Trends
Sci.
2026;
23(9):
13457
Chemical Profiling, Antioxidant, and Antibacterial Activities of Ethanolic Extract of Melaleuca cajuputi Flower and Its Mode of Action
Somruedee Yorsin1, Patcharaporn Siwayaprahm2, Watchara Chongsa3 and Tanes Sangsri4,*
1Department of Physiology, Faculty of Medicine, Princess of Naradhiwas University, Narathiwat 96000, Thailand
2Department of Microbiology, Faculty of Science, Kasetsart University, Bangkok 10900, Thailand
3Physiology Unit, Faculty of Science, Rangsit University, Pathum Thani 12000, Thailand
4Department of Microbiology, Faculty of Medicine, Princess of Naradhiwas University, Narathiwat 96000, Thailand
(*Corresponding author’s e-mail: [email protected])
Received: 26 January 2026, Revised: 24 February 2026, Accepted: 3 March 2026, Published: 10 April 2026
Despite well-documented antimicrobial properties of Melaleuca cajuputi extracts, particularly from leaves and essential oils, the antibacterial potential of flower extracts against multidrug-resistant (MDR) pathogens, such as methicillin-resistant Staphylococcus aureus (MRSA), remains largely underexplored. This study characterized the phytochemical composition, antioxidant, and antibacterial activities of ethanol extracts from M. cajuputi flowers. Dried flowers of M. cajuputi were extracted with ethanol, followed by phytochemical analysis using liquid chromatography/mass spectrometry (LC/MS). Antioxidant activity was determined using 1,1-diphenyl-2-picryl hydrazyl (DPPH) radical and 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid (ABTS) radical scavenging assay. Antibacterial activity of the crude extract was assessed using the agar well diffusion method. The minimal inhibitory concentrations (MICs) and minimum bactericidal concentrations (MBCs) were determined by microdilution, including time-kill assays. Scanning electron microscopy (SEM) was employed to investigate the effects of the extract on the morphology of MRSA cells. Phytochemical profiling established that the extract was abundant in phenolics, flavonoids, and tannins. Notable antioxidant capacity was observed, with IC50 values of 11.28 ± 0.30 μg/mL (DPPH) and 61.90 ± 14.84 μg/mL (ABTS). Evaluation against 5 significant bacterial pathogens, including MRSA, revealed broad-spectrum inhibitory effects. The lowest recorded MIC and MBC values were 0.33 and 5.31 mg/mL, respectively, with ranges spanning 0.33 - 125 mg/mL (MIC) and 5.31 - 250 mg/mL (MBC). Time-kill studies indicated concentration- and time-dependent bactericidal mode of action against most strains. At the same time, MRSA was primarily inhibited in a bacteriostatic manner. SEM revealed dose-dependent morphological damage to MRSA cells, including membrane disruption and shrinkage, indicating compromised cell envelope integrity. These findings indicate that M. cajuputi flower extract is a promising candidate for natural antibacterial and antioxidant applications, meriting further investigation for its role in controlling MDR bacterial infections.
Keywords: Melaleuca cajuputi, Antioxidant activity, Antibacterial activity, Liquid chromatography/mass spectrometry (LC/MS), Methicillin-resistant Staphylococcus aureus (MRSA)
Plants have historically served as a rich source of bioactive molecules with antibacterial properties, attracting attention for drug discovery efforts [1,2]. Melaleuca cajuputi, commonly known as Cajuput, Gelam tree, Paper bark, or White Samet, is a species in the family Myrtaceae, widely distributed across
Southeast Asia, Australia, and surrounding regions [3,4]. Its essential oils and extracts have been traditionally used to treat respiratory, skin, and wound infections [4,5]. The White Samet (Melaleuca cajuputi) forest in Narathiwat province represents a vital natural resource with diverse ecological, economic, and cultural significance. Various parts of the tree served multiple valuable roles in traditional and commercial applications. The soft bark provides firewood and materials for household items, the wood supports construction, fuel needs, and the leaves are traditionally brewed as herbal tea or distilled to yield cajuput oil for ointments and insect repellent [6]. Ecologically, White Samet forests play a crucial role in soil conservation, erosion prevention, and providing habitats for wildlife, including aquatic birds and pollinators such as bees, that maintain ecosystem balance [6]. Moreover, as integral components of peat swamp ecosystems, these forests regenerate degraded areas, restore soil fertility, sequester carbon, and mitigate the impacts of climate change.
Multidrug resistance (MDR) imposes a significant economic and public health burden worldwide. For instance, it increases the cost and complexity of medical care, prolongs hospital stays, and reduces the productivity of patients and their caregivers [7]. MDR is a global concern that transcends national borders and affects countries of all income levels. Contributing factors include inadequate access to clean water, sanitation, and hygiene for both humans and animals; poor infection prevention and control in homes, healthcare facilities, and farms; limited availability of quality, affordable vaccines, diagnostics, and medicines; lack of awareness and knowledge; and insufficient enforcement of relevant legislation [8]. Populations living in resource-limited settings and vulnerable groups bear a disproportionate burden from both the drivers and consequences of antimicrobial resistance (AMR) [8].
Methicillin-resistant Staphylococcus aureus (MRSA) is a major nosocomial and community-acquired pathogen worldwide, associated with severe infections and high morbidity [9]. Its resistance to methicillin and related antibiotics severely limits treatment options, driving the search for novel antimicrobials. MRSA causes a spectrum of infections, ranging from mild skin and soft tissue infections, characterized by painful, swollen, erythematous lesions that often resemble boils or abscesses, to invasive diseases such as pneumonia, osteomyelitis, septic arthritis, endocarditis, bacteremia, and sepsis [10]. These infections can present with fever, chills, fatigue, and altered mental status [10]. MRSA’s multidrug resistance complicates treatment, leading to prolonged hospitalizations, increased healthcare costs, and higher morbidity and mortality [1]. Transmission occurs primarily through direct contact with colonized or infected individuals or exposure to contaminated surfaces and medical equipment. MRSA infections are categorized as healthcare-associated (HA-MRSA), which is linked to invasive procedures and multidrug resistance, or community-associated (CA-MRSA), which predominantly causes skin infections and generally shows resistance only to β-lactam antibiotics [9,11]. Both forms can persist asymptomatically for extended periods, posing ongoing challenges to infection control. Research into natural product antimicrobials, particularly plant-derived compounds, offers promising alternatives to combat MRSA’s resistance profile [12].
Numerous studies have demonstrated the antimicrobial and antioxidant properties of various M. cajuputi extracts [4,6,13-15]. Antibacterial assays reveal potent activity against Gram-positive but not Gram-negative pathogens, notably Staphylococcus aureus, Staphylococcus epidermidis, and Bacillus cereus [4]. Most research has focused on leaf extracts or essential oils, with limited detailed investigation of the flower extracts. Methanolic extracts of M. cajuputi leaves and flowers exhibit significant antioxidant activity, primarily due to high phenolic and flavonoid contents identified via LC/MS and GC/MS analyses [4,13]. These extracts efficiently scavenge free radicals and demonstrate strong ferric reducing antioxidant power. However, the activity of M. cajuputi flower extract against multidrug-resistant bacteria such as MRSA remains underexplored. Therefore, this study addresses these knowledge gaps by evaluating the antioxidant and antibacterial activities of M. cajuputi flower extract, with a focus on MRSA, to provide insights into its potential as a novel bioactive compound for therapeutic use. These findings offer a scientific basis for developing M. cajuputi flower constituents as alternative antibacterial therapies.
Extraction of Melaleuca cajuputi flowers
A voucher specimen (herbarium number: A 17937, BCU) was deposited at the Department of Botany, Faculty of Science, Chulalongkorn University, Bangkok, Thailand. The extraction method was slightly modified from the previous study [16]. Fresh flowers of M. cajuputi were collected from the Sirindhorn Peat Swamp Forest, Narathiwat Province, Thailand. The samples were thoroughly washed and dehydrated in a hot air oven at 40 °C for 48 h to ensure complete removal of moisture. The dried flowers were then pulverized into a fine powder using a laboratory blender. A total of 100 g of powdered flowers was macerated in 400 mL of ethanol (1:4 w/v) and shaken at 150 rpm for 48 h at room temperature. The filtrate was obtained by passing the extract through a muslin cloth, followed by Whatman No. 1 filter paper. The combined filtrates were concentrated by evaporating ethanol using a rotary evaporator and subsequently dried with a freeze dryer. The dried extract was kept in amber-colored bottles at 4 °C until further analysis.
Phytochemical constituent analysis
Total phenolic content analysis
The total phenolic content of the M. cajuputi flower extract was measured using the Folin-Ciocalteu method as previously described [17]. Briefly, the extract stock solution was diluted to 1 mg/mL with distilled water. For the assay, 20 µL of the diluted extract, 50 µL of distilled water, and 20 µL of Folin-Ciocalteu reagent were mixed in a 96-well microplate. After standing for 6 min, 100 μL of 7% Na₂CO₃ was added to each well. The mixture was incubated at room temperature for 90 min before measuring absorbance at 760 nm with a microplate reader. Gallic acid was used as the standard for calibration (6.25 - 300 mg/mL, R² = 0.999). Results are expressed as milligrams of gallic acid equivalents (GAE) per gram of dry weight. All assays were performed in triplicate.
Total flavonoid content analysis
The total flavonoid content was quantified using the aluminium chloride (AlCl3) colorimetric method [17]. The stock extract solution was diluted to 1 mg/mL with distilled water. Twenty-five µL of the extract were mixed with 125 µL of distilled water and 10 µL of 5% NaNO₂ solution in a 96-well microplate, and left for 6 min at room temperature. The mixture was added with 15 μL of 10% AlCl3 and incubated in the dark for 5 min. Then, 50 µL of 1 M NaOH was added to each well, and the absorbance was measured at 510 nm using a microplate reader. Calibration curves were constructed using epicatechin and quercetin standards (6.25 - 300 g/mL, R² = 0.999). The results were expressed as milligrams of catechin and quercetin equivalents per gram of dry sample weight.
Total tannin content analysis
The total tannin concentration was determined according to the method described by Haile and Kang [18]. Standard solutions of tannic acid (6.25 - 400 µg/mL) and extract solutions (1 mg/mL) were freshly prepared in distilled water. For the assay, 20 µL aliquots of the solutions were mixed with 50 µL of distilled water and 10 µL of Folin-Ciocalteu reagent in each well of a 96-well microplate. After allowing the mixture to react for 6 min at room temperature, 100 µL of 35% Na2CO3 solution was added. The reaction mixture was then incubated at room temperature for 30 min, and the absorbance was measured at 700 nm using a microplate reader. The total tannin concentration was expressed as milligrams of tannic acid equivalent (TAE) per gram of dry weight sample.
LC/MS analysis
Metabolite profiling of the extract was performed at the Central Equipment Unit, Prince of Songkla University, Thailand. The analysis was conducted using a Liquid Chromatograph-Quadrupole Time-of-Flight Mass Spectrometer (LC-QTOF MS), 1290 Infinity II LC-6545 Quadrupole-TOF, Agilent Technologies USA, in both positive and negative electrospray ionization modes. Chromatographic separation was performed on a Zorbax Eclipse Plus C18 Rapid Resolution HD column (100×2.1 mm2, 1.8 µm) maintained at 30 °C with a total run time of 35 min. The mobile phase consisted of water/acetonitrile (95:5%) with 0.1% formic acid. The sample was prepared by dissolving in ethanol and filtering through a 0.22 µm nylon membrane; 2.0 µL was injected for each run. Data were acquired over an m/z range of 100 - 1,000. Compound identification was accomplished by comparing the obtained mass spectra to entries in an online reference database using LC-MS data processing software.
Antioxidant activity assay
The DPPH radical scavenging assay
The antioxidant activity was determined using the DPPH radical scavenging assay [19]. Ascorbic acid solutions (0.39 - 25 µg/mL) and extract solutions (1.56 - 100 µg/mL) were prepared in methanol. One hundred microliters of each ascorbic acid or extract solution was dispensed into a 96-well microtiter plate in triplicate, followed by the addition of 100 µL of 0.2 mM DPPH solution. The mixtures were incubated in the dark at room temperature for 30 min, and the absorbance was measured at 517 nm. The antioxidant activity was expressed as percent inhibition, calculated using the formula: % inhibition = [(A sample/A control)] 100. The half-maximal inhibitory concentration (IC50) of the extract was calculated by plotting the percentage of DPPH radical scavenging activity against various extract concentrations, using ascorbic acid as the reference antioxidant standard. All experiments were performed in triplicate.
ABTS radical scavenging assay
The ABTS radical scavenging activity was determined according to the previous method [20] with minor modifications. The solution of ABTS (7 mM) and potassium persulfate (2.45 mM) was prepared. The ABTS + radical was produced by mixing the ABTS solution with potassium persulfate at a ratio of 1:1 and incubating in the dark for at least 16 h. The ABTS + radical solution was diluted with ethanol to obtain an absorbance of 1.4 0.02 at 734 nm. The ascorbic acid standard (0.39 - 25 µg/mL) and extract solution (1.56 - 100 µg/mL) were prepared in ethanol. Then, the sample or standard solutions (100 µL of each) were added to 10 mL of the ABTS + solution and thoroughly mixed in a 96-well microtiter plate, and incubated for 10 min. The plate was measured for the absorbance at 734 nm. The IC50 of the extract was calculated by plotting the percentage of ABTS radical scavenging activity against various extract concentrations, using ascorbic acid as the reference antioxidant standard. All experiments were performed in triplicate.
Antibacterial activity of the extract
Bacterial strains
Five reference bacterial strains used in this study, including Methicillin-Resistant Staphylococcus aureus (MRSA) DMST 20652, Staphylococcus aureus ATCC 6538, Bacillus cereus BCC 6386, Enterococcus faecalis TISTR 1485, and Pseudomonas aeruginosa ATCC 27853, were kindly provided by the Department of Microbiology, Faculty of Science, Kasetsart University, Bangkok, Thailand.
Agar well diffusion assay
The antibacterial activity of the crude extract was evaluated using a slightly modified agar well diffusion method [21]. Tryptic soy agar (TSA) plates containing 1.5×108 CFU/mL of each bacterial suspension were prepared. The crude extract was dissolved in DMSO to obtain a final concentration of 170 mg/mL. A 100 μL aliquot of each extract was loaded into a 7 mm diameter well cut into the agar using a sterile Cork borer, and maintained at room temperature for 5 h to permit adequate diffusion of the extract. The plates were incubated aerobically at 37 °C for 24 - 48 h. Zones of inhibition were measured to determine antibacterial activity. All assays were performed in triplicate.
Determination of minimal inhibitory concentration (MIC) and minimal bactericidal concentration (MBC)
MIC and MBC values of the extract were determined using the broth microdilution method in a 96-well plate [22,23]. Each well (2 - 11) received 100 L of TSB broth, and 200 L of TSB was added to well 12. Then, 200 L of the extract solution was added to well 1 and serially diluted twofold across the plates to achieve final concentrations of 1/2, 1/4, 1/8, 1/16, 1/32, 1/64, 1/128, 1/256, and 1/512 (Raw 10). Subsequently, 100 L of bacterial inoculum (1×106 CFU/mL) was transferred to each well. Wells 11 and 12 served as the positive and negative controls. The microplates were then incubated at 37 °C for 24 h. The lowest concentration of crude extract that inhibited bacterial growth and did not show turbidity is reported as the MIC. To determine the MBC, 100 L aliquots from the 4 clearest wells were spread onto TSA plates and incubated at 37 °C for 48 h. The concentration that produced ≥ 99.9% reduction in viable cells (no growth or fewer than 15 colonies) was recorded as the MBC [24]. Each assay was performed in triplicate.
Time-kill assay in the presence of the extract against the tested bacteria
This experiment was conducted following a slightly modified method from a previous study [25]. Five mL of each bacterial suspension in tryptic soy broth (TSB) containing 1.5×10⁶ CFU/mL were incubated with the crude extract at concentrations equivalent to the MIC of 0.5×, 1×, 2×, and 4×, and shaken at 150 rpm at 37 °C, and viable bacterial counts (CFU/mL) were determined every 2 h until 24 h. Tubes without the extract were incubated under identical conditions. All experiments were performed in triplicate.
Scanning electron microscopy (SEM) of treated MRSA
The effect of the crude extracts on MRSA cells was examined using scanning electron microscopy (SEM), following a modified protocol based on Santajit et al. [26]. MRSA cells (1×10⁶ CFU/mL) were incubated in TSB containing the crude extract at its MIC and MBC for 6 h. Cells treated with 1% DMSO served as the negative control. After incubation, the bacterial suspensions were centrifuged at 3,500×g for 3 min, transferred onto sterile glass coverslips (1.0×1.0 cm2), and fixed with 2.5% glutaraldehyde (Sigma-Aldrich, USA) for 2 h. All SEM preparation and imaging procedures were performed according to the manufacturer’s instructions by the staff at the Scientific Equipment Center, Kasetsart University, Thailand.
Statistical analysis
All experiments were conducted in triplicate, and the data are presented as the mean ± standard deviation (SD). Statistical analyses were carried out using GraphPad Prism version 10.0 (GraphPad Software, La Jolla, CA, USA). Statistical significance was considered as p < 0.05.
Total phenolic content, total flavonoid content, and total tannin content
The M. cajuputi flower ethanol extract exhibited a high total phenolic content (TPC) of 327.11 ± 3.90 mg gallic acid equivalents (GAE)/g, alongside substantial total flavonoid content (TFC) quantified as 28.88 ± 1.32 mg catechin equivalents (CE)/g and 200.25 ± 1.97 mg quercetin equivalents (QE)/g and total tannin content (TTC) of 350.09 ± 6.89 mg tannic acid equivalent (TA)/g. These phytochemical profiles suggest that the flower extract contains abundant phenolic, flavonoid, and tannin compounds, which are well-established contributors to antioxidant and antimicrobial activities in plant-derived extracts [4]. Phenolic compounds in plant extracts are well-recognized antioxidants, exerting their effects through radical scavenging and metal ion chelation, thereby protecting biomolecules from oxidative damage [4]. The high TPC value indicates a significant presence of these bioactive molecules, which likely play a major role in the observed radical scavenging and antimicrobial efficacy of the extract [4]. Flavonoids, quantified here in terms of catechin and quercetin equivalents, also have established pharmacological roles, including anti-inflammatory, antimicrobial, and antioxidative effects, which occur as diverse subclasses such as flavonols and flavones within M. cajuputi [13,27]. Similarly, the quantified flavonoid content, represented by both catechin and quercetin equivalents, reflects a diverse flavonoid composition, including flavonols and flavones that contribute to various pharmacological effects [13,27]. Comparisons with previous studies revealed that M. cajuputi flower extracts typically exhibit variable but generally high phenolic content, consistent with their strong antioxidant potential [4,13]. Variation in TPC and TFC values among studies can be attributed to differences in extraction solvents, geographical location, harvest time, and plant part used [28]. Notably, the use of ethanol as an extraction solvent in this study effectively recovered polar phenolic compounds, aligning with reports that polar solvents enhance the extraction of antioxidant phytochemicals [29,30]. The presence of these phenolics and flavonoids strengthens the therapeutic prospects of M. cajuputi flower extract as a natural source of antioxidants and antibacterials, supporting its application in food, pharmaceutical, and cosmetic industries.
LC/MS analysis of the flower from M. cajuputi
The phytochemical profile of the M. cajuputi flower extract was determined by LC/MS analysis. The LC/MS chromatogram is presented in Figure 1, and the molecular formulas, along with the relative abundances (expressed as peak area %) of the identified compounds, are summarized in Table 1. Analysis in negative ionization mode revealed that galloyl derivatives were the predominant compounds, with tannins constituting the major chemical group. The compound with the highest relative content was 1,2,3,5-Tetra-O-galloylhamamelofuranose (17.72%), followed by other galloylated tannins such as 1,2'-Di-O-galloylhamamelofuranose (9.57%) and 3-O-Galloylhamamelitannin (8.28%). These polyphenolic compounds are known for their potent antioxidant activities, protein-binding properties, and antimicrobial effects [14]. Catechols and simple phenolics, including gallic acid and 4-glucogallic acid, were present at approximately 4% each. Both are simple phenolic acids with effective radical-scavenging and anti-inflammatory activities [4,27,31]. Flavonoids and glycosides, including Quercetin 3-galactoside (8.89%), Kaempferol-7-o-glucoside (8.37%), and Quercetin (4.55%), highlight a significant flavonoid presence. Flavonoid glycosides are generally more bioavailable and contribute to antioxidant, anti-inflammatory, and antimicrobial activities [32]. The abundance of galloyl derivatives and flavonoids suggests a high capacity for neutralizing free radicals and reducing oxidative stress, properties that benefit cellular protection and potentially lower the risk of chronic diseases [4]. Tannins and flavonoids are also well-documented for their antimicrobial, antifungal, and antiviral activities, exerted through mechanisms such as membrane destabilization, enzyme inhibition, and interaction with microbial proteins [33,34]. The combination and diversity of these compounds suggest strong potential for the extract in health-related applications, including anti-inflammatory, anticancer, and antimicrobial uses.
Meanwhile, analysis conducted in positive ionization mode revealed 3 major phytochemical groups in the extract, which consisted of flavonoid glycosides (quercetagetin 3-rhamnoside), phenolic compounds (eugenitol), and triterpenoids (2-(2-methylbutanoyl)-9-(3-methyl-2E-pentenoyl)-2b,9a-dihydroxy-4Z,10(14)-oplopadien-3-one). Extensive studies corroborate these physiological functions and nutritional significance. Quercetin derivatives, including rhamnosides, are prominent flavonoid glycosides that offer substantial antioxidant [35], anti-inflammatory [36], and antimicrobial properties [37], as well as cellular protection against oxidative stress. Eugenitol and related phenolic compounds are noted for their potent antioxidant [38] and antimicrobial activities [39], and possibly exert anti-inflammatory and cytoprotective effects. Triterpenoids display a range of biological activities, with evidence for anti-inflammatory [40], antibacterial [41], immunomodulatory [42], and hepatoprotective effects [43], which enhances the pharmacological value of triterpenoid-rich plant extracts. This systematic phytochemical classification provides a framework for understanding the extract’s bioactive profile and its potential pharmacological effects.
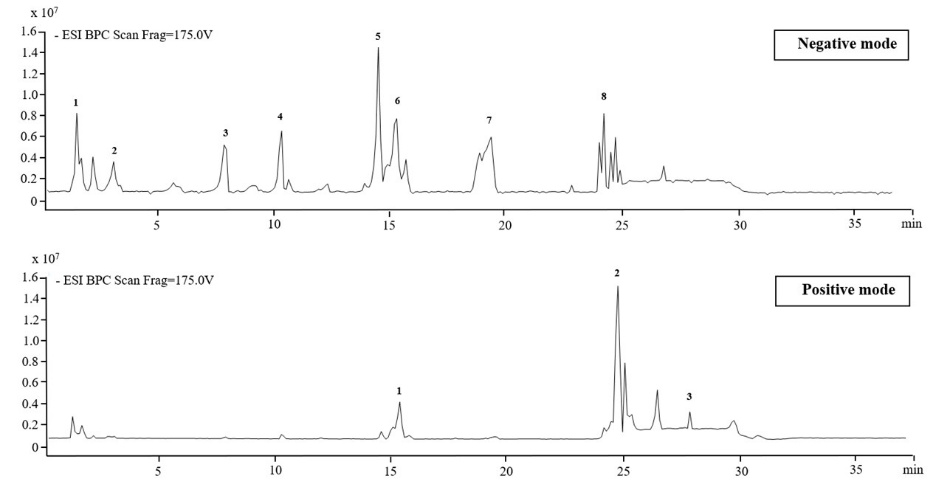
Figure 1 The LC/MS Chromatogram of M. cajuputi flower ethanolic extract.
Table 1 Phytochemical constituents in the crude ethanol extract of M. cajuputi flowers identified by LC/MS.
No |
Compound |
Molecular Formula |
Retention time (min) |
Fragment ions (m/z) |
Score (DB) |
Diff (DB, ppm) |
Relative content (%) |
Negative mode |
|
|
|
|
|
|
|
1. |
4-Glucogallic acid |
C13 H16 O10 |
1.464 |
99.54 |
0.38 |
4.74 |
|
2. |
Gallic acid |
C7 H6 O5 |
3.056 |
99.95 |
−0.26 |
4.04 |
|
3. |
1,2'-Di-O-galloylhamamelofuranose |
C20 H20 O14 |
7.906 |
483.07 |
99.28 |
−0.07 |
9.57 |
4. |
3-O-Galloylhamamelitannin |
C27 H24 O18 |
10.352 |
635.09 |
99.5 |
0.13 |
8.28 |
5. |
1,2',3,5-Tetra-O-galloylhamamelofuranose |
C34 H28 O22 |
14.587 |
787.10 |
99.65 |
0.45 |
17.72 |
6. |
Quercetin 3-galactoside |
C21 H20 O12 |
15.351 |
99.09 |
−0.03 |
8.89 |
|
7. |
Kaempferol-7-o-glucoside |
C21 H20 O11 |
19.468 |
98.84 |
0.02 |
8.37 |
|
8. |
Quercetin |
C15 H10 O7 |
24.405 |
99.46 |
−0.26 |
4.55 |
|
Positive mode |
|
|
|
|
|
|
|
1. |
Quercetagetin 3-rhamnoside |
C21 H20 O12 |
15.385 |
487.08 |
98.49 |
−0.48 |
8.06 |
2. |
Eugenitol |
C11 H10 O4 |
24.806 |
207.07 |
98.26 |
−1.98 |
44.96 |
3. |
2-(2-Methylbutanoyl)-9-(3-methyl-2E-pentenoyl)-2b,9a-dihydroxy-4Z,10(14)-oplopadien-3-one |
C26 H38 O5 |
27.923 |
453.26 |
98.81 |
−0.32 |
2.51 |
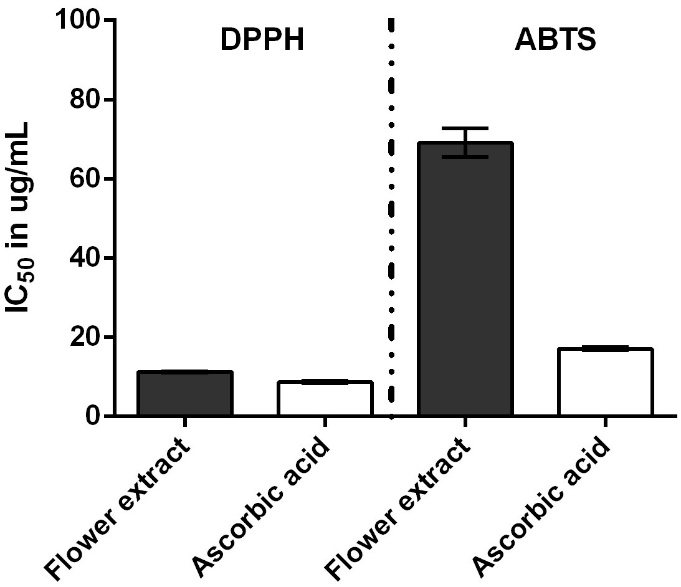
The antioxidant activity was evaluated using 2 complementary radical scavenging assays, DPPH and ABTS, with results expressed as IC50 values (μg/mL). Ascorbic acid, a well-established antioxidant standard, exhibited IC50 values of 8.67 ± 0.41 μg/mL (DPPH) and 17.08 ± 0.38 μg/mL (ABTS), confirming the validity of the assay. The ethanolic extract of M. cajuputi flowers demonstrated superior antioxidant capacity, with notably low IC50 values of 11.28 ± 0.30 μg/mL (DPPH) and 61.90 ± 14.84 μg/mL (ABTS) (Figure 2). This indicates a strong radical scavenging ability comparable to that of ascorbic acid, particularly in the DPPH assay. The extract’s potent antioxidant activity, as evidenced by these low IC50 values in both DPPH and ABTS assays, is closely associated with its high phenolic (327.11 ± 3.90 mg GAE/g) and flavonoid contents (28.88 ± 1.32 mg CE/g and 200.25 ± 1.97 mg QE/g). Phenolic and flavonoid compounds are well-documented for their electron-donating capabilities, which enable the neutralization of free radicals by stabilizing unpaired electrons, thereby conferring antioxidant effects [44]. These compounds may also act synergistically, enhancing the overall scavenging capacity of the extract. Comparatively, the antioxidant activity of the M. cajuputi flower extract in this study is consistent with or exceeds values reported for related Melaleuca species and other medicinal plants. For example, Khongsai et al. [14] reported total phenolic contents in M. cajuputi extracts correlating with significant free radical scavenging activity. The greater activity observed in the flower extract compared to the leaf extract is a common finding in plant studies [45], as flowers tend to accumulate higher concentrations of bioactive flavonoids and phenolics due to their ecological role in attracting pollinators and protecting reproductive tissues [4].
The observed difference in antioxidant potential between the DPPH and ABTS assays aligns with their distinct radical species and reaction kinetics. DPPH primarily measures hydrogen donation to a stable nitrogen-centered radical, favoring lipophilic antioxidants, while ABTS assesses scavenging of a water-soluble radical cation and is responsive to both hydrophilic and lipophilic compounds [46]. This feature explains the stronger activity of the flower extract in the DPPH assay relative to ABTS, likely reflecting its phenolic profile. Variation in phytochemical content may also result from botanical origin, processing, and storage conditions. Overall, the high phytochemical content and robust radical scavenging activity of M. cajuputi flower extract support its potential as a valuable natural source of antioxidants. Further investigation into its application in nutraceutical, pharmaceutical, or cosmetic formulations aimed at mitigating oxidative stress-related diseases is warranted [47].
Agar well diffusion assay
The crude ethanolic extract of M. cajuputi flower at 170 mg/well exhibited significant antibacterial activity against all tested bacterial strains, as measured by the agar well diffusion assay (Table 2). MRSA DMST 20652 showed the highest susceptibility, while P. aeruginosa ATCC 27853, E. faecalis TISTR1485, and S. aureus ATCC6538 demonstrated moderate susceptibility, and B. cereus BCC6386 was the least susceptible. These results indicate that the crude extract possesses broad-spectrum antibacterial properties, particularly strong against MRSA, suggesting its potential as a natural antimicrobial agent. The highest susceptibility of MRSA DMST 20652 is with previous reports of antibacterial activity of M. cajuputi extracts, attributed to their bioactive flavonoids, alkaloids, and phenolics [4]. The moderate susceptibility observed in other strains aligns with findings that M. cajuputi extracts are generally more effective against Gram-positive bacteria, with some activity against Gram-negative species depending on extract concentration and method. The comparatively lower susceptibility of B. cereus BCC 6386 may reflect differences in cell wall structure and resistance mechanisms to the extract’s phytochemicals [13,27].
Table 2 Antibacterial activity of M. cajuputi flower ethanolic extract against test bacteria determined by agar well diffusion assay.
Bacterial strains |
Inhibition zone (mm SD) |
MRSA DMST 20652 |
38.33 0.58 |
P. aeruginosa ATCC 27853 |
30.67 0.58 |
E. faecalis TISTR 1485 |
30.00 1.00 |
B. cereus BCC 6386 |
25.67 0.58 |
S. aureus ATCC 6538 |
28.50 0.71 |
Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC)
MIC and MBC values of the M. cajuputi flower extract against 5 bacterial strains are shown in Table 3. MIC values ranged from 0.33 to 125 mg/mL, while MBC values ranged from 5.31 to 250 mg/mL. The extract exhibited the most potent antibacterial activity against MRSA DMST 20652, with MIC and MBC values of 0.33 and 5.31 mg/mL, respectively. The MBC/MIC ratio, a well-established parameter for distinguishing bactericidal from bacteriostatic effects, was calculated accordingly. Ratios ≤ 4 indicate bactericidal activity, whereas ratios > 4 suggest bacteriostatic effects [48]. For MRSA DMST 20652, the MBC/MIC ratio was 16, indicating a bacteriostatic effect. In contrast, E. faecalis TISTR1485 and P. aeruginosa ATCC 27853 both showed a ratio of 2, consistent with strong bactericidal activity. Similarly, B. cereus BCC 6386 had a ratio of 2, indicating bactericidal activity but requiring higher concentrations than the other tested strains. S. aureus ATCC 6538 demonstrated a ratio of 1, reflecting equivalent inhibitory and bactericidal concentrations, albeit at higher extract doses.
Table 3 The minimum inhibitory concentration (MIC) and the minimum bactericidal concentration (MBC) of the M. cajuputi flower extract against tested bacterial strains.
Bacterial test |
The M. cajuputi flower extract (mg/mL) |
||
MIC |
MBC |
MBC/MIC ratio |
|
MRSA DMST 20652 |
0.33 |
5.31 |
16 |
E. faecalis TISTR 1485 |
2.66 |
5.31 |
2 |
P. aeruginosa ATCC 27853 |
21.25 |
42.50 |
2 |
85 |
85 |
1 |
|
B. cereus BCC 6386 |
125 |
250 |
2 |
Values are presented as mean ± SD from 2 independent experiments performed in triplicate.
The antibacterial mechanisms underlying the observed effects likely involve multiple biochemical disruptions. Given the extract’s efficacy against both Gram-positive and Gram-negative bacteria, the potential modes of action may include perturbation of the bacterial cytoplasmic membrane, inhibition of peptidoglycan synthesis, and interference with essential intracellular processes [49,50]. The antibacterial mechanisms underlying the observed effects likely involve multiple biochemical disruptions. As illustrated in Figure 3 (agar well diffusion assay plate), the ethanol extract of M. cajuputi (White Samet) flower generated the largest and clearest zone of inhibition compared to hexane extracts from flower and leaf, as well as the ethanol leaf extract, visually confirming its superior anti-MRSA activity during initial screening. This image highlights the extract's effective diffusion and bactericidal potential against the test strains.
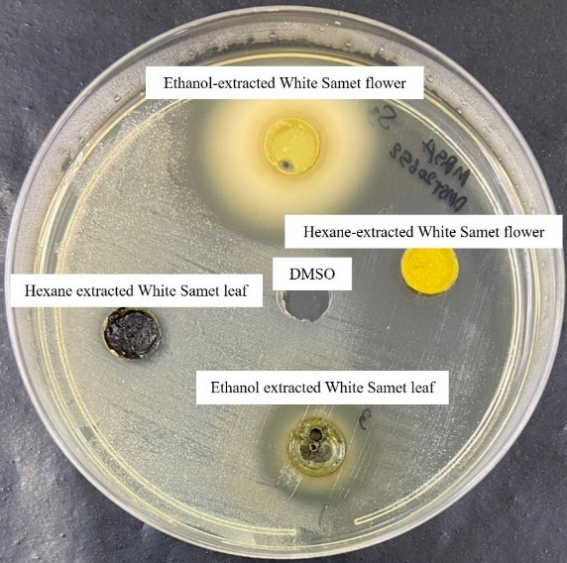
Figure 3 Representative agar well diffusion assay results. Tryptic soy agar plate inoculated with Methicillin-Resistant Staphylococcus aureus (MRSA) DMST 20652, showing zones of inhibition around wells containing M. cajuputi flower and leaf extracts (170 mg/mL). The control well (DMSO) showed no inhibition.
The antimicrobial efficacy of the flower extract can principally be attributed to its high content of total phenolic compounds and flavonoids. Phenolics destabilize bacterial membranes and inhibit enzymes [51], whereas flavonoids chelate metal ions and generate bactericidal oxidative stress [49]. For instance, phenolic acids such as gallic acid and ferulic acid permeabilize membranes, disrupt ion pumps, and acidify the cytoplasm, leading to protein denaturation and bacterial death [52]. These effects are especially pronounced in Gram-positive bacteria due to their lack of an outer membrane, whereas Gram-negative bacteria exhibit greater resistance because of their complex membrane structures [52]. Tannins are known to kill or inhibit MRSA through multi-targeted disruption of protein synthesis machinery and bacterial structure at the molecular level [53]. Flavonoids exert antibacterial effects by chelating metal ions essential for bacterial metabolism and by generating bactericidal oxidative stress [54]. Additionally, they can disrupt membrane permeability [55], inhibit DNA gyrase activity (which is important for DNA replication in bacteria), and affect enzyme functions through hydroxylation and other chemical modifications [56]. These actions result in bacterial leakage, defective cell division, and ultimately, cell death. Specific structural features of flavonoids, such as hydroxyl groups and prenylation, enhance these antibacterial activities [57]. These bioactivities corroborate previous empirical data demonstrating the inhibitory effects of M. cajuputi ethanol extracts against pathogens, including MRSA, S. aureus, and B. cereus via agar diffusion and MIC/MBC assays [4,13,27].
The time-kill assay revealed a distinct dose- and time-dependent antibacterial activity of M. cajuputi flower extract against B. cereus BCC6386, S. aureus ATCC6538, P. aeruginosa ATCC 27853, E. faecalis TISTR1485, and MRSA DMST 20652 (Figure 4). In all experiments, the untreated control showed sustained bacterial proliferation over 24 h, as demonstrated by consistently elevated log10 CFU/mL values. Upon treatment, bacterial viability declined in proportion to extract concentration and exposure duration. Notably, at concentrations of 1× MIC and above (1×, 2× and 4×), a rapid reduction in CFU counts was observed within the first 4-8 h, with near-complete bacterial eradication achieved at 2× and 4× MIC by 8 - 24 h, depending on the tested strains. Exposure to Sub-MIC (0.5× MIC) resulted in limited growth inhibition, reflected by lower but persistent CFU counts compared to controls. The extract exhibited a bacteriostatic rather than bactericidal mode of action on MRSA DMST 20652 across all evaluated concentrations, as evidenced by the stable CFU counts and the absence of further decline. These findings demonstrate that M. cajuputi flower extract exhibits strong, broad-spectrum bactericidal activity in both a concentration- and time-dependent manner against Gram-positive and Gram-negative bacteria.
The findings from the time-kill assay in this study are consistent with previous reports that demonstrate a dose- and time-dependent bactericidal mode of action of plant extracts, particularly those derived from Melaleuca species. The rapid decline in viable cell counts at concentrations equal to or higher than the MIC, especially for Gram-positive bacteria, aligns with the reports by Al-Abd et al. [4]; Isah et al. [13]. In which M. cajuputi flower extract exhibited significant antibacterial activity, with complete eradication achieved at higher concentrations and extended exposure times. In the present study, the extract was less effective against Gram-negative bacteria, including P. aeruginosa ATCC 27853, which required higher concentrations for comparable bactericidal action. Time-kill assays showed faster bactericidal killing (≥ 3-log10 CFU/mL reduction within 6 - 8 h at ≥ 2× MIC) for Gram-positive bacteria (e.g., MRSA at 0.66 mg/mL, E. faecalis TISTR 1485 at 5.32 mg/mL), while Gram-negative P. aeruginosa required higher concentrations (≥ 42.5 mg/mL) and longer exposure (8 - 14 h) for equivalent killing. This differential efficacy is attributed to Gram-positive bacteria's single peptidoglycan layer (easily disrupted by phenolics/flavonoids) versus Gram-negative outer membrane lipopolysaccharide barrier that limits extract penetration, a common pattern for plant antimicrobials [13]. This differential susceptibility is a well-established phenomenon noted in studies of various natural product antimicrobials [58]. Overall, these time-kill kinetics reinforce the potential of M. cajuputi flower extract as a broad-spectrum antibacterial agent, confirming both the rapid and concentration-dependent lethality observed for botanical extracts in the literature. Such findings support ongoing efforts to develop plant-derived antimicrobials as alternative or adjunct solutions to combat the growing challenge of drug-resistant pathogens.
MRSA remains a predominant nosocomial pathogen and poses significant clinical challenges due to its multidrug resistance [59]. Accordingly, MRSA was prioritized for subsequent mechanistic analyses, which incorporated time-kill kinetic assays and scanning electron microscopy (SEM) to visualize ultrastructural bacterial damage. Morphological alternations in MRSA DMST 20652 cells following exposure to M. cajuputi flower ethanolic extract at its MIC (0.33 mg/mL) and MBC (5.31 mg/mL) were visualized using SEM (Figure 5). Untreated MRSA cells exhibited smooth, spherical, and well-defined coccoid morphology with intact cell surfaces and no evidence of cell damage or lysis (Figure 5(A)). In contrast, cells exposed to 1× MIC extract exhibited early morphological disruption, characterized by a less uniform, rougher surface and mild deformation, while generally retaining their overall shape (Figure 5(B)). Notably, exposure to the 1× MBC extract induced severe structural abnormalities as cells appeared shrunken, extensively deformed, and exhibited major loss of surface integrity, indicative of substantial membrane damage and cellular collapse (Figure 5(C)). These findings highlight a clear concentration-dependent disruptive effect of the M. cajuputi flower extract on MRSA cell morphology, with higher concentrations producing more significant structural changes consistent with bactericidal activity.
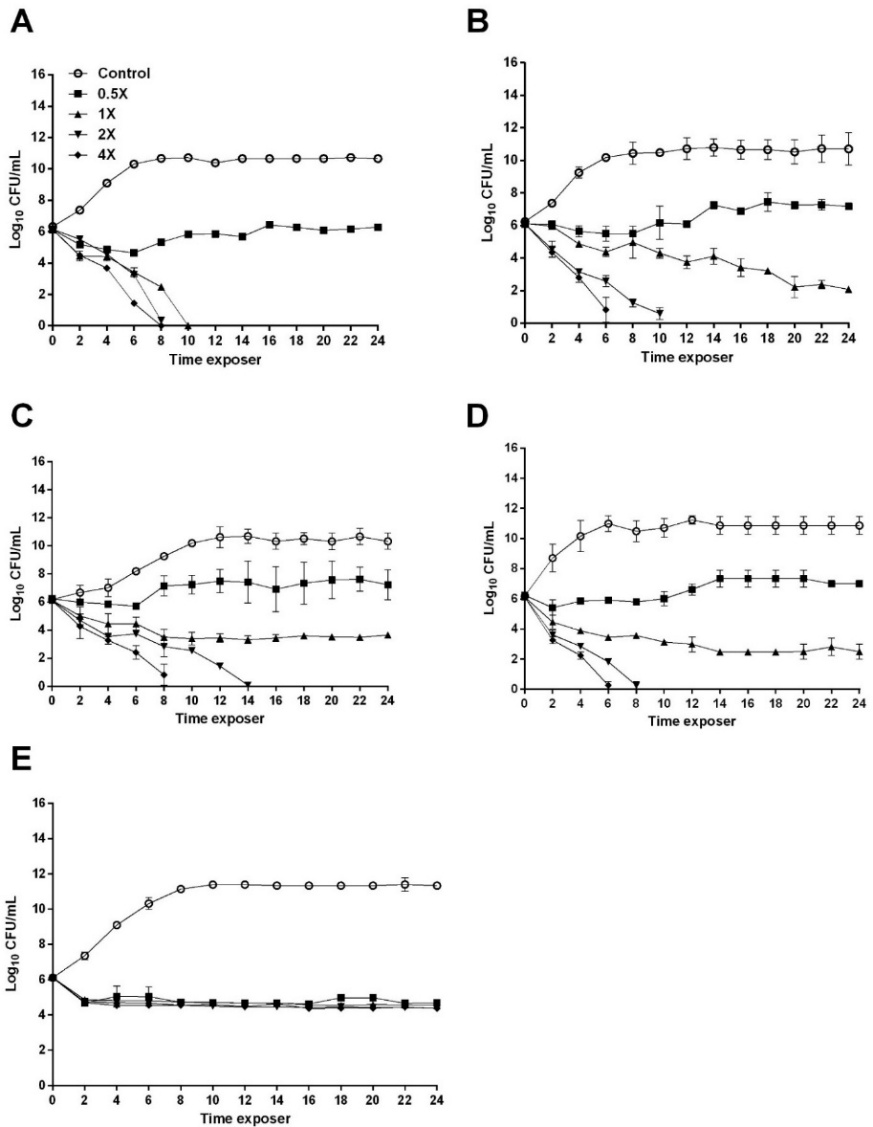
Figure 4 Time-killing kinetics of the M. cajuputi flower extract against B. cereus BCC 6386 (A), S. aureus ATCC 6538 (B), P. aeruginosa ATCC 27853 (C), E. faecalis TISTR 1485 (D), and MRSA DMST 20652 (E). Error bars represent reproducibility across 3 independent experiments.
These changes indicate compromised cell envelope integrity and cellular leakage of cellular contents. Such ultrastructural alterations have been reported in MRSA and other bacteria treated with antimicrobial agents that disrupt the cell wall and membrane [60]. Previous studies using scanning electron microscopy have observed similar phenomena, such as cell wall disintegration, shrinkage, and surface irregularities, following exposure to antibacterial compounds, including essential oils and silver nanoparticles [61]. These disruptions can increase permeability, leading to cell death [62]. The organic debris observed on the treated cells likely represents lysed cell components and extruded cytoplasmic material, further supporting the bactericidal effect seen in the time-kill assay (Figure 4). The similarity of morphological damage between MIC and MBC assay reinforces the extract's efficacy in compromising MRSA cell structures at both inhibitory and bactericidal concentrations. Collectively, these SEM findings provide visual confirmation of the extract’s disruptive effect on MRSA morphology and cell integrity, complementing the quantitative antibacterial data. This strengthens the potential of M. cajuputi flower extract as a natural antimicrobial agent against resistant bacterial strains. In future studies, we plan to determine the synergistic activity of the extract with conventional antibiotics, investigate the pure bioactive compounds of the M. cajuputi flower ethanol extract, and assess the cytotoxicity of these pure active compounds toward mammalian cells.
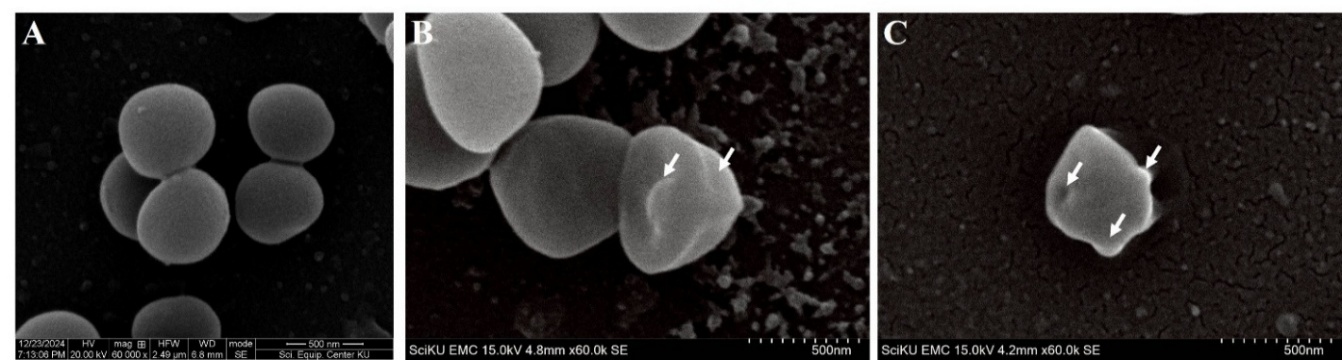
Figure 5 SEM images of Methicillin-Resistant Staphylococcus aureus DMST 20652 after incubation with M. cajuputi flower extract at 1× MIC and 1× MBC for 6 h. The initial bacterial cell density was 1×106 cells cm−3. Untreated cells served as the control (A), while bacterial cells were treated with ethanolic extract at 1× MIC (B) and 1× MBC (C). Arrows indicate disrupted cell structure.
This study has some limitations that merit consideration. First, evaluations were conducted in vitro only. While the M. cajuputi flower ethanol extract showed promising anti-MRSA activity (MIC 0.33 mg/mL), in vivo efficacy, bioavailability, and toxicity await assessment in animal models and pharmacokinetic studies. Second, although several solvents (e.g., hexane, water) were screened via agar well diffusion, the ethanol extract offered the highest yield and potency. Future fractionation could further isolate and optimize active compounds. Finally, mechanisms beyond SEM morphology (e.g., efflux pump inhibition, quorum sensing) were not examined. Targeted assays in subsequent work would elucidate the mode of action.
In conclusion, the ethanol extract of M. cajuputi flower exhibited strong antioxidant and broad-spectrum antibacterial activities, primarily attributed to its high phenolic, flavonoid, and tannin contents. The extract demonstrated significant bactericidal and bacteriostatic effects against multiple clinically relevant bacteria, including drug-resistant strains such as MRSA. Kinetic analyses confirmed a dose- and time-dependent reduction in bacterial viability, with the greatest efficacy observed at higher extract concentrations. Scanning electron microscopy revealed substantial morphological disruptions in MRSA cells, characterized by compromised envelope integrity and cell shrinkage, providing visual evidence of membrane-targeted antibacterial mechanisms. These findings indicate that M. cajuputi flower extract is a promising natural antibacterial candidate for combating multidrug-resistant pathogens. They warrant further investigation into their therapeutic applications and potential synergistic effects with existing antibiotics.
Acknowledgments
This work was financially supported by the Faculty of Medicine, Princess of Naradhiwas University, Thailand. Additional funding for Somruedee Yorsin was provided by the Thailand Science Research and Innovation Fund (Fundamental Fund), Princess of Naradhiwas University, Thailand.
Declaration of Generative AI in Scientific Writing
The authors acknowledge the use of generative AI tools (e.g., Perplexity by OpenAI) and Grammarly for language editing and grammar correction during manuscript preparation. No artificial intelligence tools were used for content generation, study design, data analysis, or scientific interpretation. The authors have carefully reviewed and edited all text, and accept full responsibility for the integrity and accuracy of the work.
CRediT Author Statement
Tanes Sangsri: Conceptualization; Investigation; Writing - Original draft preparation; Writing - Reviewing and Editing; Funding acquisition. Somruedee Yorsin: Investigation; Writing - Original draft preparation; Writing - Reviewing and Editing; Funding acquisition. Patcharaporn Siwayaprahm: Investigation; Writing - Reviewing and Editing. Watchara Chongsa: Investigation; Writing - Original draft preparation.
References
A Parmanik, S Das, B Kar, A Bose, GR Dwivedi and MM Pandey. Current treatment strategies against multidrug-resistant bacteria: A review. Current Microbiology 2022; 79(12), 388.
OF Nwabor, S Leejae and SP Voravuthikunchai. Rhodomyrtone accumulates in bacterial cell wall and cell membrane and inhibits the synthesis of multiple cellular macromolecules in epidemic methicillin-resistant Staphylococcus aureus. Antibiotics 2021; 10(5), 543.
J Sharifi-Rad, B Salehi, EM Varoni, F Sharopov, Z Yousaf, SA Ayatollahi, F Kobarfard, M Sharifi-Rad, MH Afdjei, M Sharifi-Rad and M Iriti. Plants of the Melaleuca genus as antimicrobial agents: From farm to pharmacy. Phytotherapy Research 2017; 31(10), 1475-1494.
NM Al-Abd, ZM Nor, M Mansor, F Azhar, MS Hasan and M Kassim. Antioxidant, antibacterial activity, and phytochemical characterization of Melaleuca cajuputi extract. BMC Complementary and Alternative Medicine 2015; 15(1), 385.
AAM Noor. Melaleuca cajuputi Powell essential oil: A review of botanical, phytochemical and pharmacological properties. Borneo Journal of Resource Science and Technology 2023; 13(2), 1-12.
FMA Bar. Genus Melaleuca: A review on the phytochemistry and pharmacological activities of the non-volatile components. Records of Natural Products 2021; 15(4), 219-242.
CJL Murray. Global burden of bacterial antimicrobial resistance, 1990-2021: A systematic analysis with forecasts to 2050. The Lancet 2024; 404(10459), 1199-1226.
MH Nabi, FH Rupa, I Jahan, ABMN Hasan, F Naz, I Masud, MDH Hawlader and M Hossian. Water, sanitation, and hygiene (WASH) factors and the incidence of communicable diseases in Urban Bangladesh: Evidence from municipal areas. PLOS Neglected Tropical Diseases 2025; 19(7), e0013329.
N Hiermandi, C Foster, J Campbell, K Purnell, E Tocco, T Koy, K Nobleza, D Nguyen and L Marquez. Impact of methicillin-resistant Staphylococcus aureus surveillance and decolonization in the NICU: The Texas children's hospital experience. Antimicrobial Stewardship & Healthcare Epidemiology 2025; 5(1), e60.
NA Turner, BK Sharma-Kuinkel, SA Maskarinec, EM Eichenberger, PP Shah, M Carugati, TL Holland and VG Fowler. Methicillin-resistant Staphylococcus aureus: An overview of basic and clinical research. Nature Reviews Microbiology 2019; 17(4), 203-218.
A Selim, M Marzok, HS Gattan and AM Hereba. Prevalence of methicillin-resistant Staphylococcus aureus (MRSA) in Egyptian water buffaloes and risk factors for subclinical mastitis. Transboundary and Emerging Diseases 2025; 2025(1), 8862271.
SP Voravuthikunchai and L Kitpipit. Activity of medicinal plant extracts against hospital isolates of methicillin-resistant Staphylococcus aureus. Clinical Microbiology and Infection 2005; 11(6), 510-512.
M Isah, MD Sul’Ain, WWA Wahab, H Abdullah, S Jamil, N Syamira M Shabudin, AN Shuid and WRW Ishak. Chemical profiling and mechanistic insights into the antibacterial efficacy of Melaleuca cajuputi leaf extract. BMC Complementary Medicine and Therapies 2025; 25(1), 121.
S Khongsai, C Chalad and P Phumee. Phytochemical analysis, in vitro screening of antioxidant and antibacterial potential of cajuput (Melaleuca cajuputi) extract against pathogenic Vibrio spp. Tropical Journal of Natural Product Research 2024; 8(12), 9517-9523.
NZA Wahab, NS Aqilah Ja’Afar and SB Ismail. Evaluation of antibacterial activity of essential oils of Melaleuca cajuputi Powell. Journal of Pure & Applied Microbiology 2022; 16(1), 549-556.
S Nanasombat, K Yansodthee and I Jongjaited. Evaluation of antidiabetic, antioxidant and other phytochemical properties of Thai fruits, vegetables and some local food plants. Walailak Journal of Science and Technology 2018; 16(11), 851-866.
S Settharaksa, F Madaka, K Chakree and L Charoenchai. Total phenolic and flavonoid contents and antioxidant properties of Thai traditional herbal. International Journal of Pharmacy and Pharmaceutical Sciences 2014; 6(9), 564-566.
M Haile and WH Kang. Antioxidant activity, total polyphenol, flavonoid and tannin contents of fermented green coffee beans with selected yeasts. Fermentation 2019; 5(1), 29.
R Sawczuk, J Karpinska, D Filipowska, A Bajguz and M Hryniewicka. Evaluation of total phenols content, anti-DPPH activity and the content of selected antioxidants in the honeybee drone brood homogenate. Food Chemistry 2022; 368, 130745.
OM Noman, FA Nasr, AS Alqahtani, M Al-zharani, MAW Cordero, AA Alotaibi, A Bepari, S Alarifi and A Daoud. Comparative study of antioxidant and anticancer activities and HPTLC quantification of rutin in white radish (Raphanus sativus L.) leaves and root extracts grown in Saudi Arabia. Open Chemistry 2021; 19(1), 408-416.
S Arora, SA Saquib, YA Algarni, MA Kader, I Ahmad, MY Alshahrani, P Saluja, SM Baba, AM Abdulla and SS Bavabeedu. Synergistic effect of plant extracts on endodontic pathogens isolated from teeth with root canal treatment failure: An in vitro study. Antibiotics 2021; 10(5), 552.
M Bakht, SA Alizadeh, S Rahimi, RK Anari, M Rostamani, A Javadi, A Peymani, SMA Marashi and F Nikkhahi. Phenotype and genetic determination of resistance to common disinfectants among biofilm-producing and non-producing Pseudomonas aeruginosa strains from clinical specimens in Iran. BMC Microbiology 2022; 22, 114.
G Montrucchio, S Corcione, G Sales, A Curtoni, FG De Rosa and L Brazzi. Carbapenem-resistant Klebsiella pneumoniae in ICU-admitted COVID-19 patients: Keep an eye on the ball. Journal of Global Antimicrobial Resistance 2020; 23, 398-400.
K Kawamura-Sato, JI Wachino, T Kondo, H Ito and Y Arakawa. Reduction of disinfectant bactericidal activities in clinically isolated Acinetobacter species in the presence of organic material. Journal of Antimicrobial Chemotherapy 2008; 61(3), 568-576.
A Mordmuang, E Brouillette, SP Voravuthikunchai and F Malouin. Evaluation of a Rhodomyrtus tomentosa ethanolic extract for its therapeutic potential on Staphylococcus aureus infections using in vitro and in vivo models of mastitis. Veterinary Research 2019; 50(1), 49.
S Santajit, T Thavorasak, D Horpet, T Kong-Ngoen, U Permpoon, CY Kim, TG Nam and N Indrawattana. Phytochemical inhibition of quorum sensing and biofilm formation by Paederia foetida Linn. against multidrug-resistant Acinetobacter baumannii: An integrated in vitro and in silico investigation. Veterinary World 2025; 18(8), 2181.
M Isah, RA Rosdi, H Abdullah, MD Sul’Ain and WRW Ishak. Phytoconstituents and biological activities of Melaleuca cajuputi Powell: A scoping review. Journal of Applied Pharmaceutical Science 2023; 13(1), 010-023.
W Hou, W Zhang, G Chen and Y Luo. Optimization of extraction conditions for maximal phenolic, flavonoid and antioxidant activity from Melaleuca bracteata leaves using the response surface methodology. PLoS One 2016; 11(9), e0162139.
S Assaw, MIH Mohd Amir, TT Khaw, K Bakar, SA Mohd Radzi and NW Mazlan. Antibacterial and antioxidant activity of naphthofuranquinones from the twigs of tropical mangrove Avicennia officinalis. Natural Product Research 2020; 34(16), 2403-2406.
QD Do, AE Angkawijaya, PL Tran-Nguyen, LH Huynh, FE Soetaredjo, S Ismadji and YH Ju. Effect of extraction solvent on total phenol content, total flavonoid content, and antioxidant activity of Limnophila aromatica. Journal of Food and Drug Analysis 2014; 22(3), 296-302.
PH Tran, TTT Vu, TDT Phan, VM Nguyen, TNM Ngo, CVC Le and THD Ton. Chemical compositions and biological properties of the leaf essential oil of three Melaleuca species. World Academy of Sciences Journal 2024; 6(6), 67.
A Roy, A Khan, I Ahmad, S Alghamdi, BS Rajab, AO Babalghith, MY Alshahrani, S Islam and R Islam. Flavonoids a bioactive compound from medicinal plants and its therapeutic applications. BioMed Research International 2022; 2022(1), 5445291.
J Huang, M Zaynab, Y Sharif, J Khan, R Al-Yahyai, M Sadder, M Ali, SR Alarab and S Li. Tannins as antimicrobial agents: Understanding toxic effects on pathogens. Toxicon 2024; 247, 107812.
X Villanueva, L Zhen, JN Ares, T Vackier, H Lange, C Crestini and HP Steenackers. Effect of chemical modifications of tannins on their antimicrobial and antibiofilm effect against Gram-negative and Gram-positive bacteria. Frontiers in Microbiology 2023; 13, 987164.
H Han, B Xu, A Amin, H Li, X Yu, M Gong and L Zhang. Quercetin-3-O-α-L-rhamnopyranoside derived from the leaves of Lindera aggregata (Sims) Kosterm. evokes the autophagy-induced nuclear factor erythroid 2-related factor 2 antioxidant pathway in human umbilical vein endothelial cells. International Journal of Molecular Medicine 2019; 43(1), 461-474.
D Aggarwal, M Chaudhary, SK Mandotra, HS Tuli, R Chauhan, NC Joshi, D Kaur, L Dufossé and A Chauhan. Anti-inflammatory potential of quercetin: From chemistry and mechanistic insight to nanoformulations. Current Research in Pharmacology and Drug Discovery 2025; 8, 100217.
D Yang, T Wang, M Long and P Li. Quercetin: Its main pharmacological activity and potential application in clinical medicine. Oxidative Medicine and Cellular Longevity 2020; 2020, 8825387.
ROS Damasceno, JLS Pinheiro, LHM Rodrigues, RC Gomes, ABS Duarte, JJ Emidio, LRL Diniz and DP de Sousa. Anti-inflammatory and antioxidant activities of eugenol: An update. Pharmaceuticals 2024; 17(11), 1505.
MK Elbestawy, GM El-Sherbiny and SA Moghannem. Antibacterial, antibiofilm and anti-inflammatory activities of eugenol clove essential oil against resistant Helicobacter pylori. Molecules 2023; 28(6), 2448.
H Cheng, H Liu, X Qi, Y Fan, Z Yuan, Y Xu, Y Liu, K Guo and S Li. Anti-inflammatory and hepatoprotective triterpenoids from the traditional Mongolian medicine Gentianopsis barbata. Chinese Journal of Natural Medicines 2025; 23(9), 1111-1121.
JX Ma, CS Cai, JJ Liu, S Gao, GZ Zhao and XW He. In vitro antibacterial and antitumor activity of total triterpenoids from a medicinal mushroom Sanghuangporus sanghuang (Agaricomycetes) in liquid fermentation culture. International Journal of Medicinal Mushrooms 2021; 23(7), 27-39.
G Renda, İ Gökkaya and D Şöhretoğlu. Immunomodulatory properties of triterpenes. Phytochemistry Reviews 2022; 21(2), 537-563.
M Wang, S Jiang, N Hussain, S Zafar, Q Xie, F Huang, L Mao, B Li, Y Jian and W Wang. Anti-RAFLS triterpenoids and hepatoprotective lignans from the leaves of Tujia ethnomedicine Kadsura heteroclita (Xuetong). Frontiers in Chemistry 2022; 10, 878811.
N Mohammadi, ADS Lima, L Azevedo and D Granato. Bridging the gap in antioxidant activity of flavonoids: Correlating the oxidation of human plasma with chemical and cellular assays. Current Research in Food Science 2024; 8, 100714.
NW Albert, DJ Lafferty, SM Moss and KM Davies. Flavonoids-flowers, fruit, forage and the future. Journal of the Royal Society of New Zealand 2023; 53(3), 304-331.
J Rumpf, R Burger and M Schulze. Statistical evaluation of DPPH, ABTS, FRAP, and Folin-Ciocalteu assays to assess the antioxidant capacity of lignins. International Journal of Biological Macromolecules 2023; 233, 123470.
F Shahidi and J Yeo. Bioactivities of phenolics by focusing on suppression of chronic diseases: A review. International Journal of Molecular Sciences 2018; 19(6), 1573.
GA Pankey and LD Sabath. Clinical relevance of bacteriostatic versus bactericidal mechanisms of action in the treatment of Gram-positive bacterial infections. Clinical Infectious Diseases 2004; 38(6), 864-870.
TT Cushnie and AJ Lamb. Recent advances in understanding the antibacterial properties of flavonoids. International Journal of Antimicrobial Agents 2011; 38(2), 99-107.
I Wiegand, K Hilpert and RE Hancock. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nature Protocols 2008; 3(2), 163-175.
M Takó, EB Kerekes, C Zambrano, A Kotogán, T Papp, J Krisch and C Vágvölgyi. Plant phenolics and phenolic-enriched extracts as antimicrobial agents against food-contaminating microorganisms. Antioxidants 2020; 9(2), 165.
A Lobiuc, NE Pavăl, II Mangalagiu, R Gheorghiță, GC Teliban, D Amăriucăi-Mantu and V Stoleru. Future antimicrobials: Natural and functionalized phenolics. Molecules 2023; 28(3), 1114.
N Ibrahim and WA Yaacob. Disruption of methicillin-resistant Staphylococcus aureus protein synthesis by tannins. Germs 2017; 7(4), 186.
N Shen, T Wang, Q Gan, S Liu, L Wang and B Jin. Plant flavonoids: Classification, distribution, biosynthesis, and antioxidant activity. Food Chemistry 2022; 383, 132531.
OS Faleye, JH Lee and J Lee. Selected flavonoids exhibit antibiofilm and antibacterial effects against Vibrio by disrupting membrane integrity, virulence and metabolic activities. Biofilm 2023; 6, 100165.
MS Mohamed, K Abdelkader, HA Gomaa, AS Batubara, M Gamal and AM Sayed. Mechanistic study of the antibacterial potential of the prenylated flavonoid auriculasin against Escherichia coli. Archiv der Pharmazie 2022; 355(12), 2200360.
W Xu, Z Lin, C Cortez-Jugo, GG Qiao and F Caruso. Antimicrobial phenolic materials: From assembly to function. Angewandte Chemie International Edition 2025; 64(13), e202423654.
Z Breijyeh, B Jubeh and R Karaman. Resistance of gram-negative bacteria to current antibacterial agents and approaches to resolve it. Molecules 2020; 25(6), 1340.
V Priya, S Nagarathna and KH Veena. Molecular characterization of methicillin-resistant Staphylococcus aureus: Dissemination of multidrug-resistant community-associated MRSA and emergence of LA-MRSA, in a healthcare setting. Indian Journal of Medical Microbiology 2025; 54, 100810.
F Sheydai and A Tukmechi. Cell wall disruption, membrane damage, and decrease in the expression of Yrp1 virulence factor in Yersinia ruckeri by propolis ethanol extract. Iranian Journal of Microbiology 2023; 15(4), 533.
M Hartmann, M Berditsch, J Hawecker, MF Ardakani, D Gerthsen and AS Ulrich. Damage of the bacterial cell envelope by antimicrobial peptides gramicidin S and PGLa as revealed by transmission and scanning electron microscopy. Antimicrobial Agents and Chemotherapy 2010; 54(8), 3132-3142.
EHL Chen, CH Wang, YT Liao, FY Chan, Y Kanaoka, T Uchihashi, K Kato, L Lai, YW Chang, MC Ho and RPY Chen. Visualizing the membrane disruption action of antimicrobial peptides by cryo-electron tomography. Nature Communications 2023; 14(1), 5464.