Kalim: Conceptualization, Investigation, Formal analysis, Resources, Writing - Original Draft; Mohammad Saifur Rohman: Conceptualization, Methodology, Supervision, Validation, Writing - Review & Editing; Djanggan Sargowo: Supervision, Validation, Writing - Review & Editing; Sri Winarsih: Supervision, Validation, Writing - Review & Editing; Dian Nugrahenny: Methodology, Supervision, Validation, Writing - Review & Editing; Hidayat Sujuti: Methodology, Supervision, Validation, Writing - Review & Editing; Siska Nanda Widhaningrum: Investigation and Formal analysis; Janjte Wiliem Souhaly: Investigation and Formal analysis.
References
[1] MS Islam, P Wei, M Suzauddula, I Nime, F Feroz, M Acharjee and F Pan. The interplay of factors in metabolic syndrome: understanding its roots and complexity. Molecular Medicine 2024; 30(1), 279.
[2] D Siriwan, P Trisonthi, P Temviriyanukul, P Phansuea, Y Sahasakul and W Inthachat. Effects of different Morus alba L. parts on glycemic and lipid profiles: A systematic review and meta-analysis. Trends in Sciences 2026; 23(4), 12148.
[3] KB Cokan, Ž Urlep, M Moškon, M Mraz, XY Kong, W Eskild, D Rozman, P Juvan and T Režen. Common transcriptional program of liver fibrosis in mouse genetic models and humans. International Journal of Molecular Sciences 2021; 22(2), 832.
[4] W Arozal, M Louisa and V Soetikno. Selected Indonesian medicinal plants for the management of metabolic syndrome: molecular basis and recent studies. Frontiers in Cardiovascular Medicine 2020; 7, 82.
[5] EH Herningtyas and TS Ng. Prevalence and distribution of metabolic syndrome and its components among provinces and ethnic groups in Indonesia. BMC Public Health 2019; 19(1), 377.
[6] W Jiang, Y Xiong, X Li and Y Yang. Cardiac fibrosis: Cellular effectors, molecular pathways, and exosomal roles. Frontiers in Cardiovascular Medicine 2021; 8, 715258.
[7] K Maruyama and K Imanaka-Yoshida. The pathogenesis of cardiac fibrosis: a review of recent progress. International Journal of Molecular Sciences 2022; 23(5), 2617.
[8] TA Wynn and TR Ramalingam. Mechanisms of fibrosis: Therapeutic translation for fibrotic disease. Nature Medicine 2012; 18(7), 1028-1040.
[9] C Qian, D Xu, J Wang, Y Luo, T Jin, L Huang, Y Zhou, Z Cai, B Jin, H Bao and Y Wang. Toll-like receptor 2 deficiency ameliorates obesity-induced cardiomyopathy via inhibiting NF-κB signaling pathway. International Immunopharmacology 2024; 128, 111551.
[10] J Qian, S Liang, Q Wang, J Xu, W Huang, G Wu and G Liang. Toll-like receptor-2 in cardiomyocytes and macrophages mediates isoproterenol-induced cardiac inflammation and remodeling. FASEB Journal 2023; 37(2), e22740.
[11] H Van Tin, L Rethi, S Higa, YH Kao and YJ Chen. Spike protein of SARS-CoV-2 activates cardiac fibrogenesis through NLRP3 inflammasomes and NF-κB signaling. Cells 2024; 13(16), 1331.
[12] MD Molla, Y Akalu, Z Geto, B Dagnew, B Ayelign and T Shibabaw. Role of caspase-1 in the pathogenesis of inflammatory-associated chronic noncommunicable diseases. Journal of Inflammation Research 2020; 13, 749-764.
[13] Y Liu, H Lei, W Zhang, Q Xing, R Liu, S Wu, Z Liu, Q Yan, W Li, X Liu and Y Hu. Pyroptosis in renal inflammation and fibrosis: current knowledge and clinical significance. Cell Death & Disease 2023; 14(7), 472.
[14] M Elsayed, YM Moustafa, ET Mehanna, RA Elrayess, NM El-Sayed and RM Hazem. Empagliflozin protects against isoprenaline-induced fibrosis in rat heart through modulation of TGF-β/SMAD pathway. Life Sciences 2024; 337, 122354.
[15] E Daud, O Ertracht, N Bandel, G Moady, M Shehadeh, T Reuveni and S Atar. The impact of empagliflozin on cardiac physiology and fibrosis early after myocardial infarction in non-diabetic rats. Cardiovascular Diabetology 2021; 20(1), 132.
[16] MS Rohman, IN Chomsy, SN Kurniawan and D Nugrahenny. Green tea and green coffee therapy for aortic calcification prevention in metabolic syndrome model rats: effects on expression of AKT, mTOR, RUNX2, and osteopontin levels. Trends in Sciences 2024; 21(10), 8204.
[17] IN Chomsy, MS Rohman, H Khotimah, N Widodo and NIP Nugrahini. Decaffeinated green tea and green coffee extracts as metformin's add-on enhance metabolic syndrome risk factors and improve the cardiac insulin-gene-related pathway. Journal of Pharmacy & Pharmacognosy Research 2023; 11(3), 414-425.
[18] H Yang, Y Shi, H Liu, F Lin, B Qiu, Q Feng, Y Wang and B Yang. Pyroptosis executor gasdermin D plays a key role in scleroderma and bleomycin-induced skin fibrosis. Cell Death Discovery 2022; 8(1), 183.
[19] E Arteaga, AQ de Araújo, M Bernstein, FJA Ramires, BM Ianni, F Fernandes, C Mady. Valor prognóstico da fração de volume de colágeno na cardiomiopatia hipertrófica. Arquivos Brasileiros de Cardiologia 2009; 92(3), 210-214.
[20] PH Riyadi, R Romadhon, AD Anggo, MF Atho'illah and M Rifa'i. Tilapia viscera protein hydrolysate maintain regulatory T cells and protect acute lung injury in mice challenged with lipopolysaccharide. Journal of King Saud University - Science 2022; 34(5), 102020.
[21] T Delmont, S Luce, C Henique, A Tarraf, Y Zhang, T D'Humières, G Derumeaux and L Bonnet. Impact of a switch diet after high-fat diet induced obesity combined with aging on cardiac function. Archives of Cardiovascular Diseases 2024; 117(6-7), S215.
[22] J Kruszewska, A Cudnoch-Jedrzejewska and K Czarzasta. Remodeling and fibrosis of the cardiac muscle in the course of obesity - pathogenesis and involvement of the extracellular matrix. International Journal of Molecular Sciences 2022; 23(8), 4195.
[23] MRE Wuryandari, MF Atho'illah, RD Laili, S Fatmawati, N Widodo, E Widjajanto and M Rifa'i. Lactobacillus plantarum FNCC 0137 fermented red Moringa oleifera exhibits protective effects in mice challenged with Salmonella typhi via TLR3/TLR4 inhibition and down-regulation of proinflammatory cytokines. Journal of Ayurveda and Integrative Medicine 2022; 13(2), 100531.
[24] MF Atho'illah, YD Safitri, FD Nur'aini, S Widyarti, H Tsuboi and M Rifa'i. Elicited soybean extract attenuates proinflammatory cytokines expression by modulating TLR3/TLR4 activation in high-fat, high-fructose diet mice. Journal of Ayurveda and Integrative Medicine 2021; 12(1), 43-51.
[25] X Zhang, Y Wan, J Feng, M Li and Z Jiang. Involvement of TLR2/4‑MyD88‑NF‑κB signaling pathway in the pathogenesis of intracranial aneurysm. Molecular Medicine Reports 2021; 23(4), 230.
[26] IN Chomsy, MS Rohman, H Khotimah, N Widodo and NIP Nugrahini. Cardioprotective effect of green tea and green coffee extract as metformin’s add-on to prevent cardiac fibrosis in a rat model of metabolic syndrome. Journal of Research in Pharmacy 2024; 28(1), 16-28.
[27] MS Rohman, ANII Idhil, AN Azzah, AP Gunawan, N Widodo, HW Sulistomo, M Lukitasari, DA Nugroho and IN Chomsy. Combination of green tea, green coffee, and turmeric extract improve the THOC5 and AIF1, but not ACTA2 and CNN1 gene expression in the aortic tissue of metabolic syndrome model. Trends in Sciences 2024; 21(11), 8245.
[28] J Trnovska, P Svoboda, H Pelantova, M Kuzma, H Kratochvilova, BJ Kasperova, I Dvorakova, K Rosolova, H Malinska, M Huttl, I Markova, O Oliyarnyk, M Melcova, V Skop, M Mraz, S Stemberkova-Hubackova and M Haluzik. Complex positive effects of SGLT-2 inhibitor empagliflozin in the liver, kidney and adipose tissue of hereditary hypertriglyceridemic rats: Possible contribution of attenuation of cell senescence and oxidative stress. International Journal of Molecular Sciences 2021; 22(19), 10606.
[29] Y Kamijo, H Ishii, T Yamamoto, K Kobayashi, H Asano, S Miake, E Kanda, H Urata and M Yoshida. Potential impact on lipoprotein subfractions in type 2 diabetes. Clinical Medical Insights: Endocrinology Diabetes 2019; 12, 1-8.
[30] P Shi, Z Zhan, X Ye, Y Lu, K Song, F Sheng, H Shen and P Yin. The antioxidative effects of empagliflozin on high glucose‑induced epithelial-mesenchymal transition in peritoneal mesothelial cells via the Nrf2/HO-1 signaling. Renal Failure 2022; 44(1), 1529-1543.
[31] Z Wang, Q Liu, X Wang, P Wang, Z Wang and F Zhang. Empagliflozin improves cardiac function in rats with chronic heart failure. Naunyn-Schmiedeberg’s Archive Pharmacology 2024; 397(2), 1037-1044.
[32] U Bhandari and P Arya. Involvement of the toll-like receptors-2/nuclear factor-kappa B signaling pathway in atherosclerosis induced by high-fat diet and zymosan A in C57BL/6 mice. Indian Journal of Pharmacology 2020; 52(3), 203.
[33] C Yu, D Wang, Z Yang and T Wang. Pharmacological effects of polyphenol phytochemicals on the intestinal inflammation via targeting TLR4/NF-κB signaling pathway. International Journal of Molecular Sciences 2022; 23(13), 6939.
[34] Y Ma, X Zhang, H Bao, S Mi, W Cai, H Yan, Q Wang, Z Wang, J Yan, G Fan, ML Lindsey and Z Hu. Toll-like receptor (TLR) 2 and TLR4 differentially regulate doxorubicin induced cardiomyopathy in mice. PLoS ONE 2012; 7(7), e40763.
[35] L Wang, YL Li, CC Zhang, W Cui, X Wang, Y Xia, J Du and HH Li. Inhibition of toll-like receptor 2 reduces cardiac fibrosis by attenuating macrophage-mediated inflammation. Cardiovascular Research 2014; 101(3), 383-392.
[36] S Ye, K Lin, G Wu, MJ Xu, P Shan, W Huang, Y Wang and G Liang. Toll-like receptor 2 signaling deficiency in cardiac cells ameliorates Ang II-induced cardiac inflammation and remodeling. Translational Research 2021; 233, 62-76.
[37] D Huang, W Gao, X Zhong and J Ge. NLRP3 activation in endothelia promotes development of diabetes-associated atherosclerosis. Aging 2020; 12(18), 18181-18191.
[38] N Ji, Z Qi, Y Wang, X Yang, Z Yan, M Li, Q Ge and J Zhang. Pyroptosis: A new regulating mechanism in cardiovascular disease. Journal of Inflammation Research 2021; 14, 2647-2666.
[39] Y He, S Ling, Y Sun, Z Sheng, Z Chen, X Pan and G Ma. DNA methylation regulates α-smooth muscle actin expression during cardiac fibroblast differentiation. Journal of Cellular Physiology 2019; 234(5), 7174-7185.
[40] A Ndibalema, D Kabuye, S Wen, L Li, X Li and Q Fan. Empagliflozin protects against proximal renal tubular cell injury induced by high glucose via regulation of hypoxia-inducible factor 1-alpha. Diabetes, Metabolic Syndrome and Obesity 2020; 13, 1953-1967.
[41] A Preda, F Montecucco, F Carbone, GG Camici, TF Lüscher, S Kraler and L Liberale. SGLT2 inhibitors: from glucose-lowering to cardiovascular benefits. Cardiovascular Research 2024; 120(5), 443-460.
[42] AJ Scheen, Pharmacokinetic and Pharmacodynamic Profile of Empagliflozin, a Sodium Glucose Co-Transporter 2 Inhibitor. Clinical Pharmacokinetics 2014; 53(3), 213-225.
[43] S Supakul, Y Nishikawa, M Teramura and T Takase. Short-term treatment with empagliflozin resulted in dehydration and cardiac arrest in an elderly patient with specific complications: A case report and literature review. Medicina (Kaunas) 2022; 58(6), 815.
[44] R Grempler, L Thomas, M Eckhardt, F Himmelsbach, A Sauer, DE Sharp, RA Bakker, M Mark, T Klein and P Eickelmann. Empagliflozin, a novel selective sodium glucose cotransporter‐2 (SGLT‐2) inhibitor: characterisation and comparison with other SGLT‐2 inhibitors. Diabetes, Obesity and Metabolism 2012; 14(1), 83-90.
[45] Y Jiang, P Yang, L Fu, L Sun, W Shen, Q Wu. comparative cardiovascular outcomes of sglt2 inhibitors in type 2 diabetes mellitus: A network meta-analysis of randomized controlled trials. Frontiers Endocrinology 2022; 13, 802992.
[46] J Forycka, J Hajdys, J Krzemińska, P Wilczopolski, M. Wronka, E Młynarska, J Rysz and B Franczyk. New insights into the use of empagliflozin: A comprehensive review. Biomedicines 2022; 10(12), 3294.