Trends
Sci.
2026;
23(9):
13101
Unlocking Antioxidant, Antibacterial and Anti-Inflammatory Peptides of Spirulina Platensis Through Bromelain Hydrolysis
Zidan
Armanda1,
![]() ,
Yunita Eka Puspitasari1,
,
Yunita Eka Puspitasari1,
![]() ,
Dinia Rizqi Dwijayanti2,
,
Dinia Rizqi Dwijayanti2,
![]() ,
,
Pamungkas
Rizki Ferdian3,
![]() ,
Zahrotul Firdaus4,
,
Zahrotul Firdaus4,
![]() and Heder Djamaludin1,*
and Heder Djamaludin1,*
1Fish Product Technology Study Program, Faculty of Fisheries and Marine Science, Universitas Brawijaya,
Malang East Java 65145, Indonesia
2Department of Biology, Faculty of Mathematics and Natural Sciences, Universitas Brawijaya,
Malang East Java 65145, Indonesia
3Research Center for Applied Zoology, National Research and Innovation Agency (BRIN) Cibinong,
West Java 16911, Indonesia
4Department of Aquatic Product Technology, Faculty of Fisheries and Marine Sciences, IPB University,
West Java 16680, Indonesia
(*Corresponding author’s e-mail: [email protected])
Received: 20 December 2025, Revised: 9 February 2026, Accepted: 16 February 2026, Published: 10 April 2026
Abstract
Spirulina platensis (Arthrospira platensis) is a protein-rich microalga widely recognized for its nutritional value; however, the bioactivity of its native proteins is limited. This study aimed to investigate whether bromelain-assisted enzymatic hydrolysis could enhance the antioxidant, antibacterial, and anti-inflammatory activities of S. platensis proteins. Proteins were extracted by freeze–thawing and sonication, followed by bromelain-mediated hydrolysis at pH 7 and 65 °C for 4 h. The resulting hydrolysates were characterized in terms of yield, degree of hydrolysis, molecular weight distribution, amino acid composition, and functional groups using standard analytical techniques. Bioactivities were assessed through ferric reducing antioxidant power (FRAP), disk diffusion antibacterial assays against Staphylococcus aureus and Escherichia coli, protein denaturation inhibition, and lipopolysaccharide-induced nitric oxide production in RAW 264.7 macrophage cells. The hydrolysis process yielded low-molecular-weight peptides (≤15 kDa) with an increased degree of hydrolysis and protein content compared to the crude extract. The protein hydrolysates exhibited measurable ferric reducing activity in the FRAP assay; however, their antioxidant reducing power was lower than that of the crude protein extract and therefore considered moderate rather than enhanced. In contrast, the hydrolysates demonstrated enhanced antibacterial activity compared with the non-hydrolyzed protein. In anti-inflammatory assays, the hydrolysates significantly inhibited protein denaturation and suppressed nitric oxide production in a concentration-dependent manner while maintaining high cell viability. These findings indicate that bromelain-assisted hydrolysis effectively enhances the bio-functional properties of S. platensis proteins. Thus, S. platensis protein hydrolysates show potential as natural sources of multifunctional bioactive compounds for food and biomedical applications.
Keywords: Bioactive peptides, Cell viability, Enzymatic hydrolysis, Functional biomolecules, Microalgal proteins, Nitric oxide inhibition, RAW 264.7 macrophages
Introduction
Bacterial infections remain a major global health concern, particularly in the context of increasing antimicrobial resistance resulting from the widespread
and irrational use of antibiotics [1]. Pathogenic bacteria such as Escherichia coli and Staphylococcus aureus are frequently associated with urinary tract, wound, and respiratory infections and have exhibited reduced susceptibility to many conventional antimicrobial agents [2]. These infections are commonly accompanied by inflammatory responses, which can exacerbate tissue damage and delay recovery. Consequently, there is growing interest in identifying alternative therapeutic agents that combine antibacterial and anti-inflammatory activities in a single, multifunctional platform [3].
Spirulina platensis, a protein-rich microalga containing approximately 55% - 70% protein, has attracted considerable attention as a sustainable source of bioactive compounds with antimicrobial and anti-inflammatory potential [4]. However, intact proteins often exhibit limited biological activity due to their complex structure and low bioavailability. Enzymatic hydrolysis has therefore emerged as an effective strategy to release low-molecular-weight peptides with enhanced functional properties. Among proteolytic enzymes, bromelain (EC 3.4.22.2) has been reported to efficiently hydrolyze algal and plant proteins, producing higher yields of bioactive hydrolysates compared to commonly used enzymes such as papain, particularly during prolonged hydrolysis [5,6].
The biological activities of protein hydrolysates can be evaluated through complementary in vitro assays. Anti-inflammatory potential is commonly assessed using the bovine serum albumin (BSA) denaturation model, which reflects the ability of compounds to prevent protein denaturation under inflammatory conditions. In addition to this physicochemical assay, cell-based models such as RAW 264.7 murine macrophages are widely employed to provide biologically relevant evidence of anti-inflammatory activity by measuring the inhibition of inflammatory mediators, including nitric oxide (NO) and pro-inflammatory cytokines, in response to lipopolysaccharide (LPS) stimulation. Antibacterial activity is typically evaluated using disk diffusion assays, where inhibition zone formation serves as an initial indicator of antimicrobial potency against pathogenic bacteria [7]. Although these methods are well established, their combined application to assess both antibacterial and anti-inflammatory activities of S. platensis protein hydrolysates remain insufficiently explored.
To date, studies focusing on the simultaneous antioxidant, antibacterial, and anti-inflammatory evaluation through in vitro approach of bromelain-derived S. platensis protein hydrolysates are scarce, and the potential of bromelain to unlock multifunctional bioactive peptides from this microalga has not been comprehensively addressed. Therefore, the novelty of this study lies in the integrated assessment of antioxidant, antibacterial, and anti-inflammatory properties of S. platensis protein hydrolysates produced through bromelain hydrolysis. The aim of this research is to investigate the biological activities of these hydrolysates, highlighting their potential as natural, sustainable, and multifunctional agents for future pharmaceutical or biotechnological applications.
Materials and methods
Freshwater Spirulina platensis biomass was obtained from the Balai Besar Perikanan Budidaya Air Payau (BBPBAP), Jepara, Indonesia. Bromelain enzyme (Brand Xian Best BioTech Co., China; activity 10,400 U/mg) was used for protein hydrolysis. Analytical-grade reagents included distilled water, biuret reagent, bovine serum albumin (BSA; Merck), sodium hydroxide (NaOH; Merck, Germany), hydrochloric acid (HCl; Merck, Germany), ascorbic acid (Merck, Germany), ethanol (Merck, Germany), methanol (Smart Lab), ninhydrin reagent (Smart Lab), and 2,4,6-tripyridyl-s-triazine (TPTZ; Smart Lab). Whatman GF/C filter paper and Millipore membrane filters (0.45 µm) were used for filtration. Microbiological media and materials included Mueller–Hinton Agar (HiMedia, GM173), Nutrient Broth (Oxoid, CM0001), and antibiotic discs (Macherey–Nagel). The bacterial strains used for antibacterial assays were Staphylococcus aureus subsp. aureus (ATCC 33591) and Escherichia coli (ATCC 10798). For anti-inflammatory assays, the murine macrophage cell line RAW 264.7 was obtained from the Department of Biology, Universitas Brawijaya, Indonesia, and used for in vitro cell-based experiments.
Protein extraction
Proteins from Spirulina platensis were extracted using a combined physical disruption approach involving freeze–thaw cycles and ultrasonication, with minor modifications from Wang and Zhang [8]. Briefly, 5 g of S. platensis powder was suspended in 100 mL of distilled water and homogenized using a magnetic stirrer for 30 min. The suspension was subjected to three freeze–thaw cycles, consisting of freezing at –20 °C for 4 h followed by thawing at room temperature (20 - 25 °C) for 4 h to facilitate cell wall disruption. Subsequently, the mixture was ultrasonicated in an ultrasonic bath (40 kHz) for 20 min while maintaining the temperature at 27 °C. The extract was centrifuged at 10,000 rpm for 30 min at 4 °C, and the resulting supernatant was filtered through Whatman GF/C filter paper (1.2 μm pore size). The clarified protein extract was collected and stored at 4 °C for subsequent bromelain-mediated enzymatic hydrolysis.
Protein hydrolysis
Proteins from Spirulina platensis were hydrolyzed enzymatically using bromelain to generate bioactive peptides with potential antibacterial and anti-inflammatory properties. The hydrolysis procedure was adapted from Djamaludin et al. [5] with minor modifications. Briefly, S. platensis protein extract was mixed with bromelain (enzyme activity: 10,400 U/mg) at a final enzyme concentration of 4 %(w/v) in 100 mL of substrate solution. Enzymatic hydrolysis was conducted at 65 °C and pH 7.0 for 4 h under controlled conditions. The reaction was terminated by heating the mixture at 90 °C for 10 min to inactivate the enzyme. The hydrolysate was subsequently centrifuged at 10,000 rpm for 20 min at 4 °C, and the resulting supernatant was collected. To obtain the protein hydrolysate powder, the supernatant was pre-frozen at –80 °C for 3 h and then freeze-dried at –55 °C under a vacuum pressure of 0.04 mbar. The dried hydrolysate was stored at –20 °C until further analysis.
Protein content analysis
The total protein content of S. platensis was determined using a modified Micro-Kjeldahl method in accordance with AOAC Official Method [9]. The analysis involved sequential digestion, distillation, and titration steps to quantify total nitrogen present in the sample. Protein content was calculated by applying a nitrogen-to-protein conversion factor of 5.95, which is appropriate for microalgal protein analysis. The determined protein values were used as the basis for subsequent enzymatic hydrolysis and bioactivity evaluation.
Degree of hydrolysis (DH) analysis
The degree of hydrolysis (DH) of S. platensis protein hydrolysates obtained through bromelain treatment was determined using a spectrophotometric ninhydrin assay, following the method of Djamaludin et al. [5] with minor modifications. Leucine was employed as the reference amino acid for calibration. A standard curve was prepared using leucine solutions at concentrations of 0 - 500 ppm. Samples and standards were reacted with ninhydrin reagent and incubated at 80 °C for 4 min to allow chromophore development. The absorbance was measured at 570 nm using a UV–Vis spectrophotometer. The DH was calculated based on the concentration of free amino groups released during enzymatic hydrolysis, as determined from the leucine standard curve.
Fourier transform infrared spectroscopy analysis
Fourier Transform Infrared (FTIR) spectroscopy was employed to characterize structural features of S. platensis protein hydrolysates produced by bromelain hydrolysis, following a modified method of Domenighini and Giordano [10]. Microalgal biomass was harvested by centrifugation, washed with ammonium formate solution to minimize medium interference, and deposited onto silicon windows. Samples were dried at 60 °C for 3 h prior to analysis. FTIR spectra were recorded using a Tensor 27 spectrometer (Bruker, Germany) in the range of 3,500 - 500 cm⁻¹, with a spectral resolution of 4 cm⁻¹ and 32 accumulated scans. Spectral preprocessing, including baseline correction and normalization to the amide I band (~1,655 cm⁻¹), was performed to correct for variations in sample thickness. Data analysis and hierarchical cluster analysis (HCA) were conducted using OPUS 6.0 software to evaluate similarities in protein structural profiles following enzymatic hydrolysis.
Amino acid profile analysis
The amino acid composition of S. platensis protein hydrolysates was determined using high-performance liquid chromatography (HPLC) following the method described by Ali et al. [11], with minor modifications. Samples were dissolved in 0.01 N HCl and filtered through a 0.45 µm membrane prior to analysis. Amino acids were derivatized using ortho-phthalaldehyde (OPA) reagent prepared in methanol containing mercaptoethanol, Brij-30, and borate buffer (pH 10.4) to enhance reagent stability. For derivatization, 10 µL of sample solution was mixed with 50 µL of OPA reagent and allowed to react for 1 min at room temperature before injection. Chromatographic separation was performed on a C18 reversed-phase column (250×4.6 mm, 5 µm) maintained at 30 °C. The mobile phase consisted of solvent A (50 mM sodium acetate buffer, pH 6.8) and solvent B (acetonitrile–methanol–water, 45:45:10, v/v/v). Separation was achieved using a gradient elution as follows: 0 - 5 min, 5% B; 5 - 20 min, 5% - 45% B; 20 - 25 min, 45% - 100% B, at a flow rate of 1.0 mL/min. OPA-derivatized amino acids were detected using a fluorescence detector with excitation and emission wavelengths set at 340 and 450 nm, respectively. The injection volume was 20 µL, and the total run time was 25 min.
Molecular weight analysis
Proteins from S. platensis were enzymatically hydrolyzed using bromelain following a modified procedure reported by Djamaludin et al. [5]. Briefly, S. platensis protein extract was suspended in distilled water, and bromelain (enzyme activity: 10,400 U/mg) was added at a final concentration of 4 %(w/v) in a total reaction volume of 100 mL. Enzymatic hydrolysis was conducted at 65 °C and pH 7 for 4 h under constant agitation to facilitate peptide release. The reaction was terminated by heating the mixture in a water bath at 90 °C for 10 min to inactivate the enzyme. The resulting hydrolysate was centrifuged at 10,000 rpm for 20 min at 4 °C to remove insoluble residues. The supernatant was collected, pre-frozen at −80 °C for 3 h, and subsequently freeze-dried at −55 °C under a vacuum pressure of −0.04 mbar to obtain bromelain-derived Spirulina platensis protein hydrolysate powder for further antibacterial and anti-inflammatory analyses.
Antioxidant assay
The antioxidant capacity of S. platensis protein hydrolysates was evaluated using the Ferric Reducing Antioxidant Power (FRAP) assay with minor modifications from Firdaus et al. [12]. The FRAP reagent was freshly prepared by mixing sodium acetate buffer (300 mM, pH 3.6), 2,4,6-tripyridyl-s-triazine (TPTZ, 10 in 40 mM HCl), and FeCl₃·6H₂O (20 mM) at a volume ratio of 10:1:1 (v/v/v). The resulting solution was equilibrated to room temperature prior to analysis. An aliquot of 5 µL of sample was added to 195 µL of FRAP reagent and incubated for 15 min in the dark. Absorbance was then measured at 593 nm using a microplate reader. Trolox was used as a positive control to generate standard solutions at concentrations of 25 - 125 ppm, and the results were expressed as Trolox equivalents (µg/mL).
Antibacterial assay
The antibacterial activity of S. platensis crude extract and bromelain-derived protein hydrolysates was evaluated using the disk diffusion method, adapted from Sun et al. [13] with minor modifications. Briefly, bacterial suspensions of Escherichia coli and Staphylococcus aureus (200 µL) were inoculated into sterile Petri dishes, followed by the addition of 20 mL of molten Mueller–Hinton agar (MHA). The plates were gently swirled to ensure uniform distribution of the bacteria and allowed to solidify at room temperature. Sterile paper discs were then placed on the agar surface. The crude extract and protein hydrolysates were dissolved in distilled water and tested at four concentrations (625, 1,250, 2,500, and 5,000 µg/mL). Each disc was impregnated with 20 µL of the test solution. Chloramphenicol (30 µg/disc) was used as the positive control, while distilled water served as the negative control. All plates were incubated at 37 °C for 24 h, after which antibacterial activity was determined by measuring the diameter of the inhibition zones using a digital caliper.
MIC and MBC Assay
The minimum inhibitory concentration (MIC) was determined using the broth microdilution method with minor modifications to reduce visual interference from complex samples. Overnight bacterial cultures grown in Mueller–Hinton broth (MHB) were adjusted to 0.5 McFarland standard (≈1×10⁶ CFU/mL). Serial two-fold dilutions of the test sample were prepared in MHB in 96-well microplates and inoculated to obtain a final bacterial concentration of approximately 5×10⁵ CFU/mL. Wells containing inoculated MHB without the test sample and uninoculated MHB served as growth and sterility controls, respectively. Plates were incubated at 37 °C for 18 - 24 h, and MIC was defined as the lowest concentration showing no visible turbidity compared with the growth control. When visual assessment was affected by sample turbidity or color, optical density at 600 nm was used to support MIC determination. Minimum bactericidal concentration (MBC) was determined by subculturing aliquots (10 - 20 µL) from wells at the MIC and selected adjacent concentrations onto Mueller–Hinton agar (MHA). Plates were incubated at 37 °C for 24 h. MBC was defined as the lowest concentration resulting in ≥99.9% reduction in viable bacteria, indicated by the absence of colony growth or the presence of fewer than three colonies. All assays were performed in triplicate [14].
Anti-inflammatory by bovine serum albumin (BSA) denaturation assay
The anti-inflammatory potential of S. platensis protein hydrolysates generated through bromelain hydrolysis was evaluated using the bovine serum albumin (BSA) denaturation assay, with minor modifications to the method described by Bailey-Shaw et al. [15]. Tris-buffered saline (TBS) was prepared by dissolving sodium chloride and Tris-base in distilled water, followed by pH adjustment to 6.2 - 6.5 using glacial acetic acid. A 0.2 %(w/v) BSA solution was prepared in TBS and used as the protein substrate. Diclofenac sodium was employed as the reference anti-inflammatory drug and prepared as a stock solution, followed by serial dilution to obtain working concentrations of 25, 50, and 100 µg/mL. The reaction mixtures consisted of the hydrolysate samples, positive control, or negative control combined with the BSA solution to a final volume of 5 mL. All samples were incubated at room temperature for 30 min, subjected to thermal denaturation at 72 °C for 5 min, and subsequently cooled to ambient temperature. Absorbance was measured at 660 nm using a UV–Vis spectrophotometer. Inhibition of protein denaturation was calculated to determine anti-inflammatory activity.
Anti-inflammatory by RAW 264.7 cells assay
Cell culture
Murine macrophage RAW 264.7 cells were used to evaluate the anti-inflammatory activity of S. platensis protein hydrolysates generated by bromelain hydrolysis. Cells were cultured in Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 10 %(v/v) fetal bovine serum (FBS) and 1 %(v/v) penicillin–streptomycin solution. The cells were seeded into 24-well plates at a volume of 500 μL per well and incubated for 24 h at 37 °C in a humidified atmosphere containing 5% CO₂ [16].
Nitric oxide (NO) inhibition assay
To assess the anti-inflammatory potential of the hydrolysates, RAW 264.7 cells were stimulated with lipopolysaccharide (LPS, 1 μg/mL) in the presence or absence of S. platensis protein hydrolysates at concentrations ranging from 1.25 to 320 μg/mL. After 24 h of incubation, the culture supernatants were collected and mixed with Griess reagent to quantify nitric oxide (NO) production. Absorbance was measured at 571 nm using a microplate reader, and NO levels were expressed relative to the LPS-treated control [16].
Cell viability assay
Cell viability was evaluated using the WST-1 assay to determine the cytotoxicity of the protein hydrolysates. Following treatment, the culture medium was replaced with 120 μL of fresh medium containing 7.5 μL of WST-1 reagent and incubated for 30 min at 37 °C. Subsequently, 100 μL of the reaction mixture was transferred to a 96-well plate, and absorbance was measured at 450 nm. Cell viability was expressed as a percentage relative to untreated control cells [16].
Statistical analysis
All data were presented as mean ± standard error of the mean (SEM). Statistical analyses were performed using Minitab (version 22) with the necessary statistical packages. Descriptive analysis was used to summarize the parameters. Inferential statistics, specifically ANOVA, with a significance level of p-value < 0.05.
Results and discussion
Protein content
Bromelain-mediated hydrolysis of Spirulina platensis resulted in a measurable increase in soluble protein content, indicating effective protein breakdown and peptide release. As shown in Table 1, the protein content was 0.89% in the crude extract and 0.94% in the hydrolysate powder. S. platensis is well known for its high protein content, which can account for up to 70% of its dry biomass [17]. However, intact cellular structures and complex protein matrices often limit the bioavailability of these proteins. Enzymatic hydrolysis using bromelain facilitates the cleavage of peptide bonds, generating low-molecular-weight peptides with improved solubility and functional activity. This mechanism explains the observed increase in measurable protein content following hydrolysis and aligns with previous reports demonstrating that enzymatic treatments are more effective than physical methods, such as ultrasonication, in releasing intracellular proteins [18].
Table 1 Protein content of crude extract and protein hydrolysate of Spirulina platensis.
Sample |
Protein content (%) |
Crude Extract Protein (dried biomass) |
0.89 |
Hydrolysate Protein (hydrolysate powder) |
0.94 |
Previous studies have shown that protease-assisted hydrolysis can disrupt cell wall components and liberate bioactive peptides with enhanced biological functions, including antibacterial and anti-inflammatory activities [19,20]. In particular, bromelain has been reported to generate peptides with improved interaction with bacterial membranes and inflammation-related molecular targets [21]. The reduction in protein molecular weight following hydrolysis is also known to enhance membrane permeability and cellular interactions, thereby increasing antibacterial and anti-inflammatory efficacy [22]. Therefore, the observed increase in soluble protein content in this study supports the hypothesis that bromelain hydrolysis effectively unlocks bioactive peptides from S. platensis, contributing to its functional potential as a natural source of antibacterial and anti-inflammatory agents.
%Yield of crude extract and hydrolysate protein Spirulina platensis
Protein yield is a key indicator of the effectiveness of extraction and hydrolysis processes in generating bioactive compounds. As shown in Table 2, the crude protein extraction from S. platensis resulted in a relatively low yield (16%), indicating limited protein recovery using the combined sonication and freeze–thaw approach. This outcome suggests that the applied method was insufficient to disrupt the rigid and complex cell wall structure of S. platensis, which is known to restrict the release of intracellular proteins. Similar extraction challenges have been reported for microalgal biomass with robust cell matrices, where physical disruption alone often fails to achieve maximal protein recovery.
Table 2 %Yield of crude extract and protein hydrolysate of Spirulina platensis.
Sample |
Initial weight (g) |
Recovery after hydrolysis/freeze-drying (g) |
Yield (%) |
Crude extract protein Spirulina platensis |
5 |
0.8 |
16 |
Protein hydrolysate Spirulina platensis |
0.8 |
0.54 |
67.5 |
In contrast, bromelain-assisted hydrolysis markedly increased the yield to 67.5%, demonstrating the enzyme’s effectiveness in cleaving complex proteins into smaller, more soluble peptides. This observation highlights the ability of enzymatic hydrolysis to enhance protein solubilization and release peptides with potential biological activity [6]. These findings highlight bromelain hydrolysis as a critical step in unlocking bioactive peptides from S. platensis, supporting its subsequent antibacterial and anti-inflammatory potential.
Degree of hydrolysis
Bromelain hydrolysis significantly enhanced the breakdown of S. platensis proteins. The degree of hydrolysis (DH), determined using the ninhydrin spectrophotometric method, was 60.515 ± 1.058%. These results indicate that bromelain effectively cleaves Spirulina proteins into smaller peptide fractions that are commonly associated with enhanced biological activity. The degree of hydrolysis (DH) demonstrates the efficiency of bromelain as a proteolytic enzyme with broad substrate specificity, particularly toward amino acid residues such as arginine, lysine, tyrosine, and phenylalanine that are abundant in S. platensis proteins. Higher enzyme concentrations likely enhanced peptide bond cleavage, promoting the release of low-molecular-weight peptides associated with antibacterial and anti-inflammatory activities, thereby supporting the use of bromelain for producing functional peptides from microalgal proteins. Amino acid analysis confirmed protein hydrolysis, showing an increase in amino acid content from 0.66% in the crude extract to 0.76% in the hydrolysate, exceeding the standard reference value (0.6%), with an estimated DH of approximately 15%; differences between DH values obtained by spectrophotometric and amino acid-based methods are expected due to variations in methodological sensitivity and detected hydrolysis products.
The DH value obtained in this study is consistent with previous reports, such as Lisboa et al. [23], that reported a DH of approximately 18%, and Noman et al. [24], who reported a lower degree of hydrolysis (DH) of 6.43% is typically obtained by controlling enzymatic conditions such as using lower enzyme concentrations, shorter hydrolysis times, or adjusting the pH and temperature away from the enzyme's optimal range. These variations underscore the importance of enzyme type and concentration in tailoring peptide profiles. Collectively, the results demonstrate that bromelain hydrolysis is a robust approach for unlocking bioactive peptides from Spirulina platensis, providing a biochemical basis for the antibacterial and anti-inflammatory activities explored in subsequent analyses.
Fourier transform infrared spectroscopy
The FTIR spectra of the crude extract and bromelain-produced protein hydrolysate of S. platensis (Figure 1) display several absorption bands corresponding to functional groups commonly associated with proteins and other biomolecules. The hydrolysate spectrum is characterized by prominent peaks at approximately 3,845.90 and 3,733.88 cm⁻¹, which are attributed to O–H stretching vibrations, indicating the presence of hydroxyl groups and hydrogen bonding interactions. A broad absorption band observed around 3,300.18 cm⁻¹ is associated with N–H stretching vibrations of amide groups, confirming the presence of proteinaceous components. Additional bands at 2,355.22 and 1,640.04 cm⁻¹ correspond to C–H stretching and amide I (C=O stretching) vibrations, respectively, which are characteristic of peptide backbones.
The crude extract exhibits similar functional group signatures; however, noticeable differences in peak position and intensity are evident, particularly at 3,857.39, 3,731.01 and 3,317.41 cm⁻¹. These variations suggest alterations in molecular interactions and bonding environments following enzymatic treatment. The presence of comparable absorption regions in both samples indicates that proteins, polysaccharides, and other bioactive constituents are retained in the hydrolysate, although their structural organization has been modified by bromelain hydrolysis.
Significant spectral changes were observed after enzymatic hydrolysis, particularly within the amide I, II, and III regions, which are closely related to protein secondary structures. Shifts in peak position and changes in band intensity in these regions reflect conformational modifications of the protein matrix due to peptide bond cleavage. The appearance of new absorption features and the redistribution of peak intensities indicate the breakdown of high-molecular-weight proteins into smaller peptides during hydrolysis. These findings are consistent with previous studies reporting that enzymatic hydrolysis reduces protein molecular size and enhances bioactive potential [25].
Moreover, the reduced absorption intensity in the 1,600 - 1,700 cm⁻¹ region suggests disruption of the original protein conformation, as the amide I and II bands are particularly sensitive to changes in secondary structure. The weakening of these bands confirms that bromelain hydrolysis alters the native protein arrangement [26]. According to Ji et al. [27], such enzymatic treatments induce detectable conformational changes in protein structures that can be effectively monitored using FTIR spectroscopy. The observed spectral modifications demonstrate that bromelain hydrolysis significantly modifies the structural characteristics of S. platensis proteins, thereby increasing peptide availability and potentially improving their functional and biological properties.
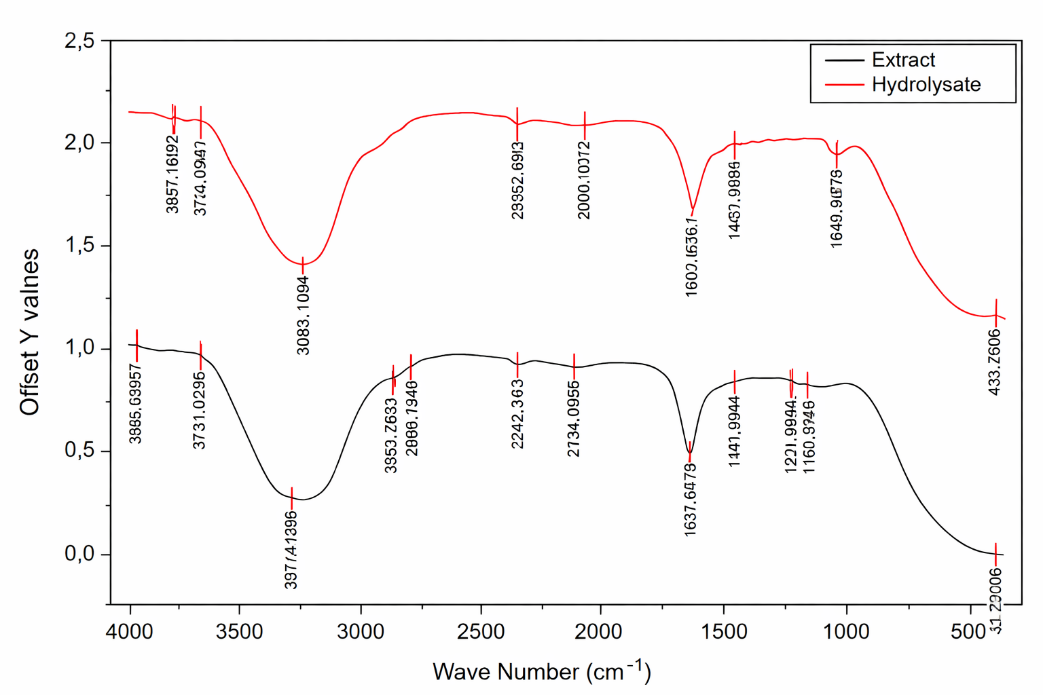
Figure 1 FTIR spectrum of crude extract and hydrolysate protein of Spirulina platensis.
Amino acid profile
The amino acid composition of S. platensis crude extract and its bromelain-derived protein hydrolysate is summarized in Table 3. Enzymatic hydrolysis led to an increase in the concentration of most amino acids, reflecting efficient proteolytic activity by bromelain and the subsequent breakdown of complex proteins into smaller peptides and free amino acids. This increase confirms the effectiveness of bromelain in enhancing protein accessibility and amino acid release. Among the hydrophilic amino acids, L-glutamic acid was the predominant component in both samples and exhibited a marked increase after hydrolysis. This finding emphasizes the natural abundance of glutamate in Spirulina proteins and its high susceptibility to enzymatic cleavage. Similar upward trends were observed for other polar amino acids, including L-aspartic acid, L-arginine, L-serine, and L-threonine. The increased presence of these hydrophilic residues suggests improved solubility and greater availability of polar peptide fractions in the hydrolysate. In contrast, L-lysine showed a slight decrease after hydrolysis, which may be attributed to selective enzyme specificity or further degradation into smaller nitrogenous compounds.
Bromelain treatment also enhanced the levels of several hydrophobic amino acids, such as L-alanine, glycine, L-leucine, L-valine, L-isoleucine, and L-proline. The enrichment of hydrophobic residues is particularly significant, as these amino acids are frequently associated with peptides possessing biological activities, including antibacterial and anti-inflammatory effects. Hydrophobic amino acids facilitate interactions with lipid bilayers of bacterial membranes, promoting membrane destabilization, and also support intracellular penetration that may influence inflammatory signaling pathways. Consequently, the observed increase in hydrophobic amino acids indicates that the protein hydrolysate may exhibit functional properties beyond basic nutritional value. Aromatic amino acids, specifically L-tyrosine and L-phenylalanine, were detected below the quantification limits in both the crude extract and hydrolysate. Despite their low measurable concentrations, these amino acids are recognized for their contribution to antioxidant and bioactive functions due to their ability to donate protons and stabilize free radicals through resonance structures. Their low detectability may reflect the intrinsic amino acid composition of S. platensis proteins or analytical sensitivity limitations rather than their complete absence.
The results of this study align well with previous findings reported in the literature. Akbarbaglu et al. [28] documented significant increases in essential and hydrophobic amino acids following enzymatic hydrolysis of Spirulina, along with improved protein efficiency ratios. Similarly, Ling et al. [29] stated that the advanced processing techniques that enhance the availability and accumulation of amino acids, particularly glutamate, alanine, and proline, in food products involve enzymatic hydrolysis, often combined with other methods such as fermentation or novel physical processing. When compared to the natural amino acid profiles of Spirulina reported by Rivera-Jiménez et al. [30] mall peptides (<1 kDa) with 2 - 20 amino acid residues are generally more active because they are more easily absorbed in the intestine and can enter cells more efficiently than larger peptides. Anti-inflammatory peptides often contain hydrophobic amino acids (Val, Ile, Pro) and positively charged amino acids (His, Arg, Lys). Notably, about 77% of these bioactive peptides originate from alternative and sustainable protein sources such as Tenebrio molitor and sunflower. These findings collectively support the potential application of bromelain-hydrolyzed S. platensis proteins as functional and bioactive ingredients.
Table 3 Amino acid profile of crude extract and protein hydrolysate of Spirulina platensis.
Amino acid type |
Amino acid |
Amino acid content |
|
Crude extract protein of Spirulina platensis (mg/g) |
Hydrolysate protein of Spirulina platensis (mg/g) |
||
Hydrophilic |
L-Glutamic Acid |
1,322.26 |
1,406.26 |
|
L-Aspartic Acid |
682.76 |
695.18 |
|
L-Arginine |
490.77 |
590.79 |
|
L-Serine |
398.95 |
501.7 |
|
L-Lysine |
379.19 |
368.75 |
|
L-Threonine |
377.75 |
427.57 |
Hydrophobic |
L-Alanine |
600.51 |
662.2 |
|
Glycine |
457.08 |
507.75 |
|
L-Leucine |
428.42 |
499.34 |
|
L-Valine |
398.95 |
469 |
|
L-Isoleucine |
314.24 |
356.96 |
|
L-Proline |
233.88 |
270.87 |
Aromatic |
L-Tyrosine |
<608.01 |
<608.01 |
|
L-Phenylalanine |
<476.07 |
<476.07 |
Molecular weight
The molecular weight distribution of S. platensis proteins before and after bromelain hydrolysis was evaluated by SDSPAGE, as presented in Figure 2. The crude protein extract exhibited multiple distinct bands distributed across a wide molecular weight range, with prominent bands observed in the high- and intermediate-molecular-weight regions. This pattern indicates the predominance of intact protein structures prior to enzymatic treatment, consistent with the strong amide I and II absorption bands observed in the FTIR spectrum of the crude extract, which reflect preserved peptide backbone integrity.
In contrast, the bromelain-derived protein hydrolysate showed a markedly altered electrophoretic profile characterized by the substantial reduction or disappearance of high-molecular-weight bands and the dominance of low-molecular-weight peptides, particularly in the range of approximately 6 - 7 kDa. This shift toward smaller peptide fractions confirms the effective proteolytic activity of bromelain and is in agreement with the FTIR results, which demonstrated decreased intensity and structural modifications in the amide I and II regions. These spectral changes collectively indicate cleavage of peptide bonds and disruption of the original protein secondary structure.

Figure 2 Profile of molecular weight crude extract and protein hydrolysate of Spirulina platensis (1. marker; 5. crude extract protein; 3. hydrolysate protein).
The generation of low-molecular-weight peptides observed in the SDS-PAGE analysis is further supported by the amino acid composition results. Bromelain hydrolysis led to an overall increase in free and accessible amino acids, particularly hydrophilic residues such as glutamic acid, aspartic acid, arginine, serine, and threonine, as well as hydrophobic amino acids including alanine, leucine, valine, isoleucine, and proline. The enrichment of these amino acids reflects extensive protein breakdown and correlates with the accumulation of short peptide chains detected in the molecular weight profile.
Importantly, peptides smaller than 10 kDa often show improved solubility and membrane permeability; however, linking their reduced size directly to enhanced biological activity is an oversimplification, as activity depends on peptide sequence, target interactions, and specific biological functions [31,32]. The dominance of low-molecular-weight peptides in the bromelain hydrolysate, together with the increased abundance of bioactivity-related hydrophobic and polar amino acids, supports the functional potential of the hydrolysate. These findings are consistent with previous reports demonstrating that enzymatic hydrolysis simultaneously alters protein conformation (as evidenced by FTIR), reduces molecular size (as shown by SDS-PAGE), and enhances amino acid availability, ultimately promoting the release of bioactive peptides [33]. The combined SDS-PAGE, FTIR, and amino acid analyses provide complementary evidence that bromelain hydrolysis effectively modifies S. platensis proteins at both structural and compositional levels, resulting in peptide fractions with improved functional and biological properties.
Antioxidant activity
The FRAP results expressed as Trolox equivalents (µg/mL) demonstrated a concentration-dependent increase in antioxidant activity for both crude protein extracts and protein hydrolysates (Table 4). The FRAP values of the crude protein extract increased from 223.69 µg TE/mL at a concentration of 25 µg/mL to 319.24 µg TE/mL at 125 µg/mL, while those of the protein hydrolysate increased from 65.54 to 147.30 µg TE/mL over the same concentration range. At all tested concentrations, the crude protein extract consistently exhibited higher FRAP values than the protein hydrolysate. The relatively lower antioxidant activity of the hydrolysate is likely related to excessive enzymatic hydrolysis, which may degrade bioactive peptides into free amino acids; antioxidant capacity is known to depend more on peptide structure and specific amino acid sequences than on total amino acid content alone [34]. In the FRAP assay, higher antioxidant activity corresponds to a greater ability of the samples to reduce Fe³⁺–TPTZ to Fe²⁺–TPTZ, reflecting stronger electron‐donating capacity [35]. Peptide antioxidant activity is strongly influenced by amino acid composition: Aromatic amino acids such as tyrosine (Tyr) and phenylalanine (Phe) contribute through proton donation, hydrophobic residues such as proline (Pro) and leucine (Leu) enhance interactions with target membranes, and hydrophilic residues such as glutamate (Glu) and aspartate (Asp) act as metal ion chelators. This combination of amino acid characteristics underlies the high antioxidant capacity of S. platensis proteins and supports their role in reducing oxidative stress, which is closely associated with the suppression of inflammatory pathways such as NF-κB and the subsequent reduction of inflammatory mediators like nitric oxide [36].
Previous studies support these findings and highlight the strong antioxidant potential of S. platensis proteins. Htoo et al. [37] reported that the protein hydrolysate of Spirulina (A. platensis) exhibited notable antioxidant activity, with a FRAP value reaching approximately 1.2 mg GAE/g extract, indicating a strong iron-reducing capacity as reported in the study. Different extraction methods produce Spirulina extracts with strong but variable antioxidant activity, as reflected by differing IC₅₀ values depending on the compounds recovered and extraction conditions. Advanced techniques such as microwave-assisted extraction, optimized freeze–thawing, and enzymatic hydrolysis generally yield higher potency than conventional maceration. Enzymatic hydrolysis is one of the most effective methods for obtaining bioactive peptides from S. platensis proteins, as it uses proteases to cleave intact proteins and release peptides with nutritional and medicinal value. Recent studies have shown that Spirulina platensis proteins and their enzymatic hydrolysates exhibit various physiological activities, including antioxidant, antimicrobial, anticancer, anti-inflammatory, immunomodulatory, and hypocholesterolemic effects [38-40].
Table 4 Antioxidant activity of crude extract and protein hydrolysate of Spirulina platensis.
Concentration (µg/mL) |
Sample trolox equivalent (µg TE/mL) |
|
Crude extract protein |
Hydrolysate protein |
|
25 |
223.69 ± 1624 |
65.54 ± 7.07 |
50 |
248.22 ± 26.32 |
72.39 ± 11.26 |
75 |
292.39 ± 4.71 |
88.13 ± 10.21 |
100 |
308.41 ± 7.72 |
105.07 ± 6.15 |
125 |
319.24 ± 5.49 |
147.29 ± 15.06 |
Antibacterial activity
Based on Table 5, the antibacterial assay clearly demonstrates that enzymatic hydrolysis significantly enhances the antibacterial efficacy of S. platensis hydrolysate protein compared to the crude extract. The protein hydrolysate consistently produced larger inhibition zones against both Gram-positive Staphylococcus aureus and Gram-negative Escherichia coli at all tested concentrations (625 - 5,000 µg/mL), indicating a stronger and more stable antibacterial effect. This concentration-dependent increase in inhibition zones suggests that higher peptide availability enhances interaction with bacterial cells. Notably, the hydrolysate showed greater activity against E. coli than S. aureus, particularly at 5,000 µg/mL (6.47 ± 0.52 mm vs. 4.73 ± 0.40 mm), which may be attributed to differences in cell wall structure; small bioactive peptides are better able to penetrate the thinner peptidoglycan layer and outer membrane of Gram-negative bacteria. The improved antibacterial performance of the hydrolysate can be explained by bromelain-mediated cleavage of proteins into low-molecular-weight peptides with higher surface charge and hydrophobic amino acid content, enabling stronger electrostatic interactions with negatively charged bacterial membranes, membrane disruption, and leakage of intracellular components. In contrast, the crude extract showed weaker inhibition, likely due to its larger and more complex protein structures that limit membrane penetration. The absence of inhibition in the distilled water control confirms that antibacterial effects originate solely from bioactive compounds, while the strong inhibition by chloramphenicol validates the assay reliability. Statistical analysis (ANOVA and Tukey’s test, p < 0.05) further confirms that protein hydrolysis significantly improves antibacterial activity.
The antibacterial activity results clearly demonstrated that the protein hydrolysate of S. platensis exhibited stronger antibacterial effects than the crude extract at all tested concentrations against both S. aureus and E. coli. Enzymatic hydrolysis using bromelain breaks down complex proteins into smaller peptides, which can more readily penetrate bacterial cell membranes and interact with intracellular targets, thereby inhibiting bacterial growth more effectively [41]. This structural simplification enhances the ability of bioactive peptides to disrupt membrane integrity and interfere with essential cellular processes, leading to improved antibacterial activity. Consequently, the protein hydrolysate consistently produced larger inhibition zones than the crude extract at equivalent concentrations. The positive control, chloramphenicol, produced large inhibition zones against both bacterial strains, confirming the validity and sensitivity of the disk diffusion assay, while distilled water, used as a negative control, showed no inhibitory effect, indicating that the observed antibacterial activity originated solely from the active compounds in the samples [42].
Table 5 Antibacterial activity affected by crude extract and protein hydrolysate of Spirulina platensis.
Bacteria |
Concentration (g/mL) |
Inhibition zone (mm) |
|
Crude extract protein |
Hydrolysate protein |
||
Chloramphenicol (+) |
20.40 0.36e |
20.40 0.36e |
|
Distilled Water (-) |
0.00 0.00a |
0.00 0.00a |
|
S. aureus |
625 |
1.16 0.15b |
2.38 0.37b |
1,250 |
1.87 0.16bc |
3.33 0.19c |
|
2,500 |
2.46 0.35c |
3.71 0.25c |
|
5,000 |
4.13 0.36d |
4.73 0.40d |
|
Chloramphenicol (+) |
20.40 0.36e |
20.40 0.36e |
|
Distilled Water (-) |
0.00 0.00a |
0.00 0.00a |
|
E. coli |
625 |
1.56 0.03ab |
2.89 0.13b |
1,250 |
2.04 0.38bc |
3.74 0.27b |
|
2,500 |
2.05 0.85bc |
4.66 0.34c |
|
5,000 |
3.42 1.03c |
6.47 0.52d |
|
Note: The results in the same column followed by the same letter do not significantly differ by Tukey’s test (p < 0.05).
MIC and MBC
Based on Table 6, the MIC and MBC results clearly demonstrate that the protein hydrolysate of S. platensis exhibits stronger antibacterial activity than the crude extract against both S. aureus and E. coli. The protein hydrolysate showed consistently lower MIC and MBC values compared to the crude extract, indicating a higher effectiveness in inhibiting bacterial growth and inducing bacterial cell death at lower concentrations. For S. aureus, the MIC and MBC values of the protein hydrolysate (1.234 and 0.309 µg/mL, respectively) were lower than those of the crude extract (2.058 and 0.514 µg/mL), while for E. coli, the protein hydrolysate also demonstrated superior activity with MIC and MBC values of 1.380 and 0.345 µg/mL compared to 2.541 and 0.635 µg/mL for the crude extract. This enhanced antibacterial activity is likely attributable to the enzymatic hydrolysis process, which generates smaller bioactive peptides capable of more effectively interacting with and disrupting bacterial cell membranes.
The protein hydrolysate of S. platensis demonstrated a stronger ability to inhibit and kill both Gram-positive (S. aureus) and Gram-negative (E. coli) bacteria compared to the crude extract, as indicated by consistently lower MIC and MBC values. Enzymatic hydrolysis produces low-molecular-weight peptides that are more bioactive and able to interact effectively with bacterial cells. In Gram-positive bacteria such as S. aureus, which possess a thick but relatively porous peptidoglycan layer, these peptides can readily penetrate the cell wall and disrupt cytoplasmic membrane integrity, leading to growth inhibition and cell death at lower concentrations. In Gram-negative bacteria like E. coli, although the outer membrane acts as an additional barrier, the amphipathic and cationic nature of the hydrolysate peptides facilitates their interaction with negatively charged lipopolysaccharides, increasing membrane permeability and causing leakage of intracellular components [43,44]. The lower MIC and MBC values observed for the hydrolysate against both bacteria suggest that hydrolysis enhances antibacterial potency by increasing peptide accessibility, surface activity, and membrane-disruptive mechanisms. The protein hydrolysate of Spirulina platensis is effective against both Gram-positive and Gram-negative bacteria, with its antibacterial action primarily attributed to peptide-mediated membrane disruption and subsequent inhibition and killing of bacterial cells.
Table 6 MIC and MBC of crude extract and protein hydrolysate of Spirulina platensis.
Bacteria |
Sample |
MIC (g/mL) |
MBC (g/mL) |
S. aureus |
Crude Extract |
2.058 |
0.514 |
|
Hydrolysate Protein |
1.234 |
0.309 |
E. coli |
Crude Extract |
2.541 |
0.635 |
|
Hydrolysate Protein |
1.380 |
0.345 |
Anti-inflammatory activity by bovine serum albumin (BSA) denaturation
The anti-inflammatory activity evaluated using the Bovine Serum Albumin (BSA) denaturation method demonstrated that the protein hydrolysate of S. platensis consistently exhibited higher inhibitory activity than the crude extract at all tested concentrations (Table 7). At 25 μg/mL, the protein hydrolysate showed an inhibition of 38.02 ± 1.45%, which was more than twice that of the crude extract (17.71 ± 0.63%). This trend continued at 50 μg/mL, where the hydrolysate achieved 42.64 ± 0.81% inhibition, exceeding the crude extract value of 31.10 ± 2.90%. At the highest concentration tested (100 μg/mL), the hydrolysate reached 46.47 ± 0.25% inhibition, which was higher than both the crude extract (40.87 ± 0.21%) and the positive control sodium diclofenac (40.27 ± 0.21%). Statistical analysis using ANOVA followed by Tukey’s test confirmed that the differences in inhibition percentages among treatments at each concentration were significant (p < 0.05).
The superior anti-inflammatory activity of the S. platensis protein hydrolysate can be attributed to the formation of bioactive peptides during bromelain-assisted enzymatic hydrolysis. These peptides possess smaller molecular sizes and simpler structures, enabling more effective interaction with BSA and stronger inhibition of protein denaturation, a key indicator of inflammatory processes [45]. The markedly higher inhibition observed at 25 μg/mL indicates that enzymatic hydrolysis enhances the exposure of active sites and increases peptide accessibility, while simultaneously disrupting the native protein structure. This structural modification also influences the binding of pigment molecules to peptides, thereby significantly enhancing biological activity and improving protection against inflammation-induced protein damage [46]. As the concentration increased to 50 and 100 μg/mL, the inhibition effect of the hydrolysate continued to rise, confirming a concentration-dependent response and demonstrating superior efficacy compared to the crude extract. The hydrolysate’s ability to outperform sodium diclofenac at 100 μg/mL suggests that it may provide a competitive natural alternative to synthetic anti-inflammatory agents. This enhanced activity is likely associated with increased levels of specific amino acids, such as L-glutamate, L-leucine, and L-valine, following hydrolysis, which are known to contribute to anti-inflammatory and antioxidant mechanisms through membrane stabilization and free radical neutralization [30].
Table 7 Anti-inflammatory activity affected by crude extract and protein hydrolysate of Spirulina platensis.
Concentration (ppm) |
Diclofenac sodium (%) (Control Positive) |
Crude extract protein Spirulina platensis (%) |
Hydrolysate protein Spirulina platensis (%) |
0 |
0.000 0.000a |
0.000 0.000a |
0.000 0.000a |
25 |
15.969 1.905b |
17.717 0.627b |
38.012 1.454b |
50 |
30.695 2.260c |
31.109 2.899c |
42.614 0.808c |
100 |
40.267 2.012d |
40.865 0.210d |
46.479 0.253d |
Note: The results in the same column followed by the different letter that significantly differ by Tukey’s test (p < 0.05).
Anti-inflammatory activity by RAW 264.7 cells
Measurement of nitric oxide (NO) production in LPS-stimulated RAW 264.7 macrophages (Table 8) revealed distinct, concentration-dependent inflammatory responses between the crude extract and the bromelain-derived protein hydrolysate of S. platensis (p < 0.05). The negative control (LPS −) showed negligible NO levels (0.00 ± 0.00 µM), confirming the absence of basal inflammation, while the positive control (LPS +) significantly elevated NO production to 3.98 ± 0.62 µM, indicating effective macrophage activation. At 50 µg/mL, the crude extract produced a higher NO level (4.38 ± 0.73 µM) than the LPS (+) control, indicating no inhibitory effect and suggesting a possible pro-inflammatory or stimulatory response at this concentration. Noticeable NO inhibition by the crude extract was only observed at higher concentrations, particularly at 200 µg/mL (0.49 ± 0.36 µM), which is consistent with its relatively high IC₅₀ value (136.1 µg/mL) and overall weak anti-inflammatory activity. In contrast, the protein hydrolysate significantly suppressed NO production even at 50 µg/mL (1.14 ± 0.44 µM) compared with the LPS (+) group and reduced NO levels to near basal values at 200 µg/mL (0.43 ± 0.12 µM), reflecting substantially stronger anti-inflammatory efficacy supported by a low IC₅₀ value (10.0 µg/mL). The distinct statistical grouping of hydrolysate-treated samples relative to the LPS (+) control confirms significant NO inhibition at low concentrations. These findings demonstrate that enzymatic hydrolysis markedly enhances the anti-inflammatory potential of S. platensis protein, likely through the generation of low-molecular-weight bioactive peptides capable of more effectively modulating iNOS activity and suppressing LPS-induced inflammatory signaling pathways, including NF-κB and MAPK [47].
The protein hydrolysate of S. platensis was more effective than the crude protein extract in inhibiting nitric oxide (NO) production because enzymatic hydrolysis with bromelain generated smaller, low–molecular weight peptides with higher biological activity. As reflected in Table 8, the hydrolysate markedly reduced NO levels even at the lowest tested concentration (50 µg/mL), producing NO values close to the negative control and yielding a very low IC₅₀ value (10.0 µg/mL), whereas the crude extract required much higher concentrations to achieve a comparable effect (IC₅₀ 136.1 µg/mL). These small peptides are more soluble and can more readily penetrate macrophage cell membranes, allowing direct interaction with intracellular inflammatory targets such as inducible nitric oxide synthase (iNOS) and upstream signaling pathways [48-50]. In addition, bromelain hydrolysis likely exposed or released bioactive amino acid sequences rich in hydrophobic and aromatic residues, which are known to enhance binding to inflammatory mediators and suppress NO synthesis more efficiently. Consequently, the hydrolysate exerts stronger inhibition of LPS-induced NO production than the crude extract, demonstrating that enzymatic modification significantly improves the anti-inflammatory potency of microalgae S. platensis protein.
Table 8 Nitric oxide (NO) production affected by crude extract and protein hydrolysate of Spirulina platensis.
Crude extract of Spirulina platensis protein |
Hydrolysate protein of Spirulina platensis |
||||
Concentration (µg/mL) |
IC50 (µg/mL) |
Nitric oxide (NO) (µM) |
Concentration (µg/mL) |
IC50 (µg/mL) |
Nitric oxide (NO) (µM) |
LPS (-) |
- |
0.00 ± 0.00a |
LPS (-) |
- |
0.00 ± 0.00a |
LPS (+) |
- |
3.98 ± 0.62b |
LPS (+) |
- |
3.98 ± 0.62c |
50 |
136.10 |
4.38 ± 0.73b |
50 |
10.00 |
1.14 ± 0.44b |
100 |
3.72 ± 0.14b |
100 |
0.89 ± 0.41b |
||
200 |
0.49 ± 0.36a |
200 |
0.43 ± 0.12ab |
||
Note: The results in the same column followed by the same letter do not significantly differ by Tukey’s test (p < 0.05).
Cell viability analysis using the WST assay demonstrated that both the crude extract and the protein hydrolysate of S. platensis were non-cytotoxic to RAW 264.7 macrophages under LPS stimulation, as all treatments maintained viability well above the 70% safety threshold relative to the negative control (LPS−, 100%) (Table 9). The positive control LPS (+) significantly increased cell viability to 194.90 ± 2.70%, which is consistent with macrophage activation and proliferation under inflammatory stimulation. Treatment with the crude extract resulted in a concentration-dependent decline in viability, decreasing from 158.21 ± 1.85% at 50 µg/mL to 125.78 ± 1.03% at 200 µg/mL, suggesting that higher concentrations of the crude extract may partially attenuate LPS-induced proliferative responses, possibly due to the presence of non-protein components or less bioavailable macromolecules. In contrast, the protein hydrolysate consistently exhibited significantly higher viability across all concentrations, with values of 195.67 ± 3.71%, 175.34 ± 3.34%, and 168.87 ± 0.74% at 50, 100, and 200 µg/mL, respectively, indicating superior cytocompatibility and a protective effect against inflammatory stress. The statistically significant differences among treatments (p < 0.05) further confirm that enzymatic hydrolysis improved the biological performance of S. proteins. The enhanced viability observed with the hydrolysate is likely attributable to the presence of low–molecular weight bioactive peptides that can support cellular metabolism, reduce oxidative stress, and modulate inflammatory signaling without inducing cytotoxicity [51]. Similar proliferative and non-toxic effects of protein hydrolysates have been reported by Lee et al. [52] for enzymatically hydrolyzed fish proteins. When considered together with the marked reduction in nitric oxide production and the low IC₅₀ value (10 µg/mL), the cell viability data reinforce that the S. platensis protein hydrolysate, particularly at 200 µg/mL, represents an optimal anti-inflammatory treatment that effectively suppresses inflammatory mediators while maintaining high macrophage viability, underscoring its safety and potential for functional or therapeutic applications.
Table 9 Cell viability affected by crude extract and protein hydrolysate of Spirulina platensis.
Crude extract of Spirulina platensis protein |
Hydrolysate protein of Spirulina platensis |
||
Concentration (µg/mL) |
Cell viability (%) |
Concentration (µg/mL) |
Cell viability (%) |
LPS (-) |
100.00 ± 0.01a |
LPS (-) |
100.00 ± 0.01a |
LPS (+) |
194.90 ± 2.702e |
LPS (+) |
194.90 ± 2.70c |
50 |
158.21 ± 1.853d |
50 |
195.67 ± 3.71c |
100 |
139.29 ± 0.75c |
100 |
175.34 ± 3.34b |
200 |
125.78 ± 1.03b |
200 |
168.87 ± 0.74b |
Note: The results in the same column followed by the same letter do not significantly differ by Tukey’s test (p < 0.05).
Conclusions
This study demonstrates that enzymatic hydrolysis of Spirulina platensis protein using bromelain effectively enhances antibacterial and anti-inflammatory, whereas the antioxidant activity, as evaluated by the FRAP assay, was detectable but not significantly improved compared to the crude protein under the applied conditions. The observed bioactivities were attributed to the generation of low-molecular-weight peptides, which were known to exhibit higher biological effectiveness than intact proteins, highlighting the significance of this approach for developing natural bioactive agents. The hydrolysate showed promising safety and efficacy profiles, supporting its potential application in functional foods, nutraceuticals, or pharmaceutical formulations. However, this study is limited to in vitro evaluations, and the specific peptide sequences responsible for the observed activities were not identified. Therefore, future research should focus on peptide characterization, mechanism-of-action studies, and in vivo validation to further substantiate the bioactive potential and practical applicability of Spirulina platensis protein hydrolysates.
Acknowledgements
The authors would like to express gratitude to the Universitas Brawijaya, Indonesia Student Academic Research Scheme (UB STARS) for the opportunity to participate in the final project research activity through a research collaboration with researchers from the National Research and Innovation Agency (BRIN) Cibinong (Contract No. 00623/UN10.A0101/B/TU/ 2025), as well as to the Applied Zoology Research Center of BRIN Cibinong, Faculty of Fisheries and Marine Science, and Department of Biology-Universitas Brawijaya, Indonesia for the support, facility, and opportunity provided during the study.
Declaration of Generative AI in Scientific Writing
The authors acknowledge the use of generative AI tools (e.g., Grammarly) in the preparation of this manuscript, specifically for language editing and grammar correction. No content generation or data interpretation was performed by AI. The authors take full responsibility for the content and conclusions of this work.
CRediT Author Statement
Zidan Armanda: Conceptualization/research design, Methodology, Data analysis, Investigation, Data Curation, Writing -Original Draft, Review & Editing/conclusion. Yunita Eka Puspitasari: Conceptualization/research design, Resources, Methodology, Data analysis, Validation, Writing-review & Editing/conclusion, Supervision/criticism. Dinia Rizqi Dwijayanti: Conceptualization/research design, Resources, Investigation, Data analysis, Validation, Writing –review & editing. Pamungkas Rizki Ferdian: Investigation, Data analysis, Validation, Writing –review & editing. Zahrotul Firdaus: Resources, Investigation, Data analysis, Validation, Writing –review & editing. Heder Djamaludin: Conceptualization/research design, Resources, Methodology, Data analysis, Validation, Writing -Original Draft, Review & Editing/conclusion, Supervision/criticism, Project administration, Funding acquisition.
References
[1] R Urban-Chmiel, A Marek, D Stępień-Pyśniak, K Wieczorek, M Dec, A Nowaczek and J Osek. Antibiotic resistance in bacteria—a review. Antibiotics 2022; 11(8), 1079.
[2] G Liu and M Qin. Analysis of the distribution and antibiotic resistance of pathogens causing infections in hospitals from 2017 to 2019. Evidence-Based Complementary and Alternative Medicine 2022; 2022, 3512582.
[3] M Ameena, M Arumugham, K Ramalingam, S Rajeshkumar and R Shanmugam. Evaluation of the anti-inflammatory, antimicrobial, antioxidant, and cytotoxic effects of chitosan thiocolchicoside-lauric acid nanogel. Cureus 2023; 15(9), e46003.
[4] DR Oktarina, Y Susilowati and E Halimah. The potential of Phyllanthus genus plants as immunomodulatory and anti-inflammatory. Indones. Journal of Pharmaceutical Biology 2021; 1(2), 47-77.
[5] H Djamaludin, I Inayatussakinah, AA Kusdiyarlistio, Z Armanda, DR Dwijayanti, IK Maharsih, KD Palupi, PR Ferdian, RR Elfirta and H Notonegoro. Chemical profile and antioxidant activity of various cysteine-proteases’ impact on Spirulina protein hydrolysate. Indonesian Journal of Chemistry 2025; 25(3), 811.
[6] Y Sun, Y Cui, R Wang, J Ma, H Sun, L Cheng and R Yang. The hydrolysis of pigment-protein phycoerythrin by bromelain enhances the color stability. Foods 2023; 12(13), 2574.
[7] CBN Kumari, N Ambalavanan, SR Kumar, J Mahendra and U Sudhakar. Anti-oxidant, anti-inflammatory and antimicrobial activity of aqueous extract from acerola and amla. Bioinformation 2024; 20(7), 765-770.
[8] Z Wang and X Zhang. Inhibitory effects of small molecular peptides from Spirulina (Arthrospira) platensis on cancer cell growth. Food & Function 2016; 7(2), 781-788.
[9] AOAC International. Official methods of analysis, 22nd edition (2023). AOAC International, Rockville, Maryland, 2023.
[10] A Domenighini and M Giordano. Fourier transform infrared spectroscopy of microalgae as a novel tool for biodiversity studies, species identification, and assessment of water quality. Journal of Phycology 2009; 45(2), 522-531.
[11] A Ali, S Iqbal, A Khan and I Rabbani. Nutritional, bio-functional, and antioxidant properties of enzymatic hydrolysates derived from Spirulina platensis proteins. Foods 2025; 14(24), 4242.
[12] Z Firdaus, SD Hardiningtyas and I Setyaningsih. Green extraction and purification of phycocyanin from Spirulina platensis: Evaluation of its bioactivity and photoprotection. Biochemical Engineering Journal 2025; 223, 109881.
[13] Y Sun, R Chang, Q Li and B Li. Isolation and characterization of an antibacterial peptide from protein hydrolysates of Spirulina platensis. European Food Research and Technology 2016; 242(5), 685-692.
[14] I Wiegand, K Hilpert and REW Hancock. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nature Protocols 2008; 3(2), 163-175.
[15] YA Bailey-Shaw, LA Williams, CE Green, S Rodney and AM Smith. In-vitro evaluation of anti-inflammatory potential of Jamaican plant extracts using BSA denaturation assay. International Journal of Pharmaceutical Sciences and Research 2017; 47(1), 145-153.
[16] DR Dwijayanti, MH Widyananda, FE Hermanto, A Soewondo, M Afiyanti and N Widodo. Anti-inflammatory activity of Euphorbia hirta extract via transcriptomic and NO inhibition in RAW 264.7 cells. Food and Agricultural Immunology 2024; 35(1), 2351360.
[17] NS Parimi, M Singh, JR Kastner, KC Das, LS Forsberg and P Azadi. Optimization of protein extraction from Spirulina platensis. Frontiers in Energy Research 2015; 3, 30.
[18] A Naseri, GS Marinho, SL Holdt, JM Bartela and C Jacobsen. Enzyme-assisted extraction of protein from Palmaria palmata. Algal Research 2020; 47, 101849.
[19] LND Duy, PT Duy, LTT Hoa, NTX Quynh and ND Ninh. Protein extraction from Spirulina platensis with cellulase assistance. Journal of Technology Education Science 2022; 17(1), 25-32.
[20] Z Akbarbaglu, A Ayaseh, B Ghanbarzadeh and K Sarabandi. Techno-functional and biological properties of Spirulina platensis peptides. Algal Research 2022; 66, 102755.
[21] S Sun, X Xu, X Sun, X Zhang, X Chen and N Xu. ACE inhibitory peptides from Ulva intestinalis. Marine Drugs 2019; 17(3), 179.
[22] J Pekkoh, K Ruangrit, C Pumas, K Duangjan, S Chaipoot, R Phongphisutthinant, I Jeerapan, K Sawangrat, W Pathom-Aree and S Srinuanpan. Transforming microalgal Chlorella biomass into cosmetically and nutraceutically protein hydrolysates using high-efficiency enzymatic hydrolysis approach. Biomass Conversion and Biorefinery 2023; 13(7), 6299-6315.
[23] CR Lisboa, AM Pereira, SP Ferreira and JAV Costa. Utilisation of Spirulina and Chlorella for enzymatic hydrolysates. Journal of Engineering Research and Applied Science 2014; 4(1), 29-38.
[24] A Noman, Y Xu, WQ Al-Bukhaiti, SM Abed, AH Ali, AH Ramadhan and W Xia. Influence of enzymatic hydrolysis conditions on the degree of hydrolysis and functional properties of protein hydrolysate obtained from Chinese sturgeon (Acipenser sinensis) by using papain enzyme. Process Biochemistry 2018; 67, 19-28.
[25] I Nugrahani, R Oktaviary, S Ibrahim, T Gusdinar and C Apsari. FTIR for peptide content estimation. Indonesian Journal of Pharmacy 2020; 31(2), 78-83.
[26] Đ Tintor, K Ninković, J Milošević and NĐ Polović. Gaining insight into protein structure via ATR-FTIR spectroscopy. Vibrational Spectroscopy 2024; 134(13), 103726.
[27] Y Ji, X Yang, Z Ji, L Zhu, N Ma, D Chen, X Jia, J Tang and Y Cao. DFT-calculated IR spectrum amide I, II, and III band contributions of N-methylacetamide fine components. ACS Omega 2020; 5(15), 8572-8578.
[28] Z Akbarbaglu, A Ayaseh, B Ghanbarzadeh and K Sarabandi. Techno-functional, biological and structural properties of Spirulina platensis peptides from different proteases. Algal Research 2022; 66, 102755.
[29] B Ling, T Cheng and S Wang. Recent developments in applications of radio frequency heating for improving safety and quality of food grains and their products: A review. Critical Reviews in Food Science and Nutrition 2020; 60(15), 2622-2642.
[30] J Rivera-Jiménez, C Berraquero-García, R Pérez-Gálvez, PJ García-Moreno, FK Espejo-Carpio, A Guadix and EM Guadix. Peptides and protein hydrolysates exhibiting anti-inflammatory activity: Sources, structural features and modulation mechanisms. Food & Function 2022; 13(24), 12510-12540.
[31] D Iram, MS Sansi, S Meena, AK Puniya and S Vij. In vitro antimicrobial and synergistic effect of fermented Indian zebu (Sahiwal) cow colostrum whey derived peptides with Lactobacillus rhamnosus against pathogenic bacteria. Journal of Food Science and Technology 2023; 60(10), 2568-2580.
[32] ME Brassesco, A Paupério, CD Pereira, JP Ferreira and M Pintado. Peptide-rich yeast fractions from brewer’s spent yeast: A scalable fractionation approach and their functional application in bakery products. Foods 2025; 14(7), 1144.
[33] D Agyei, CM Ongkudon, CY Wei, AS Chan and MK Danquah. Bioprocess challenges to the isolation and purification of bioactive peptides. Food and Bioproducts Processing 2016; 98, 244-256.
[34] L Zheng, Y Zhao, H Dong, G Su and M Zhao. Structure–activity relationship of antioxidant dipeptides: Dominant role of Tyr, Trp, Cys and Met residues. Journal of Functional Foods 2016; 21, 485-496.
[35] Y Mishra, HI Amin, V Mishra, M Vyas, PK Prabhakar, M Gupta, R Kanday, K Sudhakar, S Saini, A Hromić-Jahjefendić and AA Aljabali. Application of nanotechnology to herbal antioxidants as improved phytomedicine: An expanding horizon. Biomedicine & Pharmacotherapy 2022; 153, 113413.
[36] MB Takyar, SH Khajavi and R Safari. Evaluation of antioxidant properties of Chlorella vulgaris and Spirulina platensis and their application in order to extend the shelf life of rainbow trout (Oncorhynchus mykiss) fillets during refrigerated storage. LWT 2019; 100, 244-249.
[37] NY Htoo, O Kraseasintra, W Buncharoen, T Kaewkod, J Pekkoh, Y Tragoolpua, KS Khoo, S Chaipoot, S Srinuanpan and C Pumas. In vitro immunomodulation activity of protein hydrolysate from Spirulina (Arthrospira platensis): The ingredient of future foods. Frontiers in Marine Science 2024; 11, 1303025.
[38] M Bidram and MR Ganjalikhany. Bioactive peptides from food science to pharmaceutical industries: Their mechanism of action, potential role in cancer treatment and available resources. Heliyon 2024; 10(23), e40563.
[39] NK AlFadhly, N Alhelfi, AB Altemimi, DK Verma, F Cacciola and Narayanankutty. Trends and technological advancements in the possible food applications of Spirulina and their health benefits: A Review. Molecules 2022; 27, 5584.
[40] M Vogelsang-O’Dwyer, AW Sahin, EK Arendt and Zannini. Enzymatic hydrolysis of pulse proteins as a tool to improve techno-functional properties. Foods 2022; 11(9), 1307.
[41] MS Qoms, SK Wong, NM Fauzi, K Husain, S Makpol and JK Tan. Microalgae-derived peptides targeting lifestyle-related diseases: Discovery, mechanisms, structure–activity relationships, and structural modifications. Antioxidants 2025; 14(10), 1170.
[42] AH Alkatheri, PSX Yap, A Abushelaibi, KS Lai, WH Cheng and SHE Lim. Host–bacterial interactions and antimicrobial peptides. Membranes 2022; 12(7), 715.
[43] MN Calabrese, CA Tianco, AM Garate, MC Tan, J Ting, RA Galian, AP Bonto and AC Chang. Evaluation of Spirulina spp. crude extract revealed antimicrobial, antioxidant and UV photoprotective properties. Trends in Sciences 2024; 21(6), 7582.
[44] JC Stephani, L Gerhards, B Khairalla, IA Solov’Yov and I Brand. How do antimicrobial peptides interact with the outer membrane of gram-negative bacteria? Role of lipopolysaccharides in peptide binding, anchoring, and penetration. ACS Infectious Diseases Journal 2024; 10(2), 763-778.
[45] J Bu, N Luo, C Shen, C Xu, Q Zhu, C Chen, Y Xie, X Liu, Y Liu, C Luo and X Zhang. A fast and efficient virtual screening and identification strategy for helix peptide binders based on finDr webserver: A case study of bovine serum albumin (BSA). International Journal of Biological Macromolecules 2025; 306, 141118.
[46] R Zhang, J Chen, X Mao, P Qi and X Zhang. Anti-inflammatory and anti-aging evaluation of pigment–protein complex extracted from Chlorella pyrenoidosa. Marine Drugs 2019; 17(10), 586.
[47] NY Htoo, T Kaewkod, W Buncharoen, J Pekkoh, Y Tragoolpua, S Chaipoot, KS Khoo, B Cheirsilp, C Pumas and S Srinuanpan. Bioactive properties of protein hydrolysate from Arthrospira (Spirulina) platensis: A review. Applied Food Research 2025; 5(2), 101245.
[48] LM Moreno-Vargas and D Prada-Gracia. Exploring the chemical features and biomedical relevance of cell-penetrating peptides. International Journal of Molecular Sciences 2024; 26(1), 59.
[49] A Rahman, MA Matthews, CJ Nowell, DK Chalmers, PE Thompson, SE Nicholson, N Barlow and RS Norton. Enhanced nitric oxide production by macrophages treated with a cell-penetrating peptide conjugate. Bioorganic Chemistry 2022; 123, 105763.
[50] M Zahid and PD Robbins. Cell-type specific penetrating peptides: Therapeutic promises and challenges. Molecules 2015; 20(7), 13055-13070.
[51] Y Oh, WK Jung and JY Je. Protective effect of multifunctional peptides PIISVYWK and FSVVPSPK on oxidative stress-mediated HUVEC injury through antioxidant and anti-apoptotic action. Process Biochemistry 2023; 125, 121-129.
[52] SJ Lee, J Im, SD Marasinghe, E Jo, MS Bandara, Y Lee, J Lee, GH Park and C Oh. Antioxidant and anti-inflammatory activities of cutlassfish head peptone in RAW 264.7 macrophages. Antioxidants 2025; 14(3), 286.